Дріс 3 Жыныспен тіркескен белгілерді тым уалауды задылытары. арастырылатын сратар
 Скачать 65.51 Kb. Скачать 65.51 Kb.
|

|
Дәріс 3 Жыныспен тіркескен белгілердің тұқым қуалаудың заңдылықтары. Қарастырылатын сұрақтар: Г Мендель генетика негізінің бірі Гибридологиялық әдіс. Генетикалық символикасы.Моногибридті будандастыру. Мендельдің I және II заңдары. Мендельдің III заңы. Дигибридті будандастыру. Доминанттылқтың түрлері. Маңызды терминдер: ген, геноттип, фенотип, аллель, гомозигота, гетерозигота, ажырау, тұқым қуалаушылық, будандастыру, леталды, плейотропия, белгі. 1. Кәзіргі генетиканың тарихы 1900 жылы Э.Чермак, К.Корренс және Г.Де Фриздердің жеке белгілердің тұқым куалау зандарын тәуелсіз ашып, ген теориясының қабылдануынан басталады, бірақ олар осы зандарды 1865 жылы Г.Мендель ашқанын білмеген болатын. Мендель толық түсінікпен алға қойған мақсатқа сәйкестіріп тәжірибелеріне өте қолайлы объект - бұршақгың әр түрлі сорттарын қолданды. Бұршақ өздігінен тозаңданатын өсімдік болғандыктан, біріншіден, оның бірнеше таза түқымды сорттары бар; екіншіден, бұршақ гүлдері бөтен тозандардың енуінен жақсы қорғалған, мұның езі көбею процесін толық бақылауға мүмкіндік береді; үшіншіден, будандардың түқымдық қасиеті толық сактал-ғандыктан белгілердің тұқым куалауын бірнеше үрпактар бойы қадағалауға болады. Мендель тәжірибелерге мынадай талаптар қойды: 1) будандастыру үшін алынған ата-аналық формалардың зерттелгелі отырған белгілері қарама-карсы (альтернативті) болуы керек; 2) ата-ана-лық формалар таза линияға жатуы керек, ягни олардың әркайсысы өзара шағылысуда мәлім белгіні ұрпақган ұрпаққа әр уакытта беруі қажет; 3) будаңдар мен олардың ұрпақтарының тіршілік қабілеттілігі мен түқымдық қасиеті түракты болуы керек . Бұршақ осы талаптарға толық сәйкес келеді. Мендель осы объектіде гибридологиялық әдісті жете зерттеп, тұқым куалауды зерттеудің жаңа принциптерінің негізін салды. Мендель жүмысының баска бір ерекшелігі - сандық таддау: ол әрбір келесі ұрпақга зерттеліп отырған белгілері бойынша айырмашылығы бар дарактардың санын есепке алды. 2. Гибридологиялық әдісте белгілердің тұқым куалауын зерттеу үшін шағылыстыру схемасын күрастырады. Шағылыстыру немесе будандастыру көбейту белгісімен ( х ) таңбаланады да, бұл белгі ата-аналар арасына түсіріледі. Шағылыстыру схемасын жазган кезде алдымен ұрғашы, сонаң сон, "х" белгісінен кейін еркек жыныстың белгісі мен гендері жазылады. Үрғашы жыныс 9 (Шолпан планетасының символы) белгісімен, еркек жыныс d (Марс планетасының символы) белгісімен жазылады. Одан темен орналаскдн қатарда ата-аналар түзетін гаметалардың барлық типтері жазылады. Ата-ана Р әріпімен (лат.рагегпя- ата-ана), ал олардан алынған будандар Ғ өріпімен (лат.-үрпак) белгіленеді. Будандардың бірінші ұрпағы Ғ„ екінші үрпағы Ғ2 т.с.с. белгіленеді. Гибридологиялық талдауда будандастырудың ең қарапайым түрі - моногибридтік будандастыру. Бір-бірінен бір жұп белгілері бойынша айырмашылығы бар ата-аналық форманы будандастыру моногибридтік будандастыру деп аталады. Бүл будандастыруда дамуы аллельді гендермен аныкталатын тек бір ғана жүп белгілердің түқым қуалау заңдылығы зерттеледі. Ата-ананың басқа белгілері есепке алынбайды. Моногибридтік будандастыру тәжірибелерінің бірінде Мендель ас бүршактың тегіс және бұдыр дәнді сорттарын өзара будандастырды, нәтижесінде бірінші үрпактағы барлық будандар тегіс дәнді болып шықты, яғни сыртқы пішіні бойынша біркелкі. Түқым сортының тегіс пішінді белгісі бүдыр белгіге басымдылық жасағандай болды. Басқа белгілердің дамуын тежейтін белгі доминантты деп аталады. Мүндай белгілер Мендель бойынша доминантты түқым қуалау факторлары әсерімен аныкталады, шағылыстыру схемасында олар латын өліпбиінің бас өрпімен (А,В т.с.с) белгіленеді. Дамымай калган белгі рецессивті деп алады. Мүндай белгілердің түқым куа-лау факторлары латын әліпбиінің кіші өрптерімен (а,Ь т.с.с.) белгіленеді. 1909 жылы дания ғалымы В. Иогансен ғалымға "ген", "генотип" жөне "фенотип" үғымдарын енгізді. Ген - түқым қуалау бірлігі ген доминантты не рецессивті болуы мүмкін. Организм гендерінің жиынтығы генотип деп аталады. Зерттеліп отырған белгінің гендері бойынша генотип екі түрлі болуы мүмкін: гомозиго-талы немесе гетерозиготалы. Генотиптері бірдей гендерден тура-тын дарақгар (АА немесе аа) гомозиготалы деп, ал әр түрлі гендерден (Аа) түратын дарактар гетерозиготалы деп аталады. Генотип пен ортаның өзара әрекеттесу нәтижесінде калыптасқан ор-ганизмнің байқауға болатын белгілерінің жиынтығы фенотип деп аталады. Организмнің тіршілігі бойында оның фенотипі өзгеруі мүмкін, бірақ генотипі өзгермейді. Бірінші үрпақ будандарының біркелкілік ережесі ата-ана генотипі гомозиготалы (АА - тегіс дәнді фенотип, аа - бүжыр) болса ғана сақгалады. Бірінші үрпақ генотип бойынша да біркелкі: гетерозиготалы (Аа). Сонымен Мендельдің I заңын былай деп түжырымдауға болады: бір-бірінен карама-карсы белгілері (бір немесе бірнеше) бойынша айырмашылығы бар гомозиготалы ата-аналар формасын шағылыстырғанда барлық бірінші үрпақ бір-келкі