Экология учебник для вузов - А.С. Степановских. Экология
 Скачать 21.33 Mb. Скачать 21.33 Mb.
|

6.3. Фитогенные факторыВ отечественной литературе наиболее распространена классификация форм взаимоотношений между растениями по В. Н. Сукачеву (табл. 6.2). Таблица 6.2 Основные формы взаимоотношений между растениями (по В. Н. Сукачеву, Н. В. Дылису и др., 1964) 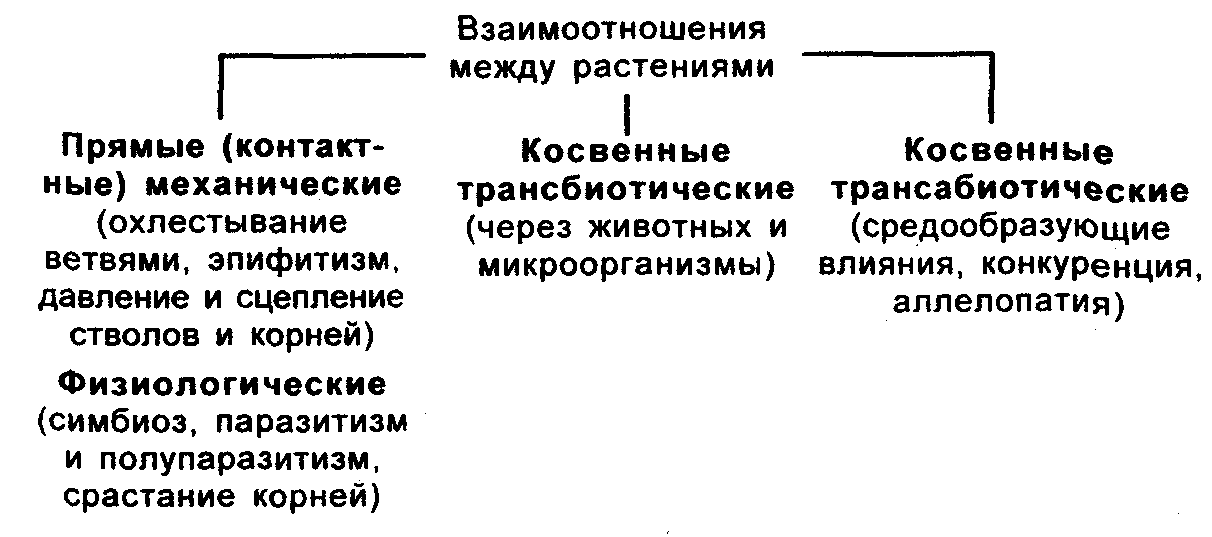 Прямые (контактные) взаимодействия между растениями. Примером механического взаимодействия является повреждение ели и сосны в смешанных лесах от схлестывающего действия березы. Раскачиваясь от ветра, тонкие ветви березы ранят хвоюели, сбивают легкие молодые иглы. Очень заметно это сказывается зимой, когда ветви березы безлиственны. Взаимное давление и сцепление стволов нередко оказывает отрицательное воздействие на растения. Однако чаще такие контакты встречаются в подземной сфере, где большие массы корней тесно переплетаются в небольших объемах почвы. Типы контактов могут быть различны — от простого сцепления до прочного срастания. Так, губительным в жизни многих деревьев тропического леса оказывается разрастание лиан, зачастую приводящее к обламыванию ветвей под их тяжестью и усыханию стволов в результате сдавливающего действия вьющимися стеблями или корнями. Не случайно некоторые лианы называют «душителями» (рис. 6.8). К форме механических контактов относится и использование в качестве субстрата одним растением другого. Растения, живущие на других растениях (на ветвях, стволах деревьев), без связи с почвой, получили название эпифитов, а поселяющиеся на листьях — эпифиллов. В отличие от паразитов они не вступают в прямой физиологический контакт с растением-субстратом, а самостоятельно существуют как автотрофные организмы. 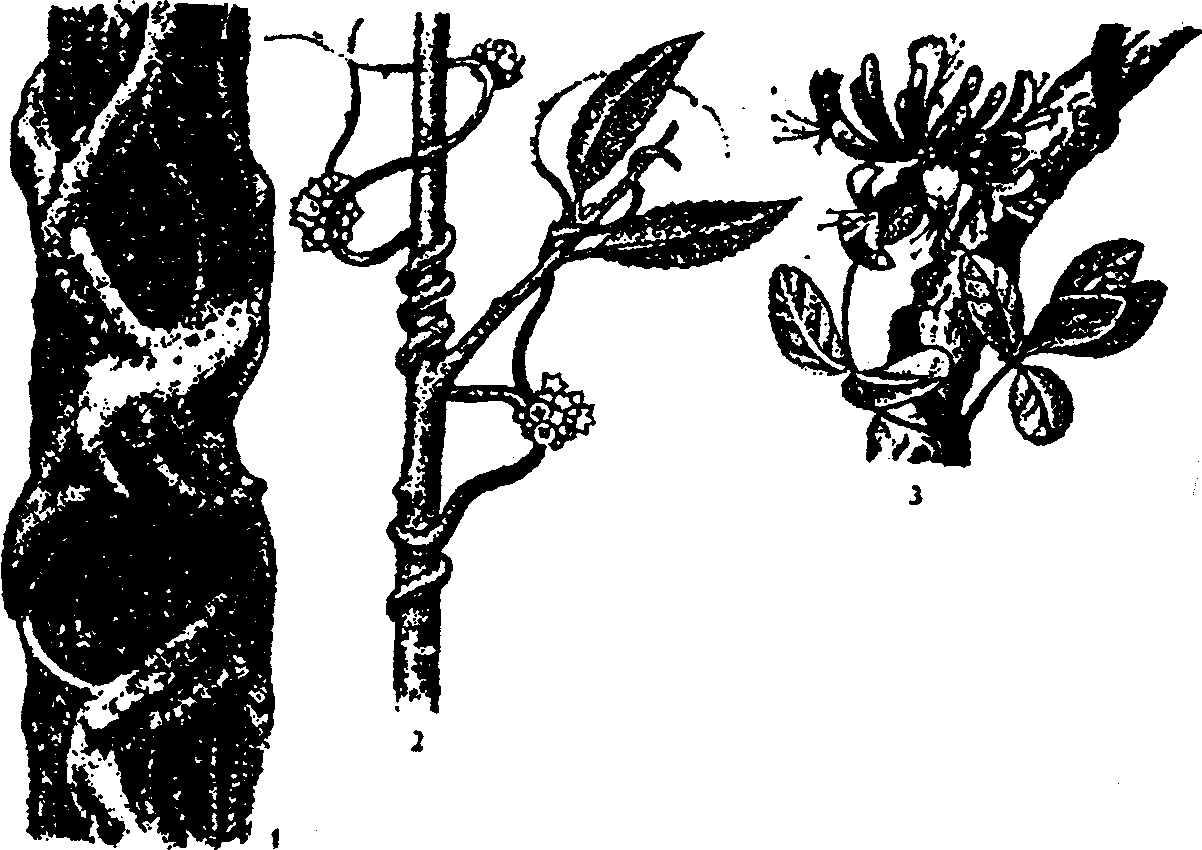 Рис. 6.8. Растения-лианы: 1 — фикус-душитель; 2 — повилика; 3 — жимолость вьющаяся (по Н. М. Черновой и др., 1995) По мнению ученых, около 10% всех видов растений веду эпифитный образ жизни. Наиболее богаты эпифитами тропические леса. К ним относятся многие виды бромелиевых, орхидейных (рис. 6.9). 