Факторы препятствующие самоопылению. Факторы препятствующие самоопылению
 Скачать 0.7 Mb. Скачать 0.7 Mb.
|

Факторы препятствующие самоопылению.Для обеспечения перекрестного опыления и устранения самоопыления у растений имеется ряд приспособлений. В случае обоеполости цветков наблюдаются: 1) автостерильность, или самобесплодность — неспособность пыльцы прорастать на рыльце пестика того же самого цветка (встречается у большинства сортов яблони, груши, вишни, некоторых орхидных); 2) дихогамия— неодновременное созревание в обоеполых цветках тычинок и пестиков; она выражена в двух формах: протероандрия и протерогиния; при протероандрии пыльники созревают раньше пестиков (виды семейств бобовых, гвоздичных, гераниевых, астро вых, зонтичных); при протерогинии пестики в цветках созревают раньше тычинок (капустные, розовые, злаки); 3) такое расположение тычинок и пестиков, при котором пыльца не может попасть на рыльце пестика того же самого цветка (вероника, губоцветные); 4)гетеростилия, или разностолбчатость, когда у одних особей в цветках образуются длинные столбики, превышающие длину тычинок, у других в цветках столбики короткие, а тычинки — длинные (примула, медуница) Самоопыление имеет одно существенное преимущество по сравнению с перекрестным: оно не зависит от погодных условий и посредников, поэтому осуществляется при любых условиях. Дихогамия — это функциональная раздельнополость. Она вызвана разновременным созреванием и экспонированием пыльцы и рыльца в цветках, вследствие чего последние выступают то в мужской (пыльниковой), то в женской (рыльцевой) фазе, Дихогамия проявляется в форме протандрии (при более раннем созревании пыльцы) или протогинии (при более раннем созревании рылец). Одновременное созревание пыльцы и рылец в цветке носит название гомогамии. File:Zr51tb 001.jpg Таблица 1. Приспособления цветков к опылению: 1 - протандрия у кипрея узколистного, или иван-чая (Chamaenerium angustifolium); 2 - пыльценосный цветок у зверобоя продырявленного (Hypericum perforatum); 3 - взрывчатое цветение у костреца безостого (Bromopsis inermis); 4 - гравитационная автогамия у одноцветки крупноцветковой (Moneses uniflora); 5 - съедобные волоски на тычинках у коровяка медвежьего уха (Verbascum tapsus); 6 - соцветие с наружными бесполыми цветками у калины обыкновенной (Viburnum opulus). 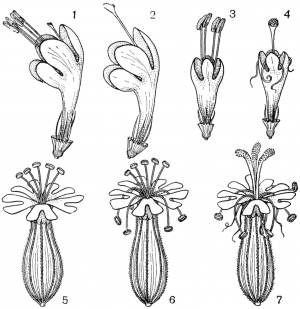 Рис. 31. Примеры протандрии. Скабиоза бледно-желтая (Scabiosa ochroleuca): 1 - краевой цветок в тычиночной стадии; 2 - он же в рыльцевой стадии; 3 - срединный цветок в тычиночной стадии; 4 - он же в рыльцевой стадии. Смолевка вильчатая (Silene dichotoma): 5 - цветок в тычиночной стадии первого дня цветения, 6 - он же в тычиночной стадии второго дня цветения, 7 - он же в рыльцевой стадии третьего дня цветения (все тычинки увяли). Дихогамия обычно рассматривается как приспособление к перекрестному опылению и как средство предотвращения самоопыления. Это традиционное представление о дихогамии нуждается в уточнении. Дихогамия необязательно исключает самоопыление. Нередко в конце цветения возможна автогамия, если по каким-либо причинам перекрестное опыление не осуществилось. Кроме того, самоопыление в форме гейтоногамии возможно у большинства дихогамных растений еще и потому, что на них имеются многочисленные цветки, находящиеся в разных фазах развития. Поэтому значение дихогамии следует усматривать в том, что она более или менее ограничивает самоопыление, способствуя тем самым перекрестному опылению. Такое сочетание перекрестного опыления и самоопыления играет, как уже говорилось, положительную роль в эволюции. Протандрия встречается чаще, чем протогиния. Она более соответствует нормальной последовательности в развитии частей цветка. Протандрия хорошо выражена у