физиология. 1.1234.физиология. Цитоплазматические мембраны
 Скачать 457.9 Kb. Скачать 457.9 Kb.
|

"Биоэлектрические явления в возбудимых тканях".Информационный блок № 1:Раздражимость и возбудимость клетки и ткани. Меры возбудимости. Клетка обладает следующими основными физиологическими свойствами. 1. Раздражимость – это способность клетки отвечать на раздражение - (т.е. воздействие каких-то факторов) изменением своего обмена веществ. 2. Возбудимость – это способность клетки отвечать на раздражение изменением проницаемости клеточной мембраны, входящим натриевым током и, как следствие, генерацией потенциала действия. 3. Проводимость – это способность клетки проводить, распространять возбуждение от места его возникновения в клетке к другим ее частям. 4. Сократимость – это способность клетки под действием раздражителя изменять свою длину или (и) напряжение цитоскелета. Законы раздражения. 1. Закон силы раздражения: Чем больше сила раздражения, тем, до известных пределов, сильнее ответная реакция. Есть сила раздражения, которая способна вызывать максимальный эффект – оптимум частоты и силы раздражения. Если сила больше, чем оптимальная, то ответная реакция ниже – пессимум частоты или силы раздражения. 2. Закон длительности раздражения: Чем длительнее раздражение, необходимое для возникновения возбуждения, тем сильнее, до известных пределов, ответная реакция живых систем. 3. Закон градиента силы: Величина ответной реакции и её характер зависят ещё и от интенсивности/крутизны/ нарастания действия силы. 4. Закон "всё или ничего": Если раздражитель меньше пороговой силы, он никогда не вызовет ПД (потенциал действия) – «ничего». Но какой бы силы не был надпороговый раздражитель, он всегда будет вызывать max для данного состояния электрическую реакцию, т.е. max пик ПД – «всё». Лабильность(функциональная подвижность) – свойство клетки, ткани, отражающее их максимальные возможности. Мера лабильности - это максимальная частота, которую способна воспроизвести ткань или клетка. Частота раздражения близкая или совпадающая с величиной лабильности вызывает максимальный ответ, т.е. оптимум. Раздражение выше по силе или по частоте лабильности данной ткани вызывает снижение ответа – пессимум. Меры возбудимости. 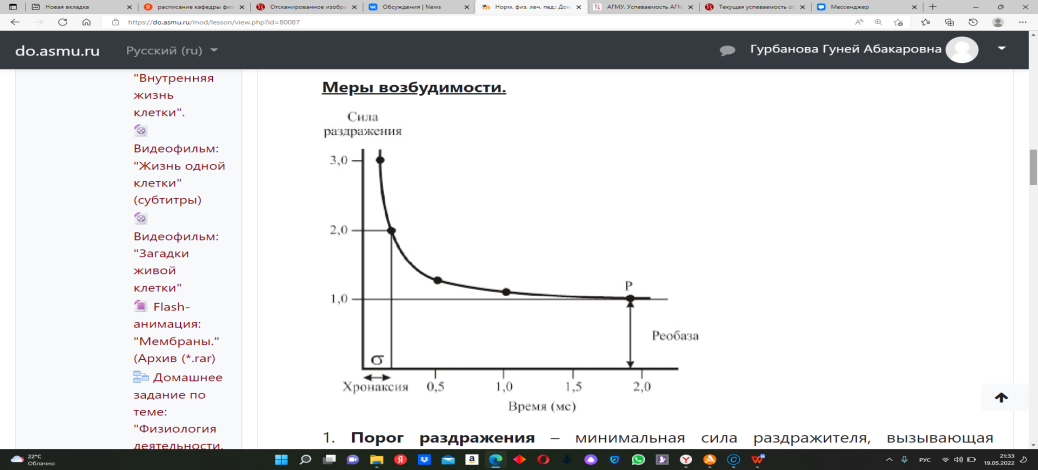 1. Порог раздражения – минимальная сила раздражителя, вызывающая возбуждение. 2. Реобаза – это минимальная сила постоянного тока, которая, действуя длительное, но определенное время, способна вызвать ответную реакцию. 3.Полезное время – это время, в течение которого должна действовать сила тока в 1 реобазу, чтобы вызвать ответную реакцию. 4. Хронаксия – это минимальное время, в течение которого должна действовать сила тока в 2 реобазы, чтобы вызвать ответную реакцию. Автоматия – это способность клетки самопроизвольно/спонтанно/ возбуждаться через определенные интервалы времени. Информационный блок № 2.Основные положения ионно-мембранной теории. 1. Электрические процессы в клетке возникают вследствие того, что мембрана обладает избирательной проницаемостью для ионов. 2. В покое мембрана проницаема для одних ионов, а при переходе в активное состояние – для других. 3. Электрические явления в тканях обусловлены неравномерным распределением ионов между цитоплазмой клетки и межклеточной жидкостью. 4. Избирательное перемещение ионов через мембрану изменяет ее электрическое состояние, что приводит к формированию новых видов электрических явлений в клетках и тканях. На уровне клетки регистрируется потенциал мембраны (ПМ) - разность потенциалов между наружной и внутренней поверхностью мембраны в каждый данный момент времени. В клетке регистрируют 2 вида потенциала мембраны (ПМ): потенциал покоя (ПП) и потенциал действия (ПД). Потенциал покоя (ПП) – это разность потенциалов между наружной и внутренней поверхностями мембраны в состоянии покоя,т.е. в покое мембрана поляризована. Происхождение ПП обусловлено: 1.Неравномерным распределением ионов калия и натрия между цитоплазмой и межклеточной жидкостью. В клетке – калия порядка 400 ммоль/литр, вне клетки – 10, соответственно натрияв клетке 50 и 460 вне клетки в состоянии покоя. 2.Избирательной проницаемостью клеточной мембраны в покое для калия. В покое - высокая проницаемость для калия, а для натрия в покое она практически отсутствует(небольшая). Постоянному выходящему калиевому току противодействует работа калиевой части калий-натриевого насоса, и в клетке возникает равновесное состояние между выходящим и входящим калиевым током. Это формирует равновесный калиевый потенциал /ЕК/ - разность потенциалов между наружной и внутренней поверхностью мембраны, если бы её создавал один вид ионов. Его величина описывается уравнением Нернста:
где: R - газовая постоянная, Т - абсолютная температура, F - число Фарадея, Ке - концентрация свободных ионов калия в наружном растворе, Кi - их концентрация в цитоплазме, n - валентность, ln- натуральный логарифм. Потенциал действия (ПД). Если на клетку нанести раздражение достаточной силы, клетка придет в новое, активноесостояние. Локальный ответ – при нанесении раздражения увеличивается проницаемость мембраны для натрия. Происходит уменьшение полярности мембраны по сравнению с исходным (с 70 мв до 40-50 мв). Эти изменения называются – пассивная или частичная деполяризация. Если силы раздражителя недостаточно, чтобы сместить ПМ до критического уровня, то происходит возращение ПМ к исходномууровню, т.е. к уровню потенциала покоя (ПП). Возникшие изменения ПМ называются локальный ответ. Критический уровень деполяризации (КУД) – это такая величина разности потенциалов (40-50 мв), при которой активируется большое количество потенциалзависимых быстрых натриевых каналов, проницаемость мембраны для натрия становится максимальной и перестает быть зависимой от силы раздражителя. Фазы ПД. 1) Деполяризация – восходящая часть ПД.Возникает лавинообразный входящий натриевый ток, который смещает потенциал мембраны до 0, а затем его силы хватает, чтобы изменить знак мембраны на противоположный (плюс 10-20 мв) – овершут или реверсия знака. 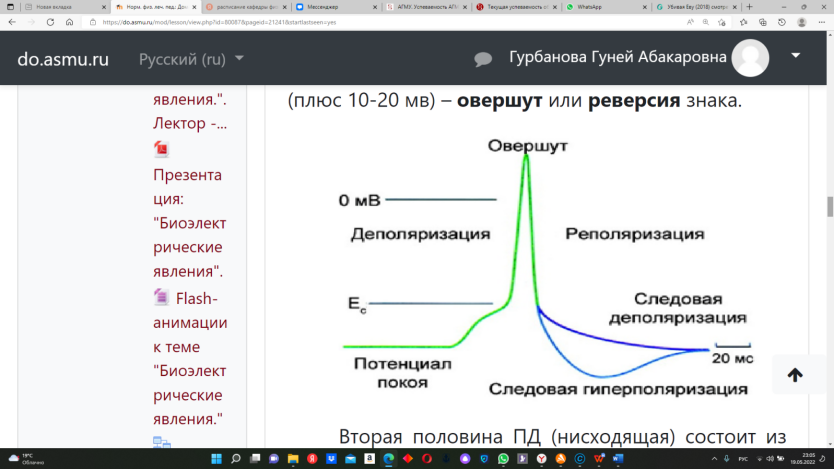 Вторая половина ПД (нисходящая) состоит из трех частей: 2. Реполяризация – нисходящая часть ПД от острия пика до КУД формируется быстро. От КУД до ПП медленнее – отрицательный следовой потенциал. 3. Гиперполяризация – ниже исходного уровня ПП - положительный следовой потенциал. На уровне ткани регистрируются следующие биоэлектрические явления: 1. В состоянии покоя: - токи повреждения - (ТП), токи градиента основного обмена (ТГОО). 2. В состоянии возбуждения: - токи действия (ТД). (ЭКГ, ЭЭГ, ЭМиоГ, ЭРетиноГ, ЭГастроГ, ЭГистероГ, т.е. в любых возбудимых тканях). Изменение возбудимости при формировании потенциала действия. Нормальная возбудимость в покое принимается за 100 %. Возбудимость характеризуется разностью между потенциалом мембраны (ПМ) и КУД. При увеличении этой разницы возбудимость увеличивается, а при снижении – уменьшается. 