биосинтез днк. Биосинтез ДНК презентация. Биосинтез днк (репликация) является
 Скачать 180.07 Kb. Скачать 180.07 Kb.
|

|
Биосинтез ДНК (репликация) является: Матричным (матрица – обе нити ДНК) комплиментарным фрагментарным (нити ДНК синтезируются в виде фрагментов, которые затем соединяются между собой) полуконсервативным (в каждой из образовавшихся молекул ДНК одна нить исходная – материнская, а одна – вновь синтезированная – дочерняя) Полуконсервативность биосинтеза ДНК 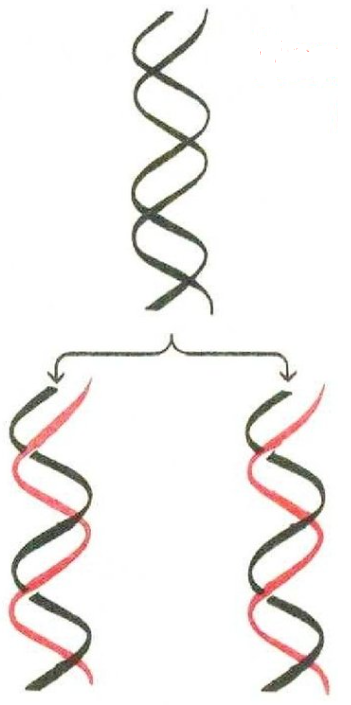 7. Этапы репликации 1. Инициация: Топоизомераза находит точку начала репликации, гидролизует одну фосфодиэфирную связь и даёт возможность компонентам репликативной системы разомкнуть нити ДНК и образовать репликативную «вилку», а затем вновь соединяет связь между мононуклеотидами Хеликаза разрывает водородные связи между нитями ДНК ДНК-связывающие белки (SSB-белки) стабилизируют репликативную вилку, не давая восстанавливаться водородным связям между комплиментарными нуклеотидами ДНК-полимераза α (праймаза) строит праймер(«затравку») из 8-10 рибонуклео-тидов и 4050 дезоксирибонуклеотидов, а ДНКполимераза δ достраивает нить из дезоксирибонуклеотидов на лидирующей нити, а ДНК-полимераза ε – на отстающей нити ДНК 11. Этапы репликации 2. Элонгация ДНК-полимераза δ продолжает удлинять нить из дезоксирибонуклеотидов на лидирующей нити, а ДНК-полимеразы α и ε – строить фрагменты из праймеров и дезоксирибонуклеотидов (фрагменты Оказаки) на отстающей нити ДНК по мере движения репликативной вилки. Этапы репликации 3. Терминация ДНК-полимераза β (фермент репарации) удаляет праймеры и достраивает фрагменты ДНК ДНК-лигаза соединяет фрагменты между собой. 16. Для биосинтеза РНК (транскрипции) необходимы: •МАТРИЦА – участок одной из нитей ДНК – (транскриптон) •СТРОИТЕЛЬНЫЙ МАТЕРИАЛ: – АТФ, ГТФ, ЦТФ, УТФ •ФЕРМЕНТАТИВНЫЕ БЕЛКИ ДНК-зависимые РНК-полимеразы I — для синтеза р-РНК II — для синтеза м-РНК III — для синтеза т-РНК •РЕГУЛЯТОРНЫЕ БЕЛКИ: факторы инициации, элонгации, терминации 17. Биосинтез РНК 18. Биосинтез РНК РНК-полимераза Биосинтез РНК Транскрибируемая нить ДНК Нетранскрибируемая нить мононуклеотиды пре-РНК (первичный транскрипт) Сайт терминации 19. Процессинг РНК (1. сплайсинг) 20. Процессинг (2. модификация концов м-РНК) 5' 3' м-РНК полиаденилат 7-метилгуанозин 21. Состав зрелой м-РНК 22. Общая схема биосинтеза белка 23. Компоненты белоксинтезирующей системы мРНК 20 Аминокислот 20 Аминоацил-тРНК синтетаз (АРС-аз) Изоакцепторные тРНК Рибосомы в виде полисом Источники энергии (АТФ, ГТФ) и Мg2+ Белковые факторы регуляции: факторы инициации, элонгации, терминации Специальные ферменты посттрансляционного процессинга полипептидной цепи 24. Ядерные стадии биосинтеза белка Биосинтез мРНК (транскрипция) Созревание мРНК (посттранскрипционный процессинг) 25. Функции т-РНК Акцепторная: связывает аминокислоту и транспортирует её к месту синтеза белка Адаптерная: встраивает аминокислоту на соответствующее место в образующемся полипептиде (за счёт взаимодействия антикодона в т-РНК с кодоном в м-РНК) антикодон 27. Таблица генетического кода 28. Характеристика генетического кода Триплетность (1 аминокислота кодируется 3нуклеотидами) Специфичность (каждому кодону соответствует только 1 аминокислота) Вырожденность (1 аминокислота может кодироваться более чем 1 триплетом) Линейная запись (прочтение кода «без знаков препинания») Универсальность (одинаков для всех живых существ) Наряду со значимыми есть и «бессмысленные» кодоны (терминирующие – УАА, УАГ, УГА) Колинеарность (соответствие линейной последовательности кодонов гена и последовательности АК в кодируемом белке) 29. Колинеарность генетического кода Глу-тРНК 30. Цитоплазматические стадии биосинтеза белка Активация аминокислот, или образование аминоацил-тРНК Инициация Элонгация Терминация Посттрансляционная модификация 31. Синтез аминоацил-тРНК 32. Рибосома эукариотов 33. Функции активной рибосомы Связывание и удерживание белоксинтезирующей системы Транслокация – перемещение м-РНК через рибосому каждый раз на один триплет Замыкание пептидной связи между аминокислотами (каталитическая функция) 34. Функционирующая рибосома 35. Образование инициирующего комплекса 36. Инициация: К малой субъединице рибосомы прикрепляется м-РНК К инициирующему кодону прикрепляется Мет-тРНК К образовавшемуся комплексу присоединяется большая субъединица таким образом, что Метт-РНК оказывается в пептидильном центре, а в аминоацильном – кодон, соответствующий второй аминокислоте 37. Инициация трансляции 38. Элонгация: В пептидильном центре находится Мет-т-РНК, а в аминоацильном – аа-тРНК, соответствующая второму кодону. Мет отрывается от т-РНК и перебрасывается в аминоацильный центр С помощью пептидилтрансферазы замыкается пептидная связь между карбоксильной группой Мет и аминогруппой второй аминокислоты (требуется энергия ГТФ и регуляторный фактор Т) Происходит передвижение рибосомы по м-РНК на один триплет (транслокация) В освободившийся аминоацильный центр входит аа-т-РНК, соответствующая третьему кодону и т.д. 39. ГТФ ГДФ+НР 40. Строение полирибосомы 41. Посттрансляционный процессинг Модификация N-конца полипептидной цепи Фолдинг (формирование пространственной структуры) Химическая модификация (гидроксилирование, гликозилирование и др.) Присоединение простетических групп (у гетеропротеинов) Объединение протомеров при образовании олигомерных белков Присоединение сигнальных пептидов для выхода белка из клетки 42. Регуляция биосинтеза 1: РНК полимераза, 2: Репрессор, 3: Промотор, 4: Оператор, 5: Лактоза, 6, 7, 8: гены, кодирующие лактазу. 43. Действие регуляторных белков 44. Регуляция биосинтеза Энхансеры – участки ДНК, присоединение к которым регуляторных белков усиливает транскрипцию Сайленсеры – участки ДНК, присоединение к которым регуляторных белков уменьшает транскрипцию 45. Амплификация 46. Типы генов в геноме Структурные гены (кодируют белки) Регуляторные гены: Гены-регуляторы (регулируют работу структурных генов) Процессинг-гены (регулируют посттранскрипционные и посттрансляционный процессинг) Темпоральные гены (включают в работу структурные гены в ходе клеточной дифференцировки) 47. Клеточная дифференцировка 48. Классификация мутаций 49. Типы мутаций 50. Типы генных мутаций 51. Проявления мутаций Вид мутации Изменения в структуре ДНК Изменения в структуре белка Замена: Замена в кодоне Белок не изменён нейтральная без изменения смысла одного нуклеотида другим кодона «миссенс-мутация» с изменением смысла кодона Замена одной аминокислоты на другую «нонсенс-мутация» с образованием терминирующего кодона На мутантном триплете синтез пептидной цепи прекращается 52. 53. Механизмы увеличения числа и разнообразия генов в геноме 54. Механизмы увеличения числа и разнообразия генов в геноме (кроссинговер) 55. Механизмы увеличения числа и разнообразия генов в геноме (амплификация) 56. Полиморфизм белков – изменение первичной структуры белка в пределах одного вида без изменения функций. В его основе лежит генетическая гетерогенность молекул ДНК. 57. Схема возникновения наследственных болезней 58. Биохимические проявления наследственных болезней 59. Клинические проявления наследственных болезней Третичная структура ДНК: нуклеопротеидные комплексы (хромосомы) Гистоновые белки: белки с высоким содержанием лиз и арг 5 типов: Н1, Н2А, Н2В, Н3, Н4 Негистоновые белки: белки и ферменты, участвующие в матричных биосинтезах Роль белков: обеспечивают суперспирализацию и компактизацию ДНК Нуклеосома ДНК (≈146 н.п.) + 8 молекул гистонов (Н2А, Н2В, Н3, Н4)2 Структура удерживается ионными связями между лиз, арг и остатками фосфорной кислоты Линкерные участки Участок ДНК (≈30 н.п.) между нуклеосомами, с которым связаны молекулы гистона Н1 Гетерохроматин – «компактный» хроматин, транскрипционно неактивный Эухроматин – деспирализованный хроматин с низким содержанием гистонов и высоким содержанием негистоновых белков (период транскрипции) 14Пространственная структура РНК • Одноцепочечная • Шпильки – спирализованные участки (водородные связи) • Не соблюдается правило Чаргаффа • Виды РНК: мРНК матрица в синтезе белка 2-4% от общего количества РНК, разнообразная первичная структура 5′ - «кэп»-конец: 7-метил ГТФ (защита от нуклеаз, участие в инициации трансляции) 3′ - поли(А)-«хвост»: 150-200 остатков АМФ (выход из ядра, защита от 16 нуклеаз) 17. тРНК молекулы-адапторы: переводят информацию мРНК в последовательность аминокислот в белке 15% содержат минорные нуклеотиды (например, метилированные АО) 17 Структура тРНК: 1 – шпильки 2 - петли 18. рРНК структурный компонент рибосом 80% от общего количества РНК в клетке 4 типа у эукариот: 5S, 5,8S, 18S, 28S S – единица Сведберга, скорость осаждения при центрифигировании 18 19. РЕПЛИКАЦИЯ: синтез ДНК Протекает в ядре в S-фазу клеточного цикла перед митозом Стимулы: гормоны, ростовые факторы, белки-циклины Матрица: обе нити ДНК, образуются 2 репликативные вилки Направление синтеза новых цепей: 5′ - 3′ по принципу комплиментарности и антипараллельности Участки синтеза – ориджины репликации Участок ДНК между соседними ориджинами - репликон Этапы репликации: инициация, элонгация, терминация Субстраты и источники энергии: дАТФ, дГТФ, дТТФ, дЦТФ Кофактор: Mg2+ Полуконсервативный процесс синтеза: каждая дочерняя молекула ДНК содержит одну родительскую нить и одну синтезированную Образуется идентичная молекула ДНК (клетка 4n) 19 20. 1 этап репликации: инициация Формирование репликативной вилки: 1. ДНК-топоизомераза расщепляет 3′,5′фосфодиэфирную связь в одной из цепей ДНК и присоединяется к 5′-концу в точке разрыва 2. ДНК-хеликаза, используя энергию АТФ, разрывает водородные связи и обеспечивает локальное разделение двойной спирали ДНК • ДНК-топоизомераза восстанавливает 3′,5′фосфодиэфирную связь и отделяется • SSB (single strand binding)–белки связываются с одноцепочечными участками, препятствуя комплементарному скручиванию цепей 21. Схема инициации репликации 22. 