зонтичных, сложноцветных, колокольчиковых, гвоздичных, ворсянковых, губоцветных и многих других семейств (рис. 31, табл. 1). У колокольчиков пыльники вскрываются еще в бутоне. Пыльца муфточкой окружает столбик, удерживаясь на волосках, покрывающих его. Опорожненные и подсохшие пыльники видны на дне венчика. Лопасти рыльца в этот момент еще вполне сомкнуты. Примерно сутки спустя после распускания цветка расходятся лопасти рыльца и становится возможным опыление их чужой пыльцой, принесенной насекомыми. Но под конец цветения возможна и автогамия, благодаря тому что лопасти рыльца, закручиваясь спирально вниз, касаются воспринимающей поверхностью столбика, сохранившего собственную пыльцу. 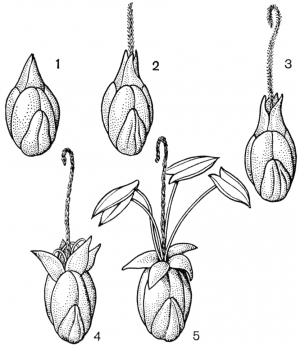 Рис. 32. Протогиния у подорожника Корнута (Plantago cornutii): 1 - цветок в стадии бутона; 2 - появление рыльца; 3 - увядание рыльца; 4 - раскрывание цветка и начало выдвижения тычинок при уже увядшем рыльце; 5 - тычиночная стадия цветка. У смолевок (Silene multiflora, S. chlorantha и др.) автогамия исключена. Они цветут и опыляются ночью. Цветки распускаются в 18- 19 ч, а утром закрываются. В первый вечер при распускании цветка экспонируются пять тычинок, во второй вечер — следующие пять тычинок, увядающих к утру, наконец, ночью на третьи сутки выдвигаются рыльца. Таким образом, пыльниковая и рыльцевая фазы в цветках смолевок разделены, но гейтоногамия возможна, поскольку на особях могут встретиться цветки в разных фазах развития. У зонтичных и ворсянковых протандрия является весьма действенным средством предотвращения самоопыления. У зонтичных протандрия строга, безупречна и охватывает не только отдельные сложные зонтики, но и всю особь в целом. У большинства видов зонтичных это достигается благодаря строгой очередности в цветении зонтиков разного порядка и полной согласованности (синхронности) его у зонтиков данного порядка. Вследствие этого каждая особь последовательно несколько раз выступает то в пыльниковой, то в рыльцевой фазе, причем эти фазы резко разграничены и, как правило, не налегают друг на друга. В качестве примера может служить порезник средний (Libanotis intermedia). Этот тип протандрии (тип Libanotis) с многократной сменой тычиночной и рыльцевой фаз очень обычен у зонтичных. Значительно реже у них встречается другой тип протандрии (тип Peucedanum), характеризующийся однократной сменой фаз у всех одновременно и согласованно цветущих зонтиков особи, независимо от того, к какому порядку они принадлежат. Такая протандрия известна пока у горичника Любименко (Peucedanum lubimenkoanum). Резкая протандрия, всего соцветия, последовательная и одновременная смена фаз в цветении соцветий разных порядков свойственна и некоторым ворсянковым: короставнику полевому (Knautia arvensis), скабиозе бледно-желтой (Scabiosa ochroleuca) и др. У зонтичных и ворсянковых в результате этого невозможна автогамия и гейтоногамия, неизбежным является перекрестное опыление. 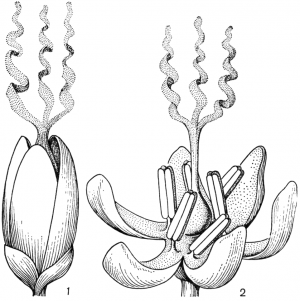 Рис. 33. Протогиния у ситника Жерарда (Juncus gerardii): 1 - цветок в рыльцевой фазе вечером (протогиния); 2 - раскрывшийся цветок утром в момент опыления. Протогиния хорошо выражена у крестоцветных, розовых, барбарисовых, жимолостных и т. д. Во многих случаях разница во времени созревания рыльца и пыльцы столь незначительна, что наличие протогинии кажется сомнительным. Более резко выражена протогиния у ветроопыляемых растений, причем не только у обоеполых, но и у однодомных и двудомных растений. Так обстоит дело у ситниковых, осоковых, злаковых, полыней, подорожников (рис. 32). У ситников популяция вида вступает в рыльцевую фазу с вечера предшествующего дня, а на следующее утро переходит в тычиночную фазу, причем рассеивание пыльцы ограничено 2-3 ч (рис. 33). У однодомных