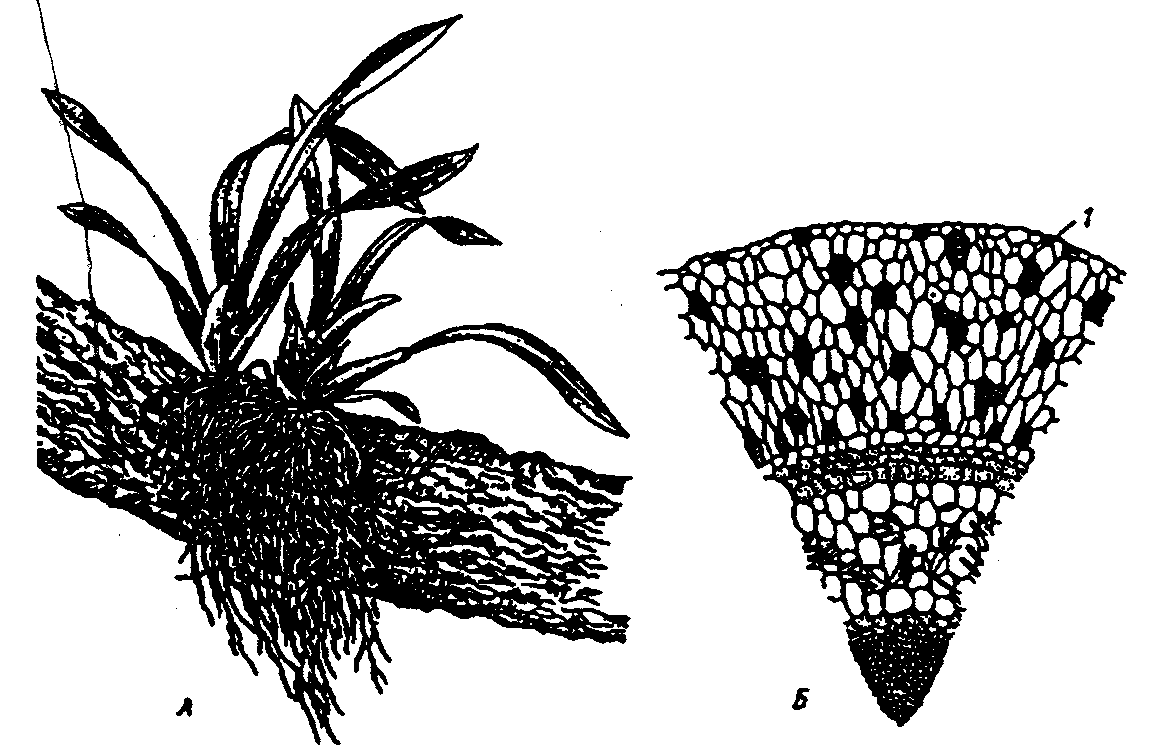 Рис. 6.9. Эпифитная орхидея с воздушными корнями: А — общий вид; Б — поперечный срез воздушного корня с наружным слоем всасывающей ткани (1) (по В. Л. Комарову, 1949) Экологический смысл эпифитизма состоит в своеобразной адаптации к световому режиму в густых тропических лесах: возможность выбраться к свету в верхних ярусах леса без больших затрат веществ на рост. Само происхождение эпифитного образа жизни связывают с борьбой растений за свет. Эволюция многих эпифитов зашла так далеко, что они уже потеряли способность расти вне растительного субстрата, т. е. являются облигатными эпифитами. Вместе с тем есть виды, которые в оранжерейных условиях могут расти и в почве. Физиологические контакты между растениями включают паразитизм, симбиоз, сапрофитизм, срастание корней. Паразитизм — наиболее яркий пример прямых физиологических воздействий между растениями, т.е. переход одного из партнеров на гетеротрофный способ питания и существование за счет организма-хозяина. Например, повилика, питающаяся соками клевера, угнетает его, не только подавляя развитие вегетативной массы, но и заметно задерживая развитие растения. Как правило, урожай семян пораженного клевера невысокий. Через несколько лет на лугу, где появилась повилика, клевер полностью выпадает из травостоя. Паразиты, как уже было отмечено ранее, многочисленны среди грибов и бактерий, значительно меньше распространены среди цветковых растений. Характерным примером тесного симбиоза, или мутуализма между растениями является сожительство водоросли и гриба, которые образуют особый целостный организм-лишайник (рис. 6.10). 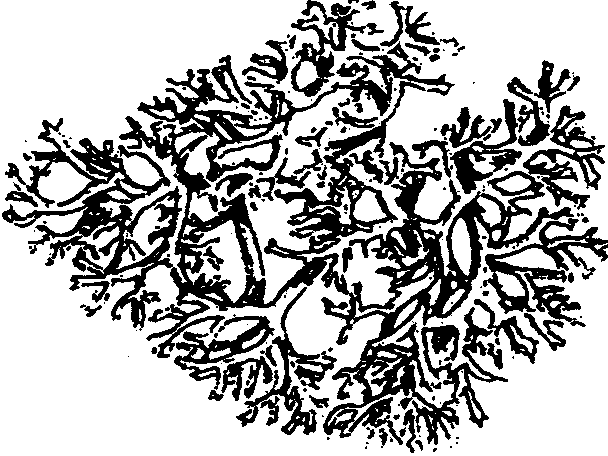 Рис. 6.10. Лишайник-кладония (по Н. М. Черновой и др., 1995) Связи между партнерами-симбионтами довольно сложны, и некоторые авторы называют симбиоз «хорошо урегулированным взаимным паразитизмом». Фотосинтезирующие водоросли в лишайнике снабжают гриб углеводами и другими органическими веществами (нуклеиновые кислоты, протеины, активаторы роста и др.). Грибы поставляют водоросли влагу и минеральные вещества. При недостатке же света и других неблагоприятных условий, угнетающих фотосинтез, водоросли могут получать от грибов и органические вещества, которые они поглощают из субстрата. Лишайники интересны тем, что на их примере можно наблюдать постепенную эволюцию от паразитизма к мутуализму. У более примитивных лишайников гриб фактически проникает в клетки водоросли и по существу является паразитом (рис. 6.11А). У эволюционно более развитых видов водоросль и гриб живут в гармоничных отношениях, принося друг другу взаимную пользу (рис. 6.11Б и В). 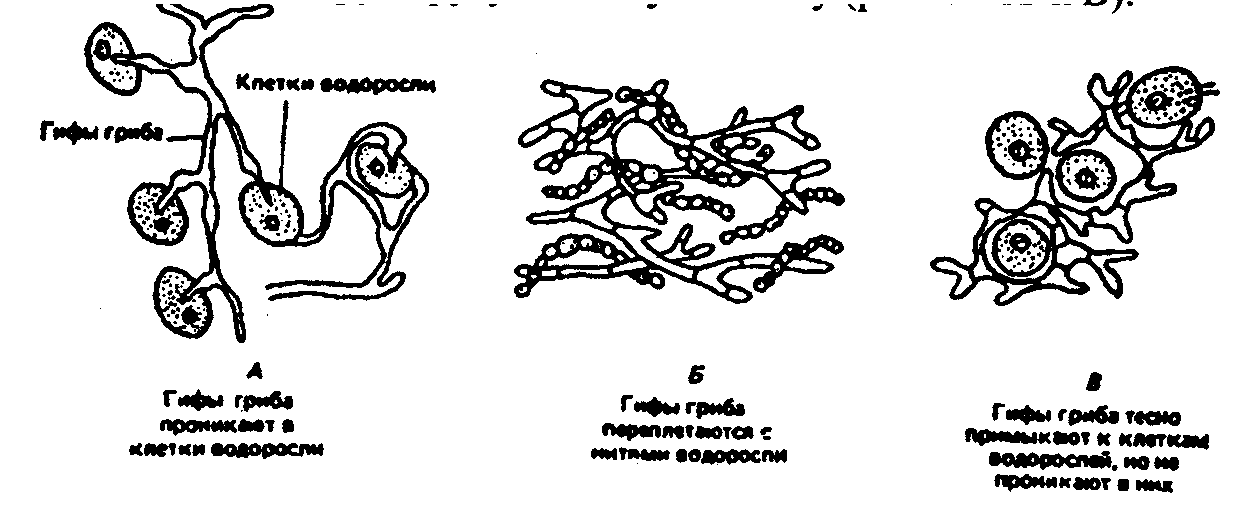 Рис. 6.11. Направление эволюции от паразитизма к мутуализму у лишайников (по Е. Одуму, 1963) Другой пример симбиоза — это сожительство высших растений с бактериями, так называемая бактериотрофия. Симбиоз с клубеньковыми бактериями-азотофиксаторами широко распространен среди бобовых (93% изученных видов) и мимозовых (87%). Так, бактерии из рода Rhizobium, живущие в клубеньках на корнях бобовых растений, обеспечиваются пищей (сахара) и ме-стообиганием, а растения получают от них взамен доступную форму азота (рис. 6.12). 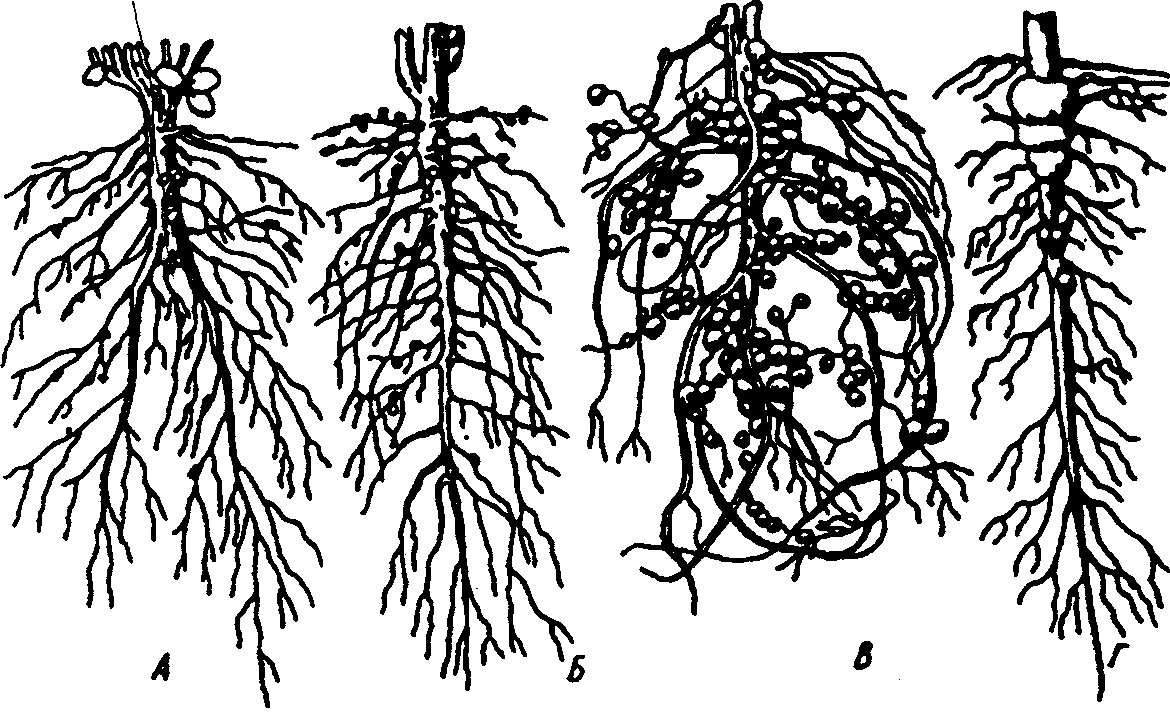 Рис. 6.12. Клубеньки на корнях бобовых растений: А — клевера красного; Б — фасоли; В — сои; Г — люпина (по А. П. Шенникову, 1950) Встречается симбиоз мицелия гриба с корнем высшего растения, или микоризообразование. Такие растения называют микотрофными, или микотрофами. Поселясь на корнях растений, гифы гриба обеспечивают высшее растение колоссальной всасывающей способностью. Поверхность соприкосновения клеток корня и гиф в эктотрофной микоризе в 10—14 раз больше, чем поверхность контакта с почвой клеток «голого» корня, тогда как всасывающая поверхность корня за счет корневых волосков увеличивает поверхность корня лишь в 2—5 раз. Из изученных в нашей стране 3425 видов сосудистых растений микориза обнаружена у 79%. В качестве примера симбиоза грибов с насекомыми можно привести симбиоз грибка Septobasidium с насекомым-червецом из сем. Coccidae, дающее новое симбиотическое образование — лаки, которое как единый организм введено в культуру человеком. Отдельную группу растений с гетеротрофным питанием составляют сапрофиты — виды, которые используют в качестве источника углерода органические вещества отмерших организмов. В биологическом круговороте это важное звено, осуществляющее разложение органических остатков и перевод сложных соединений в более простые, представлено большей частью грибами, актиномицетами, бактериями. Встречаются среди цветковых у представителей семейств грушанковых, орхидных и др. Примерами цветковых, полностью утративших хлорофилл и перешедших на питание готовыми органическими веществами, являются сапрофиты хвойных лесов — подъельник обыкновенный (Monotropa hypopitis), надбородник безлистный (Epipogon aphylluon). Среди мхов и папоротников сапрофиты редки. Срастание корней близко растущих деревьев (одного и того же вида или родственных видов) относится также к прямым физиологическим контактам между растениями. Явление не столь , уж редкое в природе. В густых насаждениях ели Picea flies срастаются корнями около 30% всех деревьев. Установлено, что между сросшимися деревьями существует обмен через корни в виде переноса питательных веществ и воды. В зависимости от степени различия или сходства потребностей сросшихся партнеров между ними не исключены отношения как конкурентного характера в виде перехвата веществ более развитым и сильным деревом, так и симбиотические. Определенное значение имеет форма связей в виде хищничества. Хищничество широко распространено не только между животными, но и между растениями и животными. Так, ряд насекомоядных растений (росянка, непентес) относят к хищникам (рис. 6.13).  Рис. 6.13. Хищное растение росянка (по Е. А. Криксунову и др., 1995) Косвенные трансбиотические взаимоотношения между растениями (через животных и микроорганизмы). Важная экологическая роль животных в жизни растений состоит в участии в процессах опыления, распространения С9мян и плодов. Опыление растений насекомыми, получившее название энтомофилии, способствовало выработке ряда приспособлений как у растений, так и насекомых. Назовем здесь такие интересные адаптации энтомофильных цветков:] узоры, образующие «путевые нити» к нектарникам и тычинкам, н  ередко видимые только в ультрафиолетовых лучах, доступных для зрения насекомых; различие окраски цветков до и после опыления; синхронизация суточных ритмов раскрывания венчика и тычинок, обеспечивающие безошибочное попадание пь1пьцы на тело насекомого, а с него — на рыльце другого цветка и т. д. (рис. 6.14). ередко видимые только в ультрафиолетовых лучах, доступных для зрения насекомых; различие окраски цветков до и после опыления; синхронизация суточных ритмов раскрывания венчика и тычинок, обеспечивающие безошибочное попадание пь1пьцы на тело насекомого, а с него — на рыльце другого цветка и т. д. (рис. 6.14).Рис. 6.14. Насекомое на цветке (по Н. М. Черно- вой и др., 1995) Разнообразное и сложное строение цветков (различная форма лепестков, симметричное или несимметричное их расположение, наличие определенных соцветий), называемое гетеростилией, — все это приспособления к строению тела и поведению строго конкретных насекомых. Например, цветки дикой моркови (Daucus carota), тмина (Carum carvi), опыляемые муравьями, цветки копытня Asarum europaeum, опыляемые муравьями и соответственно не поднимающиеся из-под лесной подстилки. В опылении