фенотиппен сипатталады. Бірінші үрпақты өздігінен тозандандыру немесе өзара будандастыру арқылы екінші үрпақ будандарын алуға болады. Мендель тәжірибелерінде екінші үрпақ будандары дәнінің сырты тегіс те және бүдыр да болатындыгы аныкталады, өрбір бүр-шаккдпта екі пішінді де дөндер кездесуі әбден мүмкін. Саналған 7324 дәннің 5474-і (74,7%) тегіс, 1850-і (25,3%) бүдыр дәнді болып шыкты, мүның өзі 2,9 тегістің 1,1 бүдырға немесе жуық шамамен 3:1 арақатынасын береді. Мүндай арақатынас басқа белгілердің түқым қуалауын зертте-генде де байқалады: Ғ2-де доминантты өсімдіктер саны рецессивті өсімдіктер санынан шамамен үш есе көп болды. Сары түқымды бүршакты жасыл түқымды бүршақпен будандастырганда бірінші үрпақ будандары түгелімен сары ренді, ал екінші үрпақ будан-дарының 3,01 бөлігі (75,1%) сары, 0,99 бөлігі (24,9%) жасыл ренді болып шыкты. Осы бақылауларға негізделген Мендельдің II заңын былайша түжырымдауға болады: екінші үрпақта белгі белгілі бір арақатынас аралығында ажырайды, атап айтқанда доминантты белгісі бар дарактардың рецессивті белгісі бар дарактарға катынасы 3:1-ге тең. Рецессивті белгілердің бірінші үрпақга байқалмай, екінші үрпақта көрілуіне сүйеніп, Мендель түқым қуалаушылықтың дискреттілігін түжырымдайды, демек, организмнің кез келген белгісі жеке түқым куалаушы факторларға байланысты. Қарама-карсы белгілердің қалайша түкьім куалайтынын үғу үшін Г. Мендель түқым Бірінші үрпақ будандарының біркелкілік ережесі ата-ана генотипі гомозиготалы (АА - тегіс дәнді фенотип, аа - бүжыр) болса ғана сақталады. Бірінші үрпақ генотип бойынша да біркелкі: гетерозиготалы (Аа). Сонымен Мендельдің I заңын былай деп түжырымдауға болады: бір-бірінен карама-кдрсы белгілері (бір немесе бірнеше) бойынша айырмашылығы бар гомозиготалы ата-аналар формасын шағылыстырғанда барлық бірінші үрпақ біркелкі фенотиппен сипатталады. Р А А х аа Мейоз процесінің ашылуымен Мендельдің осы жорамалы толық цитоло-гиялық дәлелдеме алды: әрбір жыныс клеткаға гомологты (жүп) хромосомалардың тек бір сыңары ғана түседі, яғни жүп геннің біреуі, ал гаметадағы алуан түрлі гендердің қисындасуы кездейсоқ жүреді де статистика заңцылығына бағынады. Мендельдің III заңы - белгілердің тәуелсіз ажырауы Бұл заң полигибридтік будандастыруда ашылды. Мұндай будандастыруда бастапқы формалар екі және одан көп белгілері бойынша айырмашылыгы болады. Полигибридтік будандастырудың ең қарапайым түрі дигибридтік будандастыру, мұнда екі белгінің тұқым куалауын талдайды. Мендель ас бүршактың сары тегіс қабыкты сортын (ААВВ) жасыл бұдыр қабыкты сортымен (aabb) будандастырды, яғни олардың альтернативті екі белгісі -тұқымның түсі жөне пішіні жағынан айырмашылығы болды. Будандардың бірінші ұрпағында түқым сары түсті, оның қабығы тегіс болып шықты, демек, бастапқы ата-аналық формалар атал-ған екі белгі бойынша гомозиготалы болған. Бірінші үрпақтың генотипі дигетерозиготалы — АаВЬ. Өздігінен тозанданған бірінші үрпақ өсімдіктерін зерттеп, Мендель Ғ2 де фенотип бойынша төрт категорияға бөлінген 556 дән алды, атап айтқанда 315-сары тегіс (АВ), 108-жасыл тегіс (аВ), 101-сары бұдыр (АЬ) жөне 32-жасыл бүдыр (ab). Мендель 32 саннын арақатынас бірлігі ретінде алды, сөйтіп дигибридтік ажырау формуласын жалпы түрғыдан алғанда былайша өрнектеді: 9АВ :ЗАЬ:ЗаВ:1аЬ. Дигибридтік ажырауды талдай отырып, Мендель екі жүп түқым қуалау факторларының бір-біріне мүлде төуелсіз түрде түқым куалайтынын тапты. Сөйтіп, дигибридтік белгілердің ажырауын, бір-бірімен қабаттасып келген тәуелсіз екі моногибридтік ажырау деп карауға болады. Екінші үрпакта фенотиптердің жаңа үйлесім табуы (сары бүдыр қабыкты және жасыл тегіс кдбықгы) гендердің тәуелсіз түқым куалағанын сенімді түрде дәлелдейді. Әйтпесе F1-де тек бастапқы ата-ана фенотиптерінің (сары тегіс және жасыл бүдыр қабыкты) кдйталануын күту керек еді, бірақ іс жүзінде олай болмады. Гендердің төуелсіз таралуы үшін міндетті түрде мынадай шарт қойылады: аллель жүптарының әрқайсысы әр түрлі гомологты хромосомада орналасады. Сондыктан да мейоз кезінде аллельдің бір жүбы екінші жүбынан тәуелсіз, еркін күйде тарайды хромосомада орналасады. Соңдықтан да мейоз кезінде аллельдің бір жүбы екінші жұбынан төуелсіз, еркін күйде тарайды. 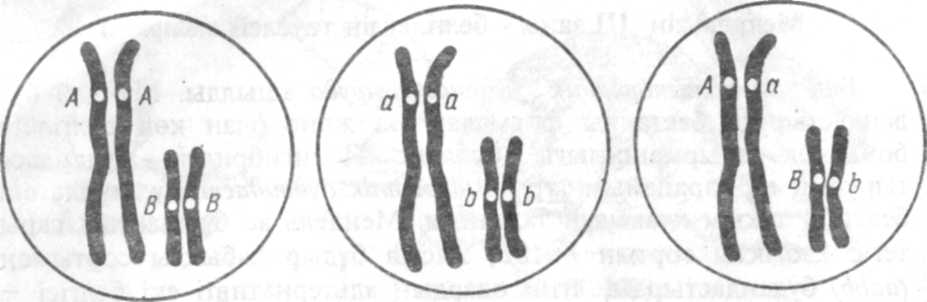 . Екі жұп аллельдің екі жұп хромосомада орналасуы. 1,2-бастапқы гомозиготалы формаларда; 3-дигетерозиготада. Сонымен Мендельдің III заңы жөнінде мынадай қорытынды жасауға болады. Әр гомологты хромосомада орналасқан түрлі гендер бір-біріне төуелсіз тұқым куалайды. Белгілердің тәуелсіз комбинациялануы туралы қорытындының дұрыстығын Мендель кері шағылыстыру арқылы дөлелдеді. Тригибридті будандастыруда - үш ген, тетрагибридтік будандастыруда - терт ген ер түрлі гомологты хромосомаларда орналасса, онда олардың тұқым қуалауы Мендельдің III заңына толық бағынады. Екі немесе бірнеше ген бір гомологты хромосомада орналасуы мүмкін, мұндай жағдайда олардың тұқым куалау заңдылығы езгеше болады (IV тарауда баяндалады). Белгілердің теуелсіз комбинациялану заңы мал селекциясында жиі қолданады. Мысалы, қазақгың ақ бас ірі кдра түкьімы жер-гілікті кдзақы сиырларын ағылшынның герефорд түқымымен шағылыстыру аркылы алынған. Осы түқымды қалыптастыруда сұрыптау жүмысы қазақы сиырдың жергілікті жағдайда ерекше бейімділігі мен ауруға тезімділігін герефорд түқымының жоғарғы еттілік сапасымен үйлестіру бағытыңда жүргізілді . Мендель бір не екі жүп белгілері бойынша айырмашылығы бар бүршақтарды шағылыстырып, олардың бірінші және екінші үрпақта түқым куалауын талдады, сонымен қатар будандарды бастапқы формалармен шағылыстыру (кері шағылыстыру) нәтижесін де зерттеді. Осы жүмыстардың нәтижесі Мендельге түқым куалаудың үш ережесін түжырымдауға мүмкіндік берді, кейін олар Мендель заңдары деп аталып кетті. Мендельдің I (Ғ 1 дің біркелкілік) жөне II (Ғ2-де белгілердің ажырау) заңдары 3 Доминанттылықгың түрлері. Моногибридтік будандастырудың екінші үрпағьгнда фено-типтің 3:1 аракдтынаста ажырауы белгінің толық доминант-тылығында байкалады. Алуан түрлі организмдердің көптеген бел-гілерінің түқым куалауын зерттеу бір белгінің екінші белгіден толық басым (доминантты) болмайтынын көрсетеді. Мүндай жағдайда "доминантты" және "рецессивті" белгілер не тендер туралы сірә айтуға болмайды. Осыған сәйкес толық доминанттылықтан баска толымсыз, кодоминанттылық және тыскары доминанттылыкты ажыратады. Толымсыз доминанттылык жануарлар мен өсімдіктерде жиі байкалады. Мүндай жағдайда бірінші үрпақ Мендельдің I заңына сөйкес генотип бойынша біркелкі, бірақ белгінің көрінісі аралық сипатта болады: гетерозиготалы (Аа) бірінші үрпактың әр түрлі гендері бір-біріне басымдық жасамайды. Екінші үрпакта фенотиптің ажырауы генотиптің ажырауына дөл келеді: 1:2:1, себебі АА жөне Аа генотиптерінің фенотиптік көрінісі өр түрлі болады. Мысалы, күлағы жоқ жөне күлағы үзын қойлар шағылыстырылса, Ғгде шүнақ үрпақ туады, ал екінші үрпактың аракатынасы 1 үзын күлақты (АА) : 2 шүнақ күлакты {Аа) : 1 күлақсыз (аа) болады (13-сурет). Осы жолмен андалузия тауығы қауырсынының көгіддір түсі, тауық кдуырсынының бүйралығы, паломино жылқысының алтын-сары түсі түқым куалайтыны дөлелдеңді. Жалпы толымсыз доминанттылықгы белгілердің қарапайым аралық көрінісі емес, жүп гендердің өзара ерекше әрекеттесу нәтижесі деп түсіну керек. Мысалы, кдра жөне ақ күстардан алынған андалузия тауығының көгілдір түсі жаңа фенотип пайда болғандай эсер береді. Алайда андалузия тауығы (Аа) шынында қара, бірақ оның қауырсы-нының боялуы төмен, сондыктан да көгілдір түстің әсерін береді. Кодоминанттылык - екі геннің де гетерозиготалы дарақ белгісін қалыптастыруға қатысуы. Кодоминанттылықгың накты үлгісі болып қан топтары мен организм белоктары типтерінің түқым куалауы саналады. Мысалы, ірі қарада гемоглобиннің (оттек таситын белок) бірнеше типі кездеседі. Бүлардың ішінде А және В типті гемоглобиндер жиі кездеседі. Гетерозиготалық (АВ) үрпақга гемоглобиннің осы екі типі болады. Кодоминанттылык көбінесе молекул ал ық деңгейде анықгалатын белгілерге тән, сонымен бірге ол кәдімгі деңгейде анықгалатын сапалы белгілерде байқалуы мүмкін. Бүған мысал ретінде шортгорн сиыры түгі түсінің түқым куалауын кдрастыруға болады . Қызыл (АА) жене ақ (аа) малдарды шағылыстыру аркьілы алынған бірінші үрпақ шұбар (Аа) түсті болып көрінеді: қызыл (А-геннің өсері) жөне ақ (а-геннің өсері) түктердің араласуы осындай көрініс береді. Ал екінші үрпакта қызыл, шұбар жөне ақ түстер-дің арақатынасы (1:2:1) генотиптік аракатынаска дөп келеді (1АА: 2Аа:1аа). Кодоминанттылықга басымдылық күбылысы төн емес екеніне кдрамастан бірінші үрпақ будандары бірдей болады, демек, Меңдельдің бірінші заңы бүл қүбылыс үшін де дөл. Тысқара доминанттылык, деп гетерозиготалы дарактар (Аа) белгілерінің дамуы жағынан гомозиготалы дарактардан (АА.аа) асып түсуін түсінеді. Бірінші үрпақ будандары даму көрсеткіштері жағынан ата-анадан асып түседі. Мүндай күбылыс гетерозис деп аталады: будаңдардың бірінші үрпага және кейбір түраралық. Әдебиеттер Б.К. Бегимкул Генетика А., 2000. Е.К. Меркурьева Генетика ., 1991 г. Ф.Айала . Кайгер Дж. Современная генетика I - III том М., Мир,1987 М.М.Гершензон, Основы современной генетики. Киев,1983 Н. П.Дубинин Горизонты генетики М.,Просвещение,1970 Н.П.Дубинин Общая генетика М., Наука,1986 Б.Льюин . Гены М.,Мир,1987 О.Солбирг. Популяционная биология и эволюция. М., Мир ,1982 Дәріс 4 Тұқым қуалаушылқтың молекулалық негіздері Генетикалық материалдың табиғаты. ДНҚ және РНҚ. Уотсон мен Ф.Крик ДНҚ молекуласы. Нуклеин қышқылдары: химиялық құрамы, құрылысы және синтезделуі. Негізгі терминдер пурин, пиримидин, ДНҚ, РНҚ, комлементарлық, нуклеотид , нуклеозид, тасымалдаушы РНҚ ,информациялық РНҚ. Қазіргі күндері геннің дезоксирибонукленин қышқылының (ДНҚ) бөлігі екені жалпыға мәлім. Осы бірегей нуклейн қышқылын 1868 ж Ф. Мишер клетка ядросынан (лат.нуклейн) бөліп ашқан еді. Кейін 1924ж биолог Р. Фельген ДНҚ-ның хромосома құрамында болатынын көрсетті. Бірақ, XX ғасырдың 20-30 жылдары ДНҚ молекуласы төрт мономерлі негіздер (аденинді,гуанинді, цитозинді, тиминді) қатаң қайталанатын жүйелі полимер болғандықтан генетикалық информацияның сақтаушысы бола алмайды деген пікір қалыптасқан болатын. ДНҚ-ның тұқым қуалаушылық құбылысындағы рөлін айқындауда, Ф.Гриффитстің 1928 ж пневмокк бактерияларында ашқан трансформация құбылысының маңызы зор. Дезоксирибозануклейн қышқылының негізгі құрылымдық бірлігі – нуклеотида. Кез-келген нуклеотид химиялық әр түрлі үш бөліктен құралған. Бірінші бөлігі – көмірсу дезоксирибоза. Нуклеотиданың екінші бөлігі – көмірсумен байланысқан пурин немесе пиримидин азотты негіздері. ДНҚ құрамына пурин негіздері – аденин (А) мен гуанин (Г) және пиримидин негіздері цитозин – (Ц) мен тимин (Т) кіреді. Үшінші бөлік – фосфор қышқылының қалдығы көршілес орналасқан нуклеотидтер байланысын қамтамасыз етеді. Организмнің түріне қарай ДНҚ-ның тұтас молекуласында нуклеотидтер саны – құбылмалы: бірнеше мыңнан бірнеше миллионға дейін. Э.Чарграфф ДНҚ молекуласына химиялық талдау жүргізіп, оның құрамындағы аденин мен тимин, гуанин мен цитозин мольдер жағынан бірдей екендігін көрсетті (Чаргафф ережесі) Демек, кез келген ДНҚ құрамын (ΣА+ΣТ):(ΣГ+ΣЦ) арақатынасымен сипаттауға болады. 1953 ж Дж Уотсон мен Ф.Крик ұсынған модель бойынша ДНҚ молекуласы қос тізбектен құралған.Әдетте бір тізбектегі аденинге екі сутекті байланыс арқылы екінші тізбектегі әрқашанда аденин тиминге, гуанинге, цитозинге сәйкес келеді (А=Т; Г=Ц). Осындай бірін-бірі толықтыра алатын, яғни өзара сәйкес келетін тізбектер комплементарлы деп аталады.Спиральдағы (оралмадағы) әрбір пурин не пиримидин негізі дезоксирибозамен байланысып нуклеозида құрайды. Нуклеозидалар бір-бірімен фозфодиэфир байланысы арқылы қосылады. ДНҚ репликациясы . Митоз немесе мейоз жолымен бөлінер алдында клетканың ДНҚ-сы екі еселенеді (репликацияланады). Бұл арада ДНҚ –ның өзін-өзі синтездей (автосинтез) алатын бірден-бір химиялық зат екендігін атап айту керек. ДНҚ секілді, РНҚ –ның негізгі құрылымдық бірлігі – нуклеотид: РНҚ-да 4 түрлі нуклеотидтер құралған. ДНҚ құрамында көмірсу дезоксирибоза болса, ал РНҚ- да рибозада 2- көміртек атомына гидроксил тобы (ОН), ал дезоксирибозада сутегі атомы (Н) жалғасқан. Клетка РНҚ-сы үш түрлі – информациялық (иРНҚ), тасмалдаушы (тРНҚ) және рибосомалық (рРНҚ)- топқа бөлінеді.Үшеуі де ДНҚ –да тікелей синтезделеді. Ал иРНҚ-ның ДНҚ-да синтезделу процесі транскрипция деп аталады.Жаңадан синтезделіп жатқан и РНҚ нуклеотидтері ДНҚ нуклеотидтеріне комплементарлық ережеге сәйкес қосылады, ал процесс РНҚ – полимераза ферментінің қатысуымен өтеді. Информациялық РНҚ-ның ең қысқа молекуласы шамамен 300 нуклеотидтен құралған. Клеткадағы барлық РНҚ-ның 80%-тін құраайтын рибосомалық РНҚ-ны кейбір хромосомалардың ядро ұйымдастырушы бөліктерінде орналасқан ерекшелі гендер анықтайды.Рибосома массасының 60%-і рРНҚ-дан тұрады. Тасымалдаушы РНҚ молекуласы үлкен емес:ұзындығы 70-80 нуклеотид.Ол иРНҚ-мен қатар белок синтезіне қатынасады: рибосомаға амин қышқылдарын тасымалдайды. Информациялық РНҚ молекуоасындағы тізбектен триплеттер негіздерінің полипептидтік тізбектегі амин қышқылдар қатарына аударылу механизмі трансляция деп аталады. Трансляция 3 стадияға бөлінеді: инициация, элоигация, терминация . Инициация- синтездің бастапқы кезеңі Элонгация- өсіп жатқан полипептидтік тізбек құрамына амин қышқылдарының тізбектеліп қосылуы. Терминация- синтездің соңы –УАА, УГА немесе УАГ (стоп - кодондар). Әдебиеттер С.Ж. Стамбеков. Жалпы генетика. А., Ана тілі 1993. Б.К. Бегимкул Генетика А., 2000. Е.К. Меркурьева Генетика ., 1991 г. Е.К. Меркурьева,З.В.Абрамова и др. Генетика., М.В.О. «Агропромиздат»,1991 Е.К.Меркурьева, Г.Н.Шангин-Березовский. Генетика с основами биометрии., М,Колос 1983 Ф.Айала . Кайгер Дж. Современная генетика I - III том М., Мир,1987 М.М.Гершензон, Основы современной генетики. Киев,1983 Н. П.Дубинин Горизонты генетики М.,Просвещение,1970 Н.П.Дубинин Общая генетика М., Наука,1986 Б.Льюин . Гены М.,Мир,1987 О.Солбирг. Популяционная биология и эволюция. М., Мир ,1982 Дәріс 5 Тұқым қуалаушылықтың хромосомалық теориясы. Белгілердің тіркес тұқым қуалауы. Кроссинговер. Рекомбинациялар және оның түрлері. Гендердің тізбектеле орналасуы. Хромосомалық генетикалық картасы және оны құрастыру принциптері. Маңызды терминдер: кроссинговер, хиазма, триангуляция, морганида гетерогаметалы, диплоидты дарақ. Геннің жэне хромосоманыц гамета тузіуі мен урыктану кезеніңдеп тертібінің параллельдіпне суйене отырып, 1902 ж АБШ галымы В.Сэттон мен немю биологы Т. Бовери тендер хромосомада орналасады деген идеяны алгаш рет болжады. Бул идея тукым куалаушылыктың хромосомалык теориясыньң бастамасын салды. Осы кезден бастап хромосоманы тексеру генетикалык зерттеулермен тыгыз байланысты. Гендердің хромосомалардан анагурлым кеп болу керектігің айкын тусіне отырып, В.