2 этап репликации: элонгация Синтез новых цепей ДНК Лидирующая цепь: 3′ - 5′ (синтез непрерывный по ходу движения репликативной вилки) Отстающая цепь: 5′ - 3′ (рост этой цепи начинается после того, как на лидирующей цепи синтезируется участок из ≈200 нуклеотидов, синтез идет против движения репликативной вилки в виде фрагментов Оказаки) Синтез цепей начинается с образования «затравки» (РНК-праймера из ≈10 нуклеотидов) Ферменты: ДНК-полимераза α синтезирует РНК-праймер и небольшой участок ДНК ДНК-полимераза δ удлиняет лидирующую цепь ДНК-полимераза δ или ε удлиняют отстающую цепь 23. 3 этап репликации: терминация Исключение праймеров Завершение формирования отстающей цепи ДНК Эндонуклеаза (РНКаза) удаляет РНКпраймер ДНК-полимераза β заполняет «брешь» ДНК-лигаза объединяет фрагменты, затрачивая энергию АТФ 24. Схема репликативной вилки 25. Репарация ошибок и повреждений ДНК Причина повреждений ДНК: действие факторов окружающей и внутренней среды Повреждение ДНК происходит с частотой от нескольких сотен до 1000 случаев в каждой клетке, каждый час Виды повреждений: дезаминирование АО (цитозин превращается в урацил), метилирование АО депуринизация, депиримидинизация, образование пиримидиновых димеров (действие УФО) разрыв цепей, ковалентные сшивки между цепями ошибки репликации Система репарации – ферменты (нуклеазы, полимеразы, лигазы) 26. Схема работы системы репарации ДНК 27. Роль системы репарации Репарация необходима для сохранения генома и возможна благодаря существованию 2-х цепей ДНК. Снижение активности ферментов репарации приводит к накоплению мутаций Полагают, что от 80 % до 90 % всех раковых заболеваний связаны с нарушением репарации ДНК ПРИМЕР: пигментная ксеродерма – наследственное заболевание, связанное с мутацией генов системы репарации ДНК; УФО таких больных приводит к накоплению мутаций в клетках кожи и развитию рака 28. ТРАНСКРИПЦИЯ: синтез РНК Протекает в ядре вне зависимости от фаз клеточного цикла Матрица: нить ДНК 3′ - 5′ Субстраты и источники энергии: АТФ, ГТФ, ЦТФ, УТФ Направление синтеза: 5′ - 3′ по принципу комплиментарности и антипараллельности Этапы: инициация, элонгация, терминация Участвуют факторы инициации, элонгации и терминации Образуются комплиментарные матрице продукты: мРНК, тРНК, рРНК Ферменты: РНК-полимераза I (синтез пре-рРНК) РНК-полимераза II (синтез пре-мРНК) 28РНК-полимераза III (синтез пре-тРНК) 29. 1 этап транскрипции: инициация Промотор – последовательность ДНК (ТАТА), с которой связывается РНК-полимераза Сайт терминации – участок завершения синтеза РНК Транскриптон – участок ДНК ограниченный промотором и сайтом терминации 1. «Активация» промотора с помощью ТАТА-фактора 2. Взаимодействие промотора с РНК-полимеразой и факторами инициации Факторы инициации обеспечивают расплетение двойной нити ДНК длиной в один виток (10 н.п.) 30. 2 этап транскрипции: элонгация и терминация Элонгация: рост нити пре-РНК Факторы элонгации (E, H, F) повышают активность РНК-полимеразы и облегчают расхождение цепей. Один ген может одновременно транскрибироваться несколькими молекулами РНК-полимеразы Терминация: прекращение транскрипции Факторы терминации облегчают отделение пре-РНК иьРНК-полимеразы от матрицы ДНК 31. Схема транскрипции 32. Посттранскрипционные модификации пре-РНК «Созревание» пре-мРНК «Кэпирование» на стадии элонгации Образование поли(А)- «хвоста» после транскрипции Сплайсинг – удаление интронов (некодирующих последовательностей) и соединение экзонов Участвуют малые ядерные рибонуклеопротеины (мяРНП), образующие комплексы – сплайсосомы Выход «зрелой» мРНК в цитоплазму Альтернативный сплайсинг – механизм образования различных видов «зрелой» мРНК из одной и той же молекулы пре-мРНК в разных тканях. В результате в разных тканях при считывании информации с одного и того же гена образуются различные мРНК, а соответственно и различные белки. 33. Схема «созревания» пре-мРНК 34. «Созревание» пре-тРНК 1. Удаление интронов 2. Модификация азотистых оснований (10-15%) Формирование акцепторного участка и антикодона 3. Выход зрелых тРНК в цитоплазму 35. «Созревание» пре-рРНК 36. ТРАНСЛЯЦИЯ: синтез белка Место синтеза: рибосомы, Матрица: мРНК Субстраты: аминокислоты (АК), Источники энергии: АТФ, ГТФ, Кофактор: Mg 2+ (стабилизирует структуру рибосом), Факторы инициации (IF), элонгации (EF), терминации (RF) Активация АК: связывание с тРНК (аминоацил-тРНК-синтетазы) Инициирующая аминоацил-тРНК (аа-тРНК): мет-тРНК Инициирующий кодон мРНК: AUG Этапы: инициации, элонгации, терминации Образуется колинеарный матрице продукт – белок Адапторы: тРНК (последовательность АК соответствует последовательности кодонов мРНК) Биологический код: запись информации о последовательности АК в белке с помощью последовательности нуклеотидов 37. Свойства биологического кода Триплетность Наличие терминирующих кодонов (UAA, UAG, UGA) Специфичность Вырожденность Универсальность Однонаправленность Колинеарность 38. Активация аминокислот 39. 1 этап трансляции: инициация К мРНК присоединяется малая субъединица рибосомы, фактор инициации IF, мет-тРНК и ГТФ. Когда комплекс свяжется с кодоном AUG, происходит присоединение большой субъединицы рибосомы, что сопровождается гидролизом ГТФ и отделением IF. Формируется полноценная рибосома с пептидильным (Р) и аминоацильным (А) центрами. 40. 2 этап трансляции: элонгация (рост пептидной цепи) Стадии элонгации: Связывание аа-тРНК в А-центре при участии фактора элонгации EF1 и с затратой энергии ГТФ Образование пептидной связи между АК Р-центра и АК А-центра при участии пептидилтрансферазы Перемещение рибосомы по мРНК (транслокация) в направлении от 5′- к 3′-концу с использованием энергии ГТФ и при участии фактора элонгации EF2 Многократное повторение стадий 41. 3 этап трансляции: терминация Высвобождение пептида из связи с тРНК и рибосомой: Стоп-кодоны UAA, UAG, UGA попадают в А-центр. Высвобождение полипептида при участии факторов терминации RF1, RF3 и энергии ГТФ 42. Посттрансляционные модификации белков – образование функционально активных белков Частичный протеолиз Фолдинг – формирование пространственной структуры (II, III) при участии белков-шаперонов Модификация аминокислот (гликозилирование, фосфорилирование, ацилирование, метилирование……). Образование дисульфидных связей (цистеин-цистеин), Присоединение простетической группы (сложные белки), Сборка протомеров в олигомерные белки (формирование IV структуры) 43. Регуляция матричных биосинтезов Экспрессия генов — процесс, в ходе которого наследственная информация от гена (последовательности нуклеотидов ДНК) преобразуется в функциональный продукт — РНК или белок (в процессе транскрипции и трансляции) Механизмы регуляции экспрессии генов различны: компактизация ДНК, модификация ДНК и гистонов, привлечение факторов транскрипции и др. Гены белков «домашнего хозяйства» экспрессируются с постоянной скоростью (конститутивные) и обеспечивают жизнеспособность клеток (например, гены ферментов энергетического обмена). Стойкая репрессия транскрипции определенных генов в различных клетках обеспечивает формирование специализированных клеток, тканей и органов. Адаптивная регуляция обеспечивает изменение скорости экспрессии генов в ответ на меняющиеся условия среды (индуцибельная экспрессия). Адаптивная регуляция осуществляется при участии: регуляторных белков, взаимодействующих с участками ДНК индукторов (стимулируют экспрессию) корепрессоров (подавляют экспрессию) Индукторы или корепрессоры стимулируют присоединение регуляторных белков к регуляторным участкам ДНК. В качестве индукторов и корепрессоров выступают гормоны, ростовые факторы, продукты метаболических путей. Регуляторные участки ДНК: Энхансер – «усилитель» транскрипции Сайленсер – «тушитель» транскрипции 45. Примеры адаптивной регуляции экспрессии генов КОРТИЗОЛ (как индуктор) стимулирует присоединение регуляторного белка к энхансеру и вызывает ФОСФОЕНОЛПИРУВАТКАРБОКСИКИНАЗЫ (ключевого экспрессию гена фермента синтеза глюкозы), что приводит к повышению уровня глюкозы в крови при голодании, стрессе и физической нагрузки ХОЛЕСТЕРИН (как корепрессор) стимулирует присоединение белка-регулятора к сайленсеру и вызывает подавление экспрессии гена ГМГ-КоА-РЕДУКТАЗЫ (ключевого фермента синтеза холестерина), что приводит к снижению синтеза холестерина (поэтому чем больше холестерина поступает с пищей, тем меньше его синтезируется) 46. Примеры ингибиторов матричных биосинтезов Токсин белой поганки аманитин ингибирует РНКполимеразу II (синтез мРНК) Энтеротоксин возбудителя дифтерии ингибирует трансляцию, модифицируя фактор элонгации EF2 и нарушая транслокацию рибосом, Интерфероны (гликопротеины лимфоцитов и макрофагов, обладающие противовирусной активностью): активируют РНК-азу, расщепляющую мРНК и рРНК стимулируют синтез протеинкиназы, которая фосфорилирует и тем самым инактивирует фактор инициации трансляции IF2 прекращается синтез белков в инфицированных клетках человека, клетка погибает, но останавливается размножение вирусов. 47. Задание для самостоятельной работы Используя интернет-ресурсы и учебник выполните задания и составьте конспект по вопросам: 1. Принцип метода полимеразной цепной реакции и его применение в медицине. 2. Роль нерепарированных изменений ДНК (мутаций) в развитии биохимической индивидуальности человека (полиморфизме генов и белков), наследственных заболеваний и канцерогенезе. 3. Заполните таблицу «Лекарственные препараты – ингибиторы матричных биосинтезов» (см. следующий слайд). 47 48. Противоопухолевые и антибактериальные препараты – ингибиторы матричных биосинтезов Препараты Механизм действия Ингибиторы репликации и транскрипции: Доксорубицин, дауномицин, Циклофосфан, мелфалан Фторхинолоны, Рифамицины Ингибиторы трансляции: Тетрациклин, Эритромицин, Левомицетин 49. Заключение Процессы репликации, транскрипции, трансляции (матричные биосинтезы) лежат в основе «производства» белков и ферментов, функционирование которых является основой жизни. Регуляция данных процессов лежит в основе адаптации. Нарушение данных процессов приводит к развитию заболеваний. Знания о нуклеиновых кислотах и механизмах матричных биосинтезов являются основой создания лекарственных препаратов, методов диагностики и терапии |