растений принимают участие и птицы. Опыление растений с помощью птиц, или орнитофилия, находит широкое распространение в тропических и субтропических областях южного полушария. Здесь известно около 2000 видов птиц, которые опыляют цветки при поисках нектара или ловле насекомых, прячущихся в их венчиках. Среди них наиболее известные опылите-ли-нектарницы (Африка, Австралия, Южная Азия) и колибри (Южная Америка). Цветки орнитофильных растений крупные, ярко окрашенные. Преобладает ярко-красная окраска, наиболее привлекательная для колибри и других птиц. В некоторых орнитофильных цветках существуют специальные защитные устройства, которые не дают нектару вылиться при движении цветка. Реже встречается опыление растений млекопитающими, или зоо-гамия. Большей частью зоогамия отмечается в Австралии, в лесах Африки и Южной Америки. Например, австралийские кустарники из рода Driandra опыляются с помощью кенгуру, охотно пьющих их обильный нектар, переходя от цветка к цветку. Распространение семян, плодов, спор растений при помощи животных называют зоохорией. Среди растений, чьи семена, плоды разносятся животными, в свою очередь, различают эпизоо-хорные, эндозоохорные и синзоохорные. Эпизоохорные растения большей частью открытых мест обитания имеют у семян, плодов всевозможные приспособления для закрепления и удерживания на поверхности тела животных (выросты, крючки, прицепки и др.), например лопухи большой и паутинистый, липучка обыкновенная и т. д. В кустарниковом ярусе лесов, где обитает много птиц, преобладают эндозоохорные виды растений. Их плоды съедобны или привлекательны для птиц яркой окраской или сочным околоплодником. Следует отметить, что у семян многих эндозоохорных растений повышается всхожесть, а иногда и способность к прорастанию только после прохождения через пищевой тракт животного — многие аралиевные, яблоня Сиверса (Malus sieversu) и др. Съедобные плоды исемена дуба, сосны сибирской животные не поедают сразу, а растаскивают и складывают в запас. Значительная их часть при этом теряется и дает при благоприятных условиях начало новым растениям. Данное распространение семян и плодов получило название синзоохории. В косвенных трансбиотических взаимоотношениях эдежду растениями нередко выступают микроорганизмы. Ризосфера корней многих деревьев, к примеру, дуба, сильно изменяет почвенную среду, особенно ее состав, кислотность, и тем самым создает благоприятные