Сэттон гендердің тіркесу кубылысы туралы болжамды да бірінші болып айтты. Болжамныц гылыми делелді 1910 жылы АБШ-тыц атакты генетип Томас Хант Морган усынды. Ол кішкене саргыш-коңыр жеміс шыбыны - дрозофилага тәжі рибелер журпзді, Бул шыбынньң тобын жазда жене ерте кузде жерге тускен жене шіріген жемштер айналасынан керуге болады. Дрозофила - генетикалык зерттеу-лерде жиг кодцанылатын жене ете колайлы объект, себебі бул шыбынньң генерация уакыты ете аз (әрбір 10-15 кунде жаца урпак береді, ал урпагыныц саны кеп (әрбір будандастырудан (Нрнеше жуз) жене шыбын лабораториялык жагдайда оңай өсіріледі. Будан баска, оның кариотипің терт-ак жуп хромосомалардан (2п=8) туруы да генетикалык зерттеулер ушш айтарлыктай мацызды. Т.Х. Морган дрозофила шыбынында дигибридтік жене одан курделі шагылыстыру нетижелерің талдай отырып, тукым куалаушылыктыц хромосомалык теориясыньң непзің салды. 2. Кез келген организмге ген жене хромосома деңгейінде сандык тургыдан талдау жасасак, гендердің хромосомалардан елде-кайда кеп екенің байкар едік. Дрозофиланын терт-ак хромосомасы бар, ал оньң зерттелген гендерінің саны мьңнан асады. Уй хайуандарында хромосома саны 80-нен аспайды (уйректе 2п=80, туйеде 2п=74 т.с.с), ал олардьң кез келгенңде тендер саны-ондаган мыц (10-50 мыц). Осыган қараганда, ербір жуп хромосомада бір емес, бірнеше мыц аллельдер орналасуы керек. Ал хромосоманыц келеа урпакка біртутас тукым куалайтын элемент ретінде бершетшш ескерсек, окда бір хромосомада орналаскан гендер тіркесіп тукым куалайды, демек, ер турлі гендер, Мендельдің III заңындай, бір бгрмен теуелсіз турде тукым куаламайды. Өйткені бул зац бойынша олар ер турлі жуп хромосомаларда орналасуы керек еді. Белгілердің тіркес тукым куалау кубылысын алгаш рет 1906 жылы агылшын генетиктері В.Бэтсон мен Р.Пеннет хош шел бур-шакка дигибриидпк будандастыру журпзу нетижесінде ашты. Олар гулі кан кызыл, тозаңы сопак өсімдікті кызыл гулді тозаңы домалак өсімдікпен будандастырганда F1, біркелі (гулі - кан кызыл, тозаңы - сопак) болып шыкты, ал F2-де классикалык, 9:3:3:1 аракатынастьң орнына 11:1:1:3 аракатынасы алынды. Бул кездей-сок, нетиже, Мендель зандарына кдйшы келедг. Оныц шешімің 1910-1915 жыддары Т.Х. Морган жене оньщ шеюрттері А. Стертевант, К.Б Бриджес жене Г. Мюллер дрозофилада тапты. 3. Морганньң дрозофилада жургізген дигибридгтік шағылыстыруын карастырайык. Канаты жетіс сур шыбын мен канатты шала кара шыбынды будандастырганда F1-де барлык дрозофила жетік канаты, сур денелі болып шыкты. Осыган орай дрозофиланыц генотипі зерттеліп отырған екі жұп бойынша былайша белгіленеді: V - жетік қанат, V - шала қанат және В - денесі сур тусі, Ь - денесі кара тусі. Одан ері, осы Е, - деп дигетерозиготалы аталык шала канатты аналык кара шыбынмен шагылыстырылса ягни талдаушы шагылыстыру журпізілсе (У УуВЬ х 9 ууЬЬ), Мендель занына сейкес куткендегідей әркайсысына 25%-тен келесі төрт турлі фенотипің (1 жетік канатты сур : 1 жетік канатты кара : 1 шала канатты сур : 1 шала канатты кара) орнына шыбынның екі фенотипі ғана ягни 50% жепк кднатты сур жэне 50% шала кднатты кара шыбындар алынды (21 а,э-сурет), Демек, жетік канаттылык пен сур дене тусі, шала канаттылык пен кара дене тусінің тендері бірігіп тукым куалайды екен ягни дене тусі мен канат формасын аныктайтын тендер бір жуп хромосомада орналасады, олар урпакққа тіркесіп беріледі деп тужырымдауга болады. Сондыктан да, сперматогенез кезінде жуп хромосомалардьщ эрқайсысы жеке-жеке жыныс клеткага ажырайтындыктан, дигетерозиготалы біришл урпак гаметалардыд ею турш гана тузеді: біршіШСі - УВ. еюнипа — уЬ (сызыкшамен хромосома белпленген).Сонымен кері шагылыстырудан алынган урпактар бастапкы ата-аналар фенотипш толык (100%) кайталайды. Бул толык тіркес қубылысыдеп аталады. Талдаушы шагылыстыруга F1-ден аналык шыбын алынса, Р, негізінде толык тіркестегідей фенотиптің екі типі емес, төрт типі, ягни жетік қанатты сұр, жетік канатты кара, шала канатты сур жене шала кднатты кара типтер алынады, бірак олардьщ ажырауы Мендельдщ III зацындагыдай тен аракдтынаста (1:1:1:1) емес Дрозофила шыбындарыньщ 41,5%- жетік канатты сур, 41,5%- шала кднатты кдра, 8,5%-іжетік қанатты кара жене 8,5%-і шала кднатты сур болып шыкты. Бул арада да біз бастапкы белплердщ кебшесе біріпп тукым куалайтынын кереміз, бірак олардьщ тіркесіп тукым куалауы толык емес (83%). Толык емес тіркес себебі тусіну ушің белгілердің жаңа комбинациясы (сур шала каналы жене кара жетәк канатты) қайдан пайда болады деген сураққа жауап беру керек. Ол ушің мейоз процесінің бірінші бөлуіндеп хромосомалардың өзгеру тәртібін еске тусіреік. Мейоздыц бірінші профазасын зигонема стадиясында бивалент гомологты хромосомалар бір-біріне тартылып, конюгацияланады да, диплонема стадиясында, керісінше хромосома жіпшілері бір-бірінен итеріліп, гректің хи әріпіне /х/ уксас фигура тузіледі, ол хиазма деп аталады. Буны алгаш рет 1909 жылы бельгия цитологы Ф.Янсенс саламандра сперматогенезш зерттеген кезде байкдган болатын. Янсенс бакылауына суйене отырып, Морган хиазма тузілуу барысында гомологты хромосомалар өз беліктерімен ею жакты алмасады деген жорамал айтты. Гомологты хромосомалар беліктерінің алмасуы кроссинговер деп аталады. Бөліктердің алмасуы төрт хроматидадан куралган бивалентп хромосомадагы екі хроматиданыц (гомологтыц ер жубыныц біреуі) узілуінен басталады, одан сон, олар узшу нуктелер^нде алмаса косылғаннан кейш мейоздың келесі фазаларында ер турлі гаметаларға ажырайды. Гомозиготалы дарактыц (У\>ВЬ) б1ршцп хромосомасында V жене В.