осок рыльцевая фаза предваряет пыльниковую на 1-6, а у подорожников на 4-6 суток. В подобных случаях роль протогинии заключается в том, что заблаговременная экспозиция рылец представляет подготовку к очень быстротечному опылению ветром во время кратких суточных периодов рассеивания пыльцы. Ту же функцию выполняет протогиния у некоторых пустынных маревых. Наиболее действенным средством предотвращения самоопыления, безусловно, является самонесовместимость. Она выражается в отсутствии прорастания пыльцы на рыльце или в прекращении роста пыльцевых трубок в столбике при самоопылении. Самонесовместимость у покрытосеменных распространена более широко, нежели двудомность. По сравнению с последней она обеспечивает более высокую семенную продуктивность, поскольку в этом случае каждый цветок может дать семена, а у двудомных растений — только половина цветков. Кроме того, самонесовместимость обеспечивает максимальную экономию женских гамет, так как возможность появления неудачных комбинаций гамет предупреждается до оплодотворения, Самонесовместимость регулируется генетическими механизмами. Различают гомоморфную и гетероморфную самонесовместимость. Гомоморфная самонесовместимость наиболее распространена в природе: она зарегистрирована примерно у 10 000 видов покрытосеменных из 78 семейств. Самонесовместимость в этом случае не сопровождается морфологическими различиями в строении цветка. Если самонесовместимость сочетается с разностолбчатостью (гетеростилией), то тогда ее называют гетероморфной. 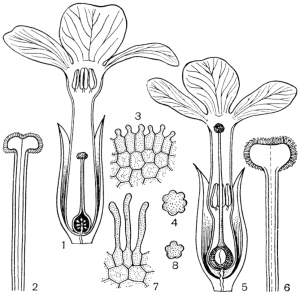 Рис. 34. Гетеростилия у примулы (Primula sp.): 1-4 - короткостолбчатая форма (1 - вид цветка в разрезе; 2 - короткий столбик с сосочковидной поверхностью рыльца при малом увеличении; 3 - то же при большом увеличении; 4 - пыльцевое зерно); 5-8 - длинностолбчатая форма (5 - вид цветка в разрезе; 6 - длинный столбик с сосочковидной поверхностью рыльца при малом увеличении; 7 - то же при большом увеличении; 8 - пыльцевое зерно). Явление гетеростилии состоит в том, что у вида имеются две или три формы цветков, находящиеся на разных особях и различающиеся по длине столбиков и расположению тычинок (диморфные и триморфные растения). У диморфных первоцветов (Primula) длинностолбчатая форма имеет цветки с длинным столбиком, рыльце которого находится в зеве венчика, а тычинки — ниже, в глубине венчика. У короткостолбчатой формы обратное расположение столбиков и пыльников (рис. 34). Формы гетеростильных растений различаются и по другим признакам. Так, короткостолбчатые цветки характеризуются по сравнению с длинностолбчатыми более крупной пыльцой и более мелкими сосочками рыльца. Самоопыление у той и другой формы, а также переопыление между особями одного морфологического типа дает ничтожное количество семян (самонесовместимость), тогда как переопыление между растениями с разной длиной столбика высокоэффективно. Примером диморфных (дистильных) видов могут служить медуница, первоцвет, гречиха, кермек и др. У триморфного гетеростильного дербенника иволистного (Lythrum salicaria) имеются коротко-, средне- и длинностолбчатые цветки, распределенные по разным особям. Тычинки в цветке каждой формы соответствуют по длине столбикам цветков двух других форм. Оплодотворение оказывается наиболее эффективным при опылении рыльца каждой формы цветка пыльцой тычинок соответственной длины из двух других форм цветка, как это было показано еще Ч. Дарвином (рис. 35). Гетероморфная самонесовместимость встречается редко. Она известна у 56 родов из 23 семейств. Особенно она распространена у мареновых (Rubiaceae), среди которых насчитывается 155 гетеростильных видов. 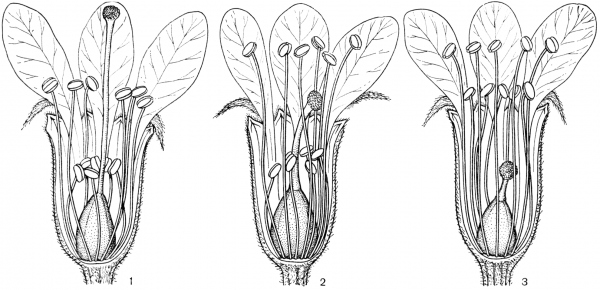 Рис. 35. Гетеростилия у дербенника иволистного (Lythrum salicaria): 1 - длинностолбчатая форма; 2 - среднестолбчатая форма; 3 - короткостолбчатая форма. Самоопыление растенийПеренос пыльцы с тычинок на рыльце того цветка называется самоопылением. У самоопылителей и тычинки, и пестики на одном цветке созревают одновременно. У растений в ходе длительного эволюционного развития выработались приспособления для перекрестного опыления, т.к. оно является прогрессивным. Образуется потомство с признаками материнского и отцовского организмов, а при самоопылении у потомства проявляются признаки лишь одного организма. Самоопыление ведет к снижению урожайности, к вырождению сорта. Поэтому наиболее распространено перекрестное опыление и цветки большинства растений имеют приспособления, препятствующие самоопылению: 1. однополость (орешник, конопля, осина и др.); 2. неодновременное созревание пыльников и рылец; 3. различная длина тычинок и пестиков. Однако самоопыление имеет значение в селекции при выведении чистых линий и может быть запасным актом на тот случай, если не произойдет перекрестного опыления. Самоопыление в естественных условиях встречается реже, чем перекрестное опыление. В естественных условиях не все цветки могут опылиться. Часто пыльца теряет свои свойства из-за воздействия слишком высоких температур или повышенной влажности. В безветренную погоду или, наоборот, при шквальном ветре опыление ветроопыляемых растений трудновыполнимо. Из культурных растений размножаются путем самоопыления, например, пшеница, вика, соя, горох, ячмень и др. Такие растения как гречиха, клевер красный, эспарцет, яблони большинства сортов, нуждаются только в перекрестном опылении. При этом следует отметить, что перекрестное опыление способствует получению более мощных, плодовитых и жизнеспособных растений. Есть группа растений, которые сохранили способность к самоопылению (подсолнечник, люцерна, хлопчатник, малина, крыжовник и некоторые другие), но и они при перекрестном опылении насекомыми дают более высокие урожаи. У многих растений в процессе длительного эволюционного развития выработался целый ряд приспособлений, препятствующих самоопылению и способствующих перекрестному опылению. У целой группы растений невозможно самоопыление, потому что у них женские цветки с пестиками развиваются на одном экземпляре, а мужские с тычинками на другом (однополые). Такие растения принято называть двудомными. В эту группу входят ива, тополь, конопля, клубника и др. Есть растения, у которых цветки, так же как и у двудомных растений, имеют или тычинки или пестики, но они находятся на одном и том же экземпляре. Такие растения называют однодомными. Препятствием самоопылению является неодновременное созревание тычинок и пестиков, находящихся в одном и том же цветке (обоеполый цветок). Так, у подсолнечника, крыжовника, кипрея пыльники созревают раньше, чем рыльце, а у яблони, груши, подорожника и других раньше созревает рыльце. В этих случаях опыление происходит пыльцой с других цветков, перенесенной насекомыми. У ряда растений (гречиха, медуница, дербенник) самоопыление затруднено из-за того, что цветки имеют разностолбчатые органы: у одних длинные тычинки и короткие пестики, у других наоборот — длинные пестики и короткие тычинки. Поэтому в пределах цветка опыление собственной пыльцой, как правило, не осуществляется. У некоторых растений (клевер красный, эспарцет) наблюдаются явления самобесплодности (самостерильности). В этом случае собственная пыльца на рыльце цветка не прорастает или прорастает значительно медленнее, чем попавшая с другого цветка. Самобесплодность встречается у яблони, груши, черешни и ряда других плодовых и ягодных культур, оплодотворение у которых происходит лишь в том случае, если на рыльце пестиков одного сорта попала пыльца с цветков другого сорта. Чтобы создать условия для нормального плодоношения