лекция по физиологии. Лекция - Физиология нервных проводников. Физиология нервной сист. Физиология нервных проводников
 Скачать 300.42 Kb. Скачать 300.42 Kb.
|

|
Лекция 4. ФИЗИОЛОГИЯ НЕРВНЫХ ПРОВОДНИКОВ Нервное волокно ( нервный проводник) представляет собой отросток нейрона, заключенный в глиальную оболочку. Нервные волокна образуют нервные пучки, совокупность которых формирует нервный ствол, или нерв. Морфологическая организация нервных проводников Различают два вида нервных волокон: • Немиелинизированные (безмякотные). • Миелинизированные (мякотные) Немиелинизированные нервные волокна образуются путем погружения аксона (осевого цилиндра) в цитоплазму леммоцитов (олигодендроцитов в ЦНС и Шванновских клеток на периферии), располагающихся в виде тяжей. Миелинизированные волокна. Осевой цилиндр также окружен глиальными клетками, но эти клетки, многократно вращаясь вокруг осевого цилиндра, образуют пластинки миелиновой оболочки, на 80% состоящей из липидов и 20% из белка. Вокруг миелиновой оболочки располагается тонкий слой цитоплазмы, ядро леммоцита, или шванновской клетки. Снаружи волокно также покрыто базальной мембраной Механизмы проведения нервного импульса в нервных волокнах Нервные волокна как возбудимые системы обладают следующими общими свойствами: возбудимостью, лабильностью, проводимостью. Различают распространение возбуждения по нервному волокну: - непрерывное (характерно для немилинизированных волокон); - сальтаторное ( характерно для миелинизированных волокон). Непрерывное распространение нервного импульса. В состоянии покоя вся внутренняя поверхность мембраны нервного волокна несет отрицательный заряд, а наружная сторона мембраны – положительный. Электрический ток между внутренней и наружной стороной мембраны не протекает, т.к. липидная мембрана имеет высокое электрическое сопротивление. Во время развития потенциала действия, т.е. при открытии натриевых каналов, в возбужденном участке мембраны происходит реверсия заряда. На границе возбужденного и невозбужденного участка начинает протекать электрический ток (движение ионов Nа+).  Внутри нервного волокна возникает ток от положительного полюса к отрицательному полюсу, т. е. ток направлен от возбужденного участка к невозбужденному Электрический ток раздражает ближайший участок мембраны, деполяризует его и приводит его в состояние возбуждения, в то время как ранее возбужденные участки возвращаются в состояние покоя через стадию рефрактерности. В связи с наличием рефрактерности, обратный ход волны оказывается невозможным. Таким образом, волна возбуждения электротонически (пассивно) охватывает все новые участки мембраны нервного волокна. Таким образом, при непрерывном проведении поверхность обладает электрогенными свойствами на всем протяжении. Поэтому малые круговые токи возникают на расстоянии в несколько микрометров. Возбуждение имеет вид постоянно бегущей волны. Размер участка мембраны, находящегося под влиянием ПД, зависит от его длительности и скорости проведения. Например, если длительность ПД равна 2 мс, а скорость его проведения 10 м/с, то потенциал распространится на участке мембраны 2 см. Сальтаторное распространение нервного импульса. В миелинизированном нервном волокне участки мембраны, покрытые миелиