1.1 Вступительное занятие. Функциональные особенности возбудимых. Биопотенциалы
 Скачать 273.99 Kb. Скачать 273.99 Kb.
|

БиопотенциалыБиопотенциалы Луиджи Гальвани 1791 г. в эксперименте показал, что живые ткани содержат «животное электричество», его научный оппонент, физик Вольта - что это электричество от разнородных металлов, он создал первый источник постоянного тока, который носит название гальванический элемент. Виды биопотенциалов: 1. Биопотенциал покоя (мембранный) - МПП. 2. Биопотенциал действия (возбуждения) - ПД. Современная, экспериментально доказанная, мембранно-ионная теория возникновения биопотенциалов (Ходжкин, Хаксли, Катц). Основные положенияОсновные положения: I. Электрические процессы возникают на плазматической мембране клетки, которая состоит из бимолекулярного слоя липидов (остов мембраны) и белков, которые выполняют различные функции в мембране: рецепторную, ферментативную, образуют в ней каналы и насосы. Канал мембраны может быть неспецифическим, он постоянно открыт, не имеет воротного механизма, электрические воздействия не изменяют его состояния. Называют каналом «утечки». Специфические каналы (селективные) имеют воротный механизм, поэтому могут находиться или в открытом, или в закрытом состоянии в зависимости от электрических воздействий на мембрану и пропускают только определенный ион. Этот канал состоит из трех частей: водной поры – выстлана внутри гидрофильными группами; селективного фильтра – на наружной поверхности, который пропускает ионы в зависимости от их размера и формы; ворот – на внутренней поверхности мембраны, управляют проницаемостью канала. Каналы для натрия имеют два типа ворот: быстрые активационные и медленные инактивационные. В покое открыты медленные инактивационные и закрыты быстрые активационные. При возбуждении происходит открытие быстрых активационных и медленное закрытие медленных инактивационных, т.е. на короткий промежуток времени оба типа ворот открыты. Калиевые каналы имеют только медленные ворота. Насосы выполняют функцию транспорта через мембрану ионов против градиента концентрации, для их работы используется энергия АТФ. II. По обе стороны мембраны существует концентрационный градиент. Внутри клетки в 40 раз > К+; Вне клетки: в 20-30 раз > Na+, в 50 раз > Cl-. III. Мембрана пропускает молекулы жирорастворимых веществ, а анионы органических кислот не проходят. Мембрана проницаема для воды, для ионов проницаемость мембраны различна: для калия в состоянии покоя проницаемость почти в 25 раз больше, чем для натрия. При возбуждении увеличивается проницаемость и для калия (постепенно), и для натрия (быстро, но на очень короткий промежуток времени). Мембранный потенциал покояМембранный потенциал покоя (МПП) - это разность потенциалов между наружной и внутренней поверхностью мембраны клетки в покое. Наружная поверхность мембраны клетки имеет положительный заряд, а внутренняя - отрицательный. Биопотенциал покоя регистрируется внутриклеточным методом - с помощью микроэлектродов, один из которых вводится внутрь клетки. В эксперименте биопотенциал покоя можно зарегистрировать между повреждённым и неповреждённым участком ткани. Повреждённый участок является моделью внутренней поверхности мембраны клетки. 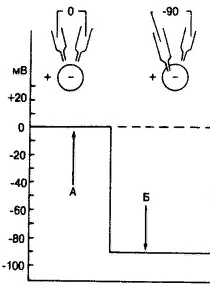 Рис. Регистрация внутриклеточным методом мембранного потенциала покоя Рис. Регистрация внутриклеточным методом мембранного потенциала покояВверху -- схематическое изображение клетки (кружок) и двух микроэлектродов. Фрагмент А -- оба микроэлектрода находятся во внеклеточном пространстве и разности потенциалов между ними нет; Б -- кончик одного микроэлектрода введен в клетку, что позволяет регистрировать разность потенциалов между внутренним пространством клетки и внеклеточной средой; в данном случае это потенциал покоя, равный --90 мВ; Проницаемость мембраны для ионов К повышена, поэтому калий играет основную роль в генерации МПП. Калий создаёт электрическое поле и заряжает наружную поверхность мембраны «+». В тот момент, когда «+» потенциал наружной стороны достигает определённой величины по отношению к «-» внутри, который создается анионами - наступает динамическое равновесие между входящими и выходящими из клетки ионами К+. Этому моменту соответствует потенциал равновесия для К - потенциал покоя. МППМПП характеризуется: 1. постоянством; 2. полярностью, снаружи «+», внутри «-»; 3. величина – в мВ, для скелетной мышцы - 60 – 90 мВ, для гладкой - -30 – 70 мВ, для нерва -50 – 80мВ, для секреторной клетки - -20мВ. МПП - один из основных показателей состояния физиологического покоя клетки. При увеличении внеклеточной концентрации калия уменьшается МПП, т.к. уменьшается диффузия калия из клетки в связи со снижением его концентрационного градиента. При действии веществ, блокирующих ресинтез АТФ, т.к. прекращается работа натрий-калиевого насоса, также снижается МПП. Ионы натрия и хлора входят в клетку, но ввиду низкой проницаемости значительного влияния на МПП не оказывают. Потенциал действия – это кратковременные высокоамплитудные изменения МПП, которые возникают при возбуждении. ПД регистрируется в раздражаемых тканях, в которых возникает волна возбуждения. Измеряется ПД с помощью внутриклеточного и внеклеточного отведений. При возбуждении – резко увеличивается проницаемость для ионов Na, которые поступают внутрь клетки лавинообразно и заряжают внутреннюю сторону «+» - происходит деполяризация мембраны, а затем количество ионов натрия внутри превышает калиевый заряд на поверхности и это приводит к перезарядке мембраны (реверсии). Постепенно увеличивающаяся проницаемость для калия и его поток из клетки инактивирует натриевую проницаемость и приводит к восстановлению заряда на мембране. Возникает фаза реполяризации. Существенным фактором является натрий-калиевый насос, который выводит из клетки 3 иона натрия в обмен на 2 иона калия, вводимые в клетку. Его работа зависит от метаболизма клетки, в частности, от ее энергоснабжения. При этом расходуется 1 молекула АТФ. ПД состоит из пикового потенциала, образуемого фазой деполяризации, реверсии и реполяризации, и следовых потенциалов. Следовые потенциалы: Отрицательный (следовая деполяризация); Положительный (следовая гиперполяризация). Причиной следовых потенциалов являются дальнейшие изменения соотношения между входом натрия в клетку и выходом калия из нее. При следовой деполяризации отмечается остаточный ток натрия в клетку при одновременном снижении калиевого тока. При следовой гиперполяризации - остаточное усиление тока калия из клетки при одновременной активации натрий-калиевого насоса.  ПД характеризуетсяПД характеризуется: 1. изменяющимся характером; 2. кратковременностью – несколько мсек; 3. зарядом мембраны, снаружи – «-», внутри – «+». При действии веществ, блокирующих натриевые каналы, ПД не генерируется, т.к. в норме деполяризация мембраны обусловлена повышение ее натриевой проницаемости. При увеличении силы раздражителя выше порога амплитуда ПД не изменяется, т.к. не изменяется число активированных натриевых каналов, которые максимально раскрываются при пороговом раздражении. КЛЮЧЕВЫЕ ПОНЯТИЯ И ОПРЕДЕЛЕНИЯ1. К возбудимым тканям относятся: нервная , мышечная , железистая. 2. Микроэлектродные методы , позволяющие зарегистрировать значение биопотенциалов : внеклеточный , внутриклеточный. 3. Значение МПП регистрируется только внутриклеточным методом. 4. Значение ПД внутриклеточным и внеклеточным методом . 5. Фазы ПД : локальный ответ , деполяризация, реполяризация . 6. Фаза деполяризации характеризуется повышением проницаемости клеточной мембраны для ионов натрия и входом его в клетку. 7. Фаза реполяризация характеризуется повышением проницаемости клеточной мембраны для ионов калия и выходом его из клетки 8. КУД- это уровень электрического потенциала мембраны возбудимой клетки, от которого локальный потенциал переходит в потенциал действия. 9. Порог раздражения – это мера возбудимости клетки, наименьшая сила раздражителя , способная вызвать распространяющийся потенциал действия . 10. МПП нервной клетки составляет -50-80 Мв 11. МПП мышечной клетки ( гладкой ) составляет -30 -70 Мв 12. МПП мышечной клетки ( скелетной ) составляет - 60 – 90 мВ 13. МПП секреторной клетки составляет -20Мв 14. Неспецифические каналы (каналы «утечки») – это постоянно открытые каналы, не имеющие воротного механизма, электрические воздействия на которые не изменяют их состояния. 15. Специфические каналы (селективные)- имеют воротный механизм, поэтому могут находиться или в открытом, или в закрытом состоянии в зависимости от электрических воздействий на мембрану и пропускают только определенный ион. 16. Мембранный потенциал покоя (МПП) - это разность потенциалов между наружной и внутренней поверхностью мембраны клетки в покое. 17. Мембрана клетки в состоянии покоя заряжена отрицательно изнутри и положительно снаружи. 18. Потенциал действия – это кратковременные высокоамплитудные изменения МПП, которые возникают при возбуждении. ПД регистрируется в раздражаемых тканях, в которых возникает волна возбуждения. 19. Натрий-калиевый насос, выводит из клетки 3 иона натрия в обмен на 2 иона калия, вводимые в клетку. 20. Следовые потенциалы: следовая деполяризация , следовая гиперполяризация. 21. Следовая деполяризация- это отрицательный следовой потенциал. 22. Следовая гиперполяризация - это положительный следовой потенциал 23. При следовой деполяризации отмечается остаточный ток натрия в клетку при одновременном снижении калиевого тока. 24. При следовой гиперполяризации - остаточное усиление тока калия из клетки при одновременной активации натрий-калиевого насоса. 25. Фазы возбудимости при возбуждении: супернормальная возбудимость ( первичная) , абсолютная рефрактерность, относительная рефрактерность, супернормальная возбудимость ( вторичная ), субнормальная возбудимость. 26. В период абсолютной рефрактерности клетка является абсолютно невозбудимой и не отвечает даже на сверхпороговые раздражители. 27. В период относительной рефрактерности возбудимость клеточной мембраны существенно снижена , но клетка отвечает на сверхпороговые раздражители. 28. Абсолютная рефрактерность соответствует фазе деполяризации 29. Относительная рефрактерность соответствет фазе реполяризации 30. Супернормальная возбудимость ( первичная) соответствует фазе локальногог ответа 31. Супернормальная возбудимость ( вторичная ) соответствует фазе следовой деполяризации 32. Субрнормальная возбудимость соответствует фазе следовой гиперполяризации. |