кт, екшпианде - V жене Ь гені орналасса, кроссин-говераз гаметалардыц екі типі ( УВ гендері жене уЬ - гендері бар) тузшедь Ал осы гендер арасында кроссинговер процесі болса, онда бул даракта тагы УЬ жене VВ гаметалары тузіледі Кроссинговерсіз тузілген гамета кроссоверссіз деп, ал кроссинговер нетижесінде тузілген гаметалар кроссоверлі деп аталады. Кроссоверлі гаметадан пайда болган организм кроссовер-немесе рекомбинат деп аталады, ал хромосомалардыц айкасуы нәтижесінде алмаскан гендер рекомбинатты__ деп аталады. Сөйтіп, кроссинговер генетикалык өзгегіштіктің маңызды көзі болып табылады. Жалпы хромосомалар айкасуының эр уакытта журетшш ес-керсек толык емес тіркесу организмдердщ барлык топтарында кецшен тараган, ал оныц дрозофила шыбыныныц аталык жынысында байкалмауы ережеден тыс, ете сирек кездесетің қубылыс. Будан баска толык тіркесу гендердің плейотропты эсерімен имитациялануы мумкін. Хромосомалар мейоздыц пахинема стадиясында узынша келген қурылым болып керінеді. Гомологты жуптарда бірнеше хиазмалар тузілу мумкін. Буныцңезі мейоз кезінде жуп хромосомалар арасында тек дара кроссинговер емес, кос жене одан да курделі хромосомалар айкасуыныц журетшднш керсетеді. Хромосомалар айкасуыныц жишп гендер арасындагы кашык-тыккд байланысты. Гендердщ аракэшыктыгы улгайган сайын кроссинговер жишп арта туседі, ал гендер арасы жакындаган сайын, хромосомалардыц айкасуы темендейдь Осыган байланысты Морган гендер аралыгыныц кашыктыгын кроссинговер жишп аркылы табуга болатынын усынды: х=(а+с)/п і00 Мунда, х-гендерінің аракашыктыгы немесе кроссинговер жийлігі (%); а - бірінші кроссовер тобындагы дарактар саны; с- екінші кроссовер тобындагы дарактар саны: п- тэжхрибедеп урпактар саны. 1% кроссинговерге тец гендер аракашыктыгы морганидадеп те аталады. 4. Морганнын жас шәкіртерінің бірі - Альфред Стертевант өзінің дипломдык жумысын орындау кезінде кроссинговер жиілігі гендердің хромосома бойында тізбектеле орналасатындыгын керсетеді деп бірші болып уәж айтты. Уш ген арасындагы өзара катынасты зерттей келе ол бірінші мен екінші, екінші мен ушінші, бірінші мен ушінші гендер арасындагы кроссинговер жиілігі не олардьщ косындысына, не айырымына тен екеннің керсетті. А жене В гендерінің аракашыктыгы, мысалы, 17 морганидага, ал В мен С гендерінің аракдшыктыгы - 10 морганидага тен болса, онда А жэне С гендерінің аракашыктыгы АВ жэне ВС кдшыктыктардьщ косындысына (АС-АВ+ВС=27 морганида) не айырымына {АС=АВ-ВС=7) тец , Бул А,В жэне С тендері хромосомада пзбектелш орналаскан жагдайда гана мумюн. Ягни олардьщ хромосомада орналасуы бір тузудеп уш нуктенщ ара-кдшыктыгыньщ геометриялык зандылыгына сай келед1 Осы ереженің дурыстығын б1р хромосомада орналаскан кептеген ушпк гендерде журпзілген тексерулер делелдеді Гендер арасындагы кроссинговер жишпнщ талдау аркылы хромосомалардын генетикалык картасы жасалды. Хромосомалардын генетикалық картасы ө дегеніміз гендердің хромосомадагы орналасу нуктелерінің улісі. Геннің хромосомадагы орнын табу ушің ең алдымен, оның кандай хромосомада орналасканын аныктау керек. Бір хромосомада орналаскан гендер тіркесу тобынкурайды. ТТркесулер тобынын саны эр уакытга турдщ хромосомаларыньщ гаплоидты санына тен болады. Мысалы дрозофиладагы хромосомалардын гаплоидты саны мен пркесулер тобы 4-ке, койда - 27-ге, жылкыда - 32-ге тен. Эдетте гендердщ аракдшыктыгын аныктау ушш бхр мезгшде ен аз дегенде уш ген арасындагы кроссинговер жишп есептелшедх. Триангуляция деп аталатын бул екі гендердің хромосомадағы орналасу тертібің аныктауга мумкіндігін береді Кәзіргі кезде көптеген организмдер ушін хромосомалардын генетикалык картасы курылды. Дрозофиланьң, тышкдннын жене кейбір микроорганизмдердің генетикалык картасы толык жасалды, кейбір есімдіктер (хош шел буршак, жугері, кызанак) жэне адам хромосомалары генетикалык картасын қурастырылуы ойдагыдай іске асуда. Мал тулпегері хромосомаларынын картасын курастыру енді ғана басталды. Уй кояны мен тауыкгң кейбір хромосомалары карталанды. Әдебиеттер С.Ж. Стамбеков. Жалпы генетика. А., Ана тілі 1993. Б.К. Бегимкул Генетика А., 2000. Ф.Айала . Кайгер Дж. Современная генетика I - III том М., Мир,1987 М.М.