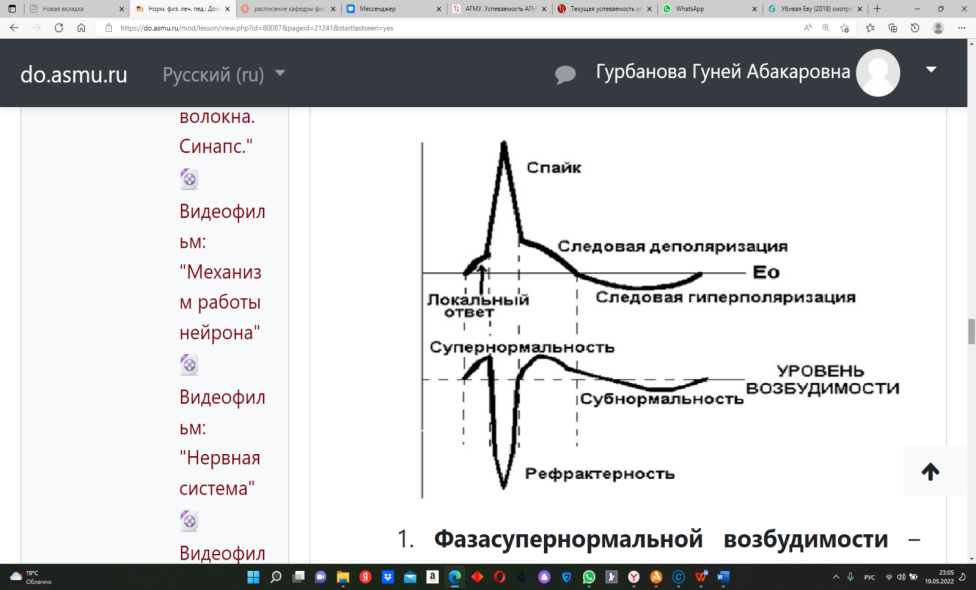 1. Фазасупернормальной возбудимости – начальное изменение возбудимости и отрицательный следовой потенциал, т.к. разница между ПМ и КУД уменьшается. 2. Фаза абсолютной рефрактерности – в момент достижения КУД возбудимость у клетки будет равна нулю. 3. Фаза относительной рефрактерности – по мере реполяризации будет происходить восстановление возбудимости. Но клетку могут возбудить только чрезвычайно сильные раздражители. 4. Фаза субнормальной возбудимости (ниже нормальной возбудимости) – это положительный следовой потенциал, так как расстояние до КУДа увеличено. Информационный блок № 3."Действие постоянного тока на ткань" Постоянный ток на ткань оказываетдва вида действия: 1. Возбуждающее действие 2. Электротоническое действие. 1. Возбуждающее действие оказывает ток пороговой силы и описывается тремя законами Пфлюгера: 1. При действии постоянного порогового тока на ткань возбуждение возникает только в момент замыкания цепи или в момент размыкания цепи, или при резком изменении силы тока. 2. Возбуждение возникает при замыкании под катодом, а при размыкании – под анодом. 3. Порог катодзамыкательного действия меньше, чем порог анодразмыкательного действия. При длительном действии сильного постоянного тока возникнет два состояния: 1.Под катодом – катодическая депрессия. 2.Под анодом – анодный блок. Главным признаком катодической депрессии и анодного блока являетсяснижение возбудимости и проводимости до нулевого уровня. Однако биологическая ткань при этом остается живой. 2. Электротоническое действие постоянного тока на ткань. Под электротоническим действием понимают такое действие постоянного подпорогового тока на ткань, которое приводит к изменению физических и физиологических свойств ткани. Различают: 1.Физический электротон. 2. Физиологический электротон. Под физическим электротоном понимают изменение физических свойств мембраны, возникающее под действием постоянного тока - изменение проницаемости мембраны,критического уровня деполяризации. Под физиологическим электротоном понимают изменение физиологических свойств ткани. А именно - возбудимости, проводимости под действием электрического тока. Кроме того, электротон разделяют на анэлектротон и катэлектротон. Анэлектротон - изменения физических и физиологических свойств тканей под действием анода. Каэлектротон - изменения физических и физиологических свойств тканей под действием катода. Анэлектротон– это снижение возбудимости ткани под анодом вследствие частичной гиперполяризации при действии на нее постоянного подпорогового электрического тока. Катэлектротон – это увеличение возбудимости ткани под катодом вследствие частичной деполяризации при действии на нее постоянного подпорогового электрического тока. Информационный блок № 1.Нейрон – структурно-функциональная единица нервной ткани. Физиологические свойства нейрона: 1. Рецепция. 2. Электрогенез. 3. Нейросекреция. Рецепция нейронов. Рецептор – специализированное образование для восприятия действия раздражителя. Выделяют: 1. Сенсорные рецепторы. 2. Клеточные химические рецепторы. Сенсорные рецепторы – это нервные окончания, способные воспринимать различные виды раздражения. Выделяют первично-чувствительные рецепторы – восприятие и преобразование информации (давление, температура) идет в чувствительном нейроне. Вторично-чувствительные рецепторы – специализированные клетки, которые передают информацию на афферентные окончания нейрона. Экстерорецепторы – воспринимают раздражения из внешней среды. Интерорецепторы– воспринимают информацию о состоянии внутренней среды и внутренних органов. Выделяют: проприорецепторы (в мышцах и сухожилиях); ангиорецепторы (в сосудистом русле); тканевые рецепторы. По адаптивным способностям: 1) слабо-адаптирующиеся 2) быстро- и полностью адаптирующиеся. Функционально рецепторы делятся на: 1) мономодальные – специфическим для них является только один раздражитель; 2) бимодальные – формируют два ощущения; 3) полимодальные – воспринимают больше 2-х раздражи-телей. Рецепторы могут быть: 1) Спонтанно-активные – постоянно генерируют импульсы. 2) Молчащие – посылают импульсы только когда имеются отклонения от нормальной деятельности. Интенсивность воздействия на рецепторы кодируется: 1) Числом потенциалов действия отрецепторы. 2) Числом рецепторов. Рецепторы передают информацию на нервные проводники. Электрогенез. Это формирование электрической активности нейрона. Выделяют: спонтанную активность (самопроизвольную) и вызванную активность (под действием раздражителей). По своей исходной активности все нейроны делятся: 1) нейроны со спонтанной одиночной активностью; 2) нейроны с «пачковой» спонтанной активностью; 3) Нейроны с групповой активностью. Кодировка информации в нейроне: 1) Неимпульсная – за счет изменения уровня потенциала мембраны и КУД. 2) Импульсная – за счет изменения частотных характеристик и конфигурации импульсов. 窗体顶端 窗体顶端 窗体底端 Информационный блок № 2.Нервные проводники обладают: возбудимостью и проводимостью. Мерой проводимости является скорость проведения возбуждения, чем больше диаметр проводника, тем больше скорость проведения возбуждения. Классификация нервных волокон. 1. По наличию оболочки: - миелиновые (мякотные); - безмиелиновые (безмякотные). 2. По скорости проведения возбуждения. Волокна группы А (миелинезированные): 1) А-альфа. Диаметр=13-22 мк; скорость проведения 70-120 м/с. 2) А-бета. Диаметр=8-13 мк; скорость- 40-70 м/с. 3) А-гамма. Диаметр=4-8 мкм; скорость проведения возбуждения 15-40 м/с. 4) А-дельта. Диаметр=1-4 мкм; скорость проведения возбуждения 5-15 м/с. Волокна группы В (миелинезированные) – 1-3,5 микрона, 3-8 м/сек. Преганглионарные вегетативные волокна. Волокна группы С(немиелинезированные) – 0,5 - 2,0 микрона, 0,5-3 м/сек. Постганглионарные вегетативные волокна. Законы проведения возбуждения 1.Закон двухстороннего проведения возбуждения. 2.Закон физиологической целостности нервного проводника. 3.Закон изолированного проведения возбуждения. 4. Закон бездекрементного проведения возбуждения. Механизмы проведения возбуждения по нервному волокну. В безмякотных волокнахвозбуждение проводится последовательно за счет разности потенциалов между возбужденным и невозбужденным участком. В мякотных волокнахвозбуждение проводитсяскачкообразно – сальтоторно. Парабиоз Парабиоз – означает "около жизни". Это явление последовательного снижения лабильности ткани вследствие действия парабиотических веществ. Фазы парабиоза: 1. Уравнительная. Одинаковый ответ на сильные и слабые стимулы. 2. Парадоксальная. На слабый стимул ответ регистрируется, на сильный – нет. 3. Тормозная. Нет ответа на стимулы различной силы. Это связано со снижением лабильности до НУЛЯ. Биологическое значение парабиоза 1. Показал, что явление смерти не мгновенно, существует переходный период между жизнью и смертью. 2. Этот переход осуществляется пофазно. 