этих культур, необходимо в садах иметь соответствующие сорта плодовых деревьев, обеспечивающие межсортовое опыление. У множества цветковых растений выработан механизм невозможности самоопыления, что предотвращает родственное скрещивание и увеличивают генетическое разнообразие вида. Это достигается благодаря процессу, названному самонесовместимостью, при котором опыление не в состоянии произойти, если пыльца идентифицирует попадание на пестик цветка, на растении которого она появилась. Но принцип действия этого механизма долго оставался загадкой, которая теперь успешно решена. Группа исследователей во главе с Те-Хуи Као, профессора биохимии и молекулярной биологии в Пенсильванском Университете, сообщает в статье, опубликованной 20 мая в выпуске журнала Nature, об открытии у петуний гена, который контролирует этот процесс самонесовместимости пыльцы. Недавно идентифицированный ген получил название PiSLF (от Petunia inflata S-locus F-box). Этот ген кодирует производство большого семейства белков F-box, которые, как известно, вызывают сворачивание белка в различных живых организмах, включая животных, растения и дрожжи. В то время как разновидность петунии может нести от 50 до 60 различных S-аллелей, каждое растение имеет только две из них, по одной от каждого родителя. Аллель - один из множества возможных вариантов специфического гена; например, две аллели существуют для каждого из трех генов, которые определяют цвет глаза у людей. Зерно пыльцы является гаплоидным. Это означает, что оно содержит только половинный набор хромосом, и таким образом каждое зерно пыльцы содержит только одну из двух S-аллелей родительского растения. Пестик, с другой стороны, является диплоидным, и имеет два набора хромосом (по одному от каждого родителя) и поэтому имеет обе S-аллели родительского растения. Во время опыления, если S-аллель пыльцы не соответствует любой из этих двух S-аллелей в пестике, то пыльца прорастет через поверхность рыльца пестика. Из ее вегетативной клетки развивается пыльцевая трубка, которая несет к зародышевому мешку спермии. Достигнув его, спермии сливаются с яйцеклеткой, и происходит оплодотворение. Однако, если S-аллель пыльцы соответствует любой из этих двух S-аллелей в пестике, рост пыльцевой трубки останавливается на одну треть пути к мешку, что предотвращает оплодотворение. Вызов этого ответа самонесовместимости требует взаимодействия между генами S-аллели пыльцы и пестика. Ответственным за эту несовместимость оказался ген PiSLF. Таким образом, открываются большие возможности генной инженерии в плане остановки безудержного скрещивания трансгенных растений с дикими. Ученым уже удалось переделать трансгенные растения в самоопыляемые (то есть те становятся самосовместимыми), и все потомство трансгенных растений также наследует эту совместимость. Это открытие может также получить коммерческое применение для гибридного производства злаковых культур. Развитие гибридных сортов является главной целью садоводческой и сельскохозяйственной практики, потому что гибридные растения наиболее производительные, чем сортовые культуры, полученные из самоопыления или случайного опыления. Однако, чтобы получить гибридный сорт, необходимо перекрестное опыление с двух определенных сортов, при котором самоопыление и родственное опыление (опыление растением того же самого сорта) необходимо исключить. До сих пор для этого применяли ручной метод опыления. Однако этот процесс является очень трудоемким и дорогим. Из-за этого становятся очень дорогими и семена гибридных сортов, а, следовательно, и будущая цена урожая и продуктов питания на нашем столе. Если удастся сократить затраты за счет снижения ручного труда при производстве гибридов, это значительно снизит цены на продукты питания. Меню
Реклама |