Гершензон, Основы современной генетики. Киев,1983 Дәріс 6 Жыныс генетикасы Жыныстың анықталуы және тұқым қуалауы. Жыныстар арақатынасы. Жыныспен тіркескен белгілердің тұқым қуалауы. Жыныстың дозалық қайтарымы Жыныспен шектелген және жынысқа тәуелді белгілердің тұқым қуалауы. Маңызды терминдер: аутосома, гомогаметалы, гетерогаметалы, диплоидты дарақ, фримартинизм. Жыныс- организмнің белгілері мен қасиеттерінің ерекше жиынтығы жыныс, организмнің кез-келген басқа белгілері сияқты генетикалық жолмен анықталып, тұқым қуалайды. Жануарлардың әр түрлі жыныстарын цитогенетикалық зерттеу олардың хромосомалар құрамында айырмашылық болатынын дәлелдейді. Айырмашылық хромосомалардың бір ғана жұбында байқалады, олар жыныс немесе гетерохромосомалар, кейде гомосомалар деп аталады. Ұрғашы және еркек жыныстардағы басқа бірдей хромосомалар аутосома деп аталады. Сүтқоректі жануарларда ұрғашының кариотипі ХХ- әріпімен белгіленетін бірдей жыныс хромосомалардан тұрады. Еркекте жұп жыныс хромосомалардың бірі ұрғашылардікімен ұқсас (Х), ал екіншісі әдетте кішірек болады және У- әріпімен белгіленеді (ХУ). Бірдей жыныс хромосомалары (ХХ) бар жыныс гомогаметалы деп аталады, ал әртүрлі жыныс хромосомалары (ХУ) бар жыныс гетерогаметалы деп аталады. Құста, көбелекте және кейбір қосмекенділер мен балықта жыныстың хромосомалық анықталуы керісінше: гомогаметалы жыныс болып еркек-ZZ, гетерогаметалы болып ұрғашы ZW саналады. Ақырында гапло- диплоидия деп аталатын жынысты анықтаудың мүлде басқа механизмі ара мен құмырсқада кеңінен тараған. Бұл насекомдардың жыныс хромосомалары жоқ: ұрғашы (патша), және жұмысшы аралар- диплоидты дарақтар (2n=32), ал еркек аралар (трутеньдер)- гаплоидты (n=16). Жыныс организмнің басқа күрделі белгілері сияқты, тек жыныс хромосоманың гендерімен ғана емес, сонымен қатар аутосомадағы көптеген гендердің әсерімен анықталады және оның дамуы сыртқы ортаның әсерінеде байланысты. Осыған байланысты, көптеген ғылыми деректер организмнің бисексуальдық немесе қосжыныстылық қабілетті екендігін дәлелдейді, яғни организм белгілі бір жағдайларда не ұрғашы не еркек жынысқа дамуы мүмкін. Ал, оның генетикалық тұрғыдан қосжыныстылығын кез-келген организмнің генотипі ұрғашы және еркек жыныстылардың гендерінен қалыптасатындығынан түсінуге болады. 1919 ж. К.Бриджес жынысты анықтаудың теңгерімдік (баланстық)теориясын ұсынды. Бұл теория бойынша дрозофиланың жынысы Х-хромосома саны мен аутосома (А) жиынтығы арасындағы арақатынаспен (Х/А) анықталады. Х/А=1 болса, қалыпты ұрғашының, Х/А=0,5 болған жағдайда қалыпты еркек шыбынның фенотипі дамиды, ал Х/А арақатынасы басқа шамаларға тең болған жағдайларда шыбын жынысының дамуы өзгереді. Организмнің интерсексуальдығын ірі қарада кездесетін фримартинизм құбылысынан көруге болады. Х және У немесе Z және W жыныс хромосомаларында тек белгілі бір жыныстың дамуын бақылайтын гендер ғана емес, сондай-ақ организмнің көптеген басқа белгілерінің де анықтайтын гендер бар. Гендері жыныс хромосомаларында орналасқан белгілері жыныспен тіркескен белгілер деп аталады. Жыныспен тіркескен белгілердің тұқым қуалау гендер аутосомада орналасқан белгілерден өзгеше. Мұндай тұқым қуалауда кәдімгі мендельдік арақатынас байқалмайды. Мұның негізгі себебі Х пен У және Z пен W хромосомаларының өзара мөлшерінің әртүрлі болуы ғана емес, сонымен қатар олардың генетикалық активтілігінің айырмашылығында қазіргі кезде Х және Z хромосомаларда орналасқан бірқатар гендердің У және W хромосомаларында жоқ болуы қағида ретінде қаралады. Жыныспен тіркесіп тұқым қуалау құбылысын алғаш рет Морган дрозофила шыбыны көз түсінің ұрпаққа тұқым қуалауын реципрокты будандастыру нәтижесін зерттеу барысында ашты. Реципрокты будандастыру дегеніміз- генотипі әртүрлі дарақтарды екі бағытта шағылыстыру: ♀Х(А) Х(А) x ♂ Х(а)У және ♀Х(а) Х(а) x ♂ Х(А)У Жыныспен тіркескен белгілердің тұқым қуалауындағы тағы бір ерекшелік – еркек жыныс хромосомаларының ұрпаққа берілуі. Аналық шыбынның Х-хромосомасы келесі ұрпаққа кездейсоқ берілсе, аталық шыбынның Х-хромосомасы келесі ұрпаққа кездейсоқ берілсе, аталық шыбынның Х-хромосомасы тек ұрғашы ұрпаққа тұқым қуалайды. Сондықтан да Х-хромосомада орналасқан гендер анықтайтын белгілер айқас тұқым қуалайды. Хромосомалар диплоидты болғанымен гетерогаметалы еркек жынысты ген жалғыз күйде болады. Мұндай генотипі бар организм гемизигота деп аталады. У хромосомада әкесінен тек ұлдарына ғана тұқым қуалайтын шиф. Беріледі. Бұл гаплоидтық тұқым қуалау деп аталады. Бір ғана жыныста дамитын белгілер жыныспен шектелген деп аталады. Мысалы, құстың жұмыртқалағыштығы мен сүтқоректі жануарлар сүттілігінің гендері екі жыныста да болғанымен, аталған белгілер тек ұрғашыда байқалады. Аутосомада орналасқан кейбір гендердің фенотипте байқалуы жынысқа байланысты, олар жынысқа тәуелді белгілер деп аталады. Бұған мысал ретінде қойдың мүйізділігінің тұқым қуалауын алуға болады. Әдебиеттер С.Ж. Стамбеков. Жалпы генетика. А., Ана тілі 1993. Б.К. Бегимкул Генетика А., 2000. Н. П.Дубинин Горизонты генетики М.,Просвещение,1970 Н.П.Дубинин Общая генетика М., Наука,1986 Б.Льюин . Гены М.,Мир,1987 О.