условия для поселения там различных микроорганизмов, в первую очередь бактерий, таких, как Azotobacter chroocoteum, Tricholome legnorum, Pseudomonas sp. Эти бактерии, поселившись здесь, питаются выделениями корней дуба и органическими остатками, создаваемыми гифами микоризообразующих грибов. Бактерии, живя рядом с корнями дуба, служат своеобразной «оборонительной линией» от проникновения в корни патогенных грибов. Этот биологический барьер создается при помощи антибиотиков, выделяемых бактериями. Поселение бактерий в ризосфере дуба сразу же сказывается положительно на состоянии растений, особенно молодых. Косвенные трансабиотические взаимоотношения между растениями (средообразующие влияния, конкуренция, аллелопатия). Изменение растениями среды — это наиболее универсальный и широко распространенный тип взаимоотношений растений при их совместном существовании. Когда тот или иной вид или группа видов растений в Ц результате своей жизнедеятельности сильно изменяет в количественном и качественном отношении основные экологические факторы таким образом, что другим видам сообщества приходится жить в условиях, которые значительно отличаются от зонального комплекса факто- ls ров физической среды, то это говорит о средообразующей роли, средообразующем влиянии первого вида по отношению к остальным. Один из них — взаимовлияния через изменения факторов микроклимата (например, ослабление солнечной радиации внутри растительного покрова, обеднение ее фотосинтетически активными лучами, изменение сезонного ритма освещенности и др.). Одни растения влияют на другие и через изменение температурного режима воздуха, его влажности, скорости ветра, содержания углекислоты и т. д. Другой путь взаимодействия растений в сообществах — через напочвенный слой мертвых растительных остатков, называемых на лугах и в степях ветошью, травянистым спадом или «степным войлоком», а в лесу — подстилкой. Этот слой (иногда толщиной в несколько сантиметров) вызывает затруднение для проникновения семян и спор в почву. Прорастающие в слое ветоши (или на нем) семена часто гибнут от высыхания раньше, чем корни проростков достигнут почвы. Для семян, попавших в почву и прорастающих, напочвенные остатки могут являться серьезным механическим препятствием на пути ростков к свету. Возможны и взаимоотношения растений через содержащиеся в подстилке продукты распада растительных остатков, тормозящих или, напротив, стимулирующих