3. Первая и вторая фазы обратимы, а третья не обратимая. Эти открытия привели в медицине к понятиям – клиническая смерть, биологическая смерть. Клиническая смерть - это обратимое состояние. Биологическая смерть - необратимое состояние. Появилась новая наука – реаниматология. Информационный блок № 3.Синапс – структурно-функциональное образование, за счет которого возбуждение передается с нервного волокна на другую возбудимую клетку. Классификация синапсов: 1). Нервно-мышечный синапс. 2). Нервно-секреторный (нервно-железистый) синапс. 3). Нервно-нервный синапс: а). Аксо-соматический; б). Аксо-дендрический; в). Аксо-аксональный. По механизму передачи возбуждения в синапсе выделяют: 1.Химический (наиболее часто) 2.Электрический. Строение химического синапса: 1. Пресинаптическая мембрана.а) электровозбудима;б) хемоневозбудима;в) содержит медиатор в везикулах. Синапс - мономедиаторен. В области пресинаптической мембраны медиатор «упаковывается» в везикулы, которые в покое прочно связаны с цитоскелетом клетки специфическими белком – синаптосином. 2. Синаптическая щель (80 – 200 А). заполнена межклеточной жидкостью. По периферии существует специфический барьер, препятствующий выходу медиатора за пределы синапса. 3. Постсинаптическая мембрана, имеющая субсинаптическую мембрану (участок, обращенный в синаптическую щель). а) хемовозбудима б) электроневозбудима в) имеет большое число хеморецепторов к медиатору. Механизм передачи возбуждения через синапс1. Когда возбуждение приходит в область пресинаптической мембраны, она деполяризуется. Это активирует кальциевые потенциалзависимые каналы. 2. В клетку входит кальций, связывается со кальций-связывающим белком – синаптосином. При этом везикулы перемещаются к внутренней поверхности пресинаптической мембраны и прилипают к ней. 3. Активируется мембранный белок – синаптопорин, который выступает как переносчик медиатора через мембрану (экзоцитоз). Медиатор диффундирует в синаптическую щель. 4. Молекулы медиатора идут к постсинаптической мембране и образуют комплекс «медиатор–рецептор». Это вызывает активацию соответствующих рецепторуправляемых ионных каналов. Открываются Na-евые каналы – деполяризация как при формировании ПД. При взаимодействии медиатора с хеморецепторами возникают постсинаптические потенциалы. Секреция малых доз медиатора вызывает потенциал концевой пластинки, или миниатюрные постсинаптические потенциалы (со свойствами локального ответа). При массивном поступлении медиатора мембранный потенциал достигает КУД, что и приводит к возбуждению следующей клетки. Различают ВПСП – возбуждающий постсинаптический потенциал, ТПСП – тормозной постсинаптический потенциал. Возбуждающие медиаторы (напр. ацетилхолин, адреналин) вызывают активацию натриевых (реже - кальциевых) каналов и формируют входящий натриевый (либо кальциевый) ток, который вызывает возникновение частичной деполяризации, т.е возбуждающий постсинаптический потенциал (ВПСП). Тормозной медиатор (глицин, ГАМК) вызывает усиление выходящего калиевого тока или входящего тока хлора, т.е. вызывает гиперполяризацию. Это формирует тормозной постсинаптический потенциал (ТПСП). В синапсе известно два вида суммации: 1. Пространственная суммация 2. Временная суммация В дальнейшем комплекс «медиатор-рецептор» распадается (диссоциирует). Медиатор отсоединяется от рецепторов и разрушается ферментами (холинэстераза и т.д.), которые имеются в синапсе. Другой способ инактивации медиатора – аптейк – обратный захват пресинаптической мембраной. За счет этого синапс экономно расходует медиатор. |