Солбирг. Популяционная биология и эволюция. М., Мир ,1982 7 дәріс Жынысты анықтаудың баланстық теориясы Организмнің бисексуалдығы Жынысты ерте анықтаудың генетикалық әдістері Партеногенез: қолдан және табиғи. Маңызды терминдер бисексуалды, фримаринизм, анеуплоидия, полиплоидия, андрогенез, гиногенез 1. Организмнің интерсексуальдығын ірі қарада кездесетін фримартинизм құбылысынан көруге болады. Х және У немесе Z және W жыныс хромосомаларында тек белгілі бір жыныстың дамуын бақылайтын гендер ғана емес, сондай-ақ организмнің көптеген басқа белгілерінің де анықтайтын гендер бар. Гендері жыныс хромосомаларында орналасқан белгілері жыныспен тіркескен белгілер деп аталады. Жыныспен тіркескен белгілердің тұқым қуалау гендер аутосомада орналасқан белгілерден өзгеше. Мұндай тұқым қуалауда кәдімгі мендельдік арақатынас байқалмайды. Мұның негізгі себебі Х пен У және Z пен W хромосомаларының өзара мөлшерінің әртүрлі болуы ғана емес, сонымен қатар олардың генетикалық активтілігінің айырмашылығында қазіргі кезде Х және Z хромосомаларда орналасқан бірқатар гендердің У және W хромосомаларында жоқ болуы қағида ретінде қаралады. Жыныспен тіркесіп тұқым қуалау құбылысын алғаш рет Морган дрозофила шыбыны көз түсінің ұрпаққа тұқым қуалауын реципрокты будандастыру нәтижесін зерттеу барысында ашты. Реципрокты будандастыру дегеніміз- генотипі әртүрлі дарақтарды екі бағытта шағылыстыру: ♀Х(А) Х(А) x ♂ Х(а)У және ♀Х(а) Х(а) x ♂ Х(А)У Жыныспен тіркескен белгілердің тұқым қуалауындағы тағы бір ерекшелік – еркек жыныс хромосомаларының ұрпаққа берілуі. Аналық шыбынның Х-хромосомасы келесі ұрпаққа кездейсоқ берілсе, аталық шыбынның Х-хромосомасы келесі ұрпаққа кездейсоқ берілсе, аталық шыбынның Х-хромосомасы тек ұрғашы ұрпаққа тұқым қуалайды. Сондықтан да Х-хромосомада орналасқан гендер анықтайтын белгілер айқас тұқым қуалайды. Хромосомалар диплоидты болғанымен гетерогаметалы еркек жынысты ген жалғыз күйде болады. Мұндай генотипі бар организм гемизигота деп аталады. 2. Жынысты ерте анықтаудың генетикалық әдістері. Жынысты анықтаудың кейде реті келеді. Мысалы бройлерлік (ет үшін) балапандарды өсіру үшін тек әтештерді пайдаланады. Бірақ ерте күннен шібилермен қораздарды айыру өте қиын. Бұл мәселені шешу үшін 20-30 жылдардың өзінде жыныспен тіркес тауықтардың қауырсын бояуларын бір күндік балапандардың анықтау үшін қолданған. Мысалы, "алтын шашақ" әтештерді күміс түктес тауықтармен шағылыстырған. Бұлардан бірі сары, екіншісі ақшыл-жасыл балапандар шыққан, біріншілері – шібилер, екіншілері – қораздар. Жыныспен тіркес белгінің тұқым қууын пайдаланып, Англияда Пеннет тауықтың камбар тұқымын шығарған. Бұл тұқымның жаңа шыққан балапандарының әтештері әлде қайда ақшыл түсті, ал шібилерінің түбіті қара теңбілді болған. А.С.Серебровский тауықтардың жолақтығын зерттеп, оларды жыныс хромосомасында орналасқан геннің анықтайтынын ашқан. Сондықтан белгілі бір шағылыстыруларға гомозиготалы әтештерде ол түс өте сирек кездеседі. Шағылыстырудың басқа варианттарында барлық әтеш балапандардың желкесінде ақ дақ, ал шібилерде ол болмаған немесе әтештерде қара дақ болса, шібилерде ол болмаған. Бұл айырмашылықтар X – жыныс хромосомасында орналасқан доминантты B – генімен анықталады. Тұқым қуалайтын түс өзгешелігі ұрғашы ұрпақтарында да табылған, бір жынысы – ақ, екіншісі – сұр. Қазіргі кезде балапандардың жынысын клоакасын (нәжістік органы) арқылы анықтау әдісі қолданылады. Ірі дара ұрықты малдарда (ірі қара, жылқы) ұрықтың жынысын ерте айыру үшін эмбрионның трофобласт клеткаларын микрохирургиялық жолмен немесе амниондық сұйық затты (қағанақты) цитогенетикалық талдау жүргізеді. 3. Партеногенез – организмнің ұрықтандырусыз дамуы. Ұрпақтардың бір ғана жынысын алу эмриондар ұрығының аталық (андрогенез) немесе аналық гаметаларынан (гимогенез) дамуына байланысты. Мысалы, Б.П.Астауров жоғары температурамен әсер ету арқылы тұт көбелегінде партеногенез туғызды, мұнда мейоз болған жоқ, ұрпақ 100% ұрғашы болды. Андрогенезде бүкіл ұрпақ тек қана аталық жынысты болды. Ұрықтанбаған тұт көбелегінің жұмыртқасын ыстық температурамен және рентгенмен сәулелендіру арқылы цитоплазманы бүлдірмей ядроларын өлтіреді. Содан кейін бұл жұмыртқаларды ұрықтандырады. Зигота жұмыртқаға енген екі сперманың қосылуынан түзіледі. Партеногенездік (еркексіз) даму табиғи немесе спонтанды жағдайларда құстарда кездеседі. И.В.Кудрявцев, З.А.Ощепкова, А.К.Голубаев және басқалары тауықтардың әр түрлі тұқымдарына эксперимент жүргізу арқылы өмір сүре алатын партеногендердің генетикалық селекциясының мүмкіндігі бар екенін және партеногенезге бейімді минияларды жасауға болатынын көрсетті. Бірақ балапандардың барлығы әтештер болады. Партеногенездің жиілігін жоғарлату үшін жасалған сұрыптауда күрке тауықтардың екі линиясы мынандай нәтиже берді. Жұмыртқалардың партеногенездік қабілеті 1,1-ден 18,6% -ке дейін бір линияда 21,1% дейін өсті. Партеногенездің маңызы тек қана бір жынысты ұрпақ алу үшін ғана емес, бұл құбылысты пайдалана отырып аталық-аналықтың біреуінің ғана генотипіне ұқсас алуға болатындығында. Табиғи партеногенез – сирек кездесетін құбылыс. Төтенше жағдайларда партеногенез көбею әдісі деп саналады. Мысалы: су шаяны (ракообразные), ара. Партеногенез көбеюдің жалғыз әдісі болмайды, ол жыныс көбеюімен кезектестіреді. Қолдан (жасанды) партеногенез – барлық жануарлардың түрлерінде болуы мүмкін. Бірақ олардың жұмыртқаларының даму қабілеті ұрықтанусыз бөлек. Әдебиеттер С.Ж. Стамбеков. Жалпы генетика. А., Ана тілі 1993. Б.К. Бегимкул Генетика А., 2000. Ф.Айала . Кайгер Дж. Современная генетика I - III том М., Мир,1987 М.М.Гершензон, Основы современной генетики. Киев,1983 |