рост растений. Так, в свежем опаде ели или бука содержатся вещества, тормозящие прорастание ели и сосны, а в местах со скудными осадками и слабым промыванием подстилки могут угнетать естественное возобновление древесных пород. Водные вытяжки из лесных подстилок отрицательно действуют и на рост многих степных трав. Существенный путь взаимного влияния растений — это взаимодействие через химические выделения. Растения выделяют в окружающую среду (воздух, воду, почву) разнообразные химические вещества в процессе гуттации, секреции нектара, эфирных масел, смол и т. д.; при вымывании минеральных солей дождевыми водами листья, например, деревьев, теряют калий, натрий, магний и другие ионы; в ходе метаболизма (корневые выделения) газообразные вещества, выделяемые надземными органами, — непредельные углеводороды, этилен, водород и др.; при нарушении целостности тканей и органов растения выделяют летучие вещества, так называемые фитонциды, и вещества из отмерших частей растений (рис. 6.15). Выделяемые соединения необходимы растениям, но с развитием большой поверхности тела растений их потеря столь же неизбежна, как и транспирация. Химические выделения растений могут служить одним из способов взаимодействия между растениями в сообществе, оказывая на организмы либо токсичное, либо стимулирующее действие. 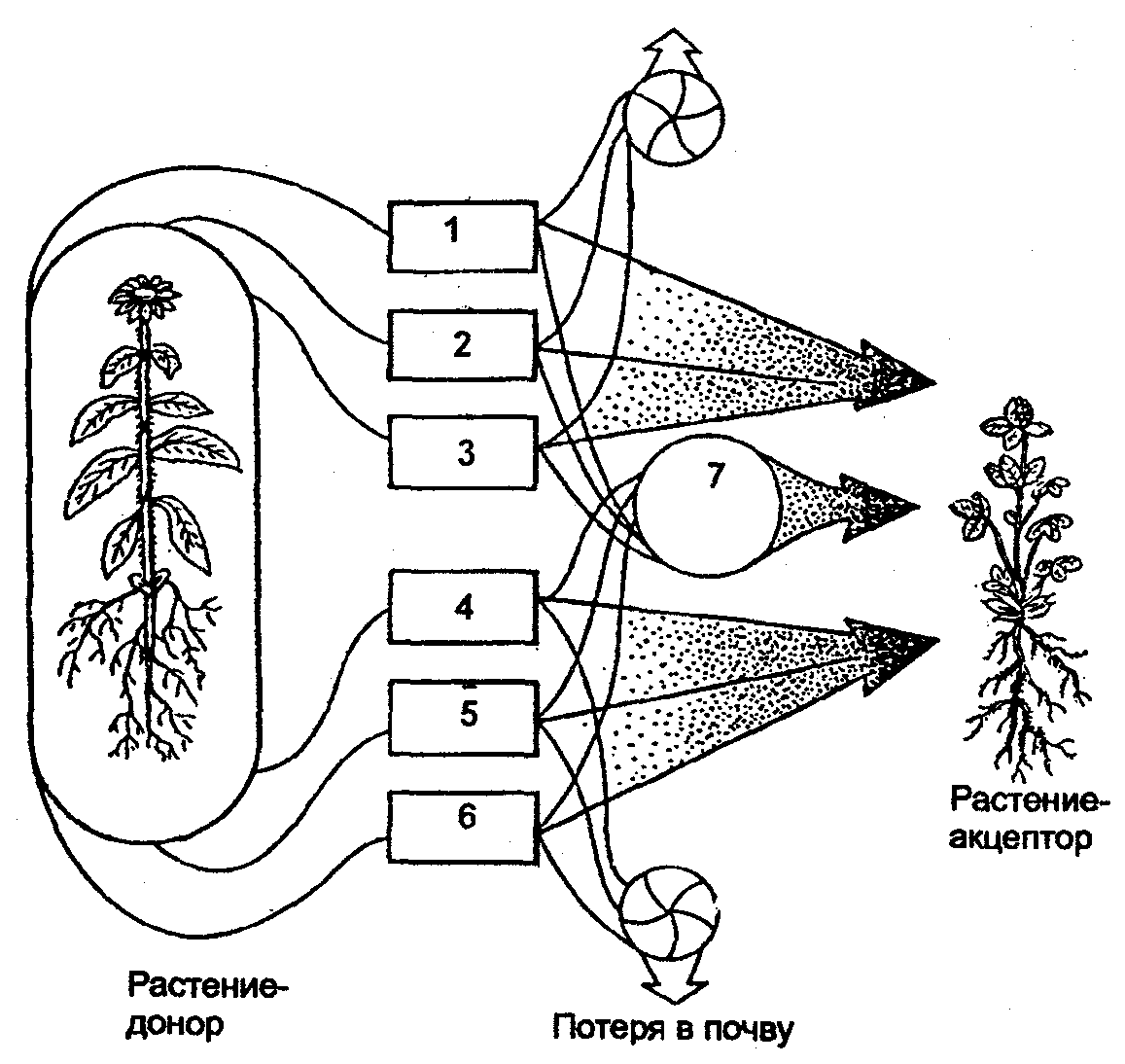 Рис. 6.15. Влияние одного растения на другое (по А. М. Гродзинскому, 1965): 1 — миазмины; 2 — фитонцидные вещества; 3 — фитогенные вещества; 4 — активные прижизненные выделения; 5 — пассивные прижизненные выделения; 6 — посмертные выделения; 7 — переработка гетеротрофными организмами Такие химические взаимовлияния получили название аллелопатии. В качестве примера можно назвать выделения соплодий свеклы, тормозящие прорастание семян куколя (Agrostemma githago). Нут (Cicer arietinum) подавляюще действует на картофель, кукурузу, подсолнечник, томаты и другие культуры, фасоль — на рост яровой пшеницы; корневые выделения пырея (Agropyron repens) и костреца (Bromus inermis) — на растущие вблизи с ними другие травянистые растения и даже деревья. Как крайнюю форму аллелопатии или невозможность существования того или иного вида в присутствии другого в результате интоксикации среды, называют аменсализмом. Аменсализм соответствует прямой конкуренции, антибиозу и антагонизму. Так, благодаря выделению корнями токсических веществ ястребянка (Hieracium pilosella) из семейства сложноцветных вытесняет другие однолетние растения и нередко образует чистые заросли на довольно больших площадях. Многие грибы и бактерии синтезируют антибиотики, которые тормозят рост других бактерий. Аменсализм широко распространен в водной среде. У разных видов растений степень воздействия на среду и таким образом на жизнь обитателей неодинакова в соответствии с особенностями их морфологии, биологии, сезонного развития и др. Растения, наиболее активно и глубоко преобразующие среду и определяющие условия существования для других сооби-тателей, называют эдификаторами. Различают сильные и слабые эдификаторы. К сильным эдификаторам относят ель (сильное затенение, обеднение почв питательными веществами и др.), сфагновые мхи (задержание влаги и создание избыточного увлажнения, увеличение кислотности, особый температурный режим и т. д.). Слабыми эдификаторами являются лиственные породы с ажурной кроной (береза, ясень), растения травянистого покрова лесов. В качестве особой формы трансбиотических взаимоотношений растений выделяют конкуренцию. Это те взаимные или односторонние отрицательные влияния, которые возникают на основе использования энергетических и пищевых ресурсов мес-тообитания. Сильное влияние на жизнь растений оказывает конкуренция за почвенную влагу (особенно четко выражена в областях с недостаточным увлажнением) и конкуренция за питательные вещества почвы, более заметная на бедных почвах. Примером конкуренции могут служить взаимоотношения лисохвоста лугового (Alopecurus pratensis) и типчака (Festuca sulcata). Типчак может произрастать во влажной почве, однако в сообществе лисохвостного луга не растет из-за подавления теневыносливым и быстро растущим лисохвостом. В формировании типчакового или лисохвостного фитоценоза решающее значение имеет не влажность почвы, а конкурентные взаимоотношения типчака и лисохвоста. В более сухих местообитаниях типчак заглушает лисохвост, а на увлажненных лугах выходит победителем лисохвост. Межвидовая конкуренция проявляется у растений так же, как и внутривидовая (морфологические изменения, снижение плодовитости, численности и т. д.). Доминирующий вид постепенно вытесняет или сильно снижает его жизнеспособность. Самая жесткая конкуренция, нередко с непредвиденными последствиями, возникает при введении в сообщества новых видов растений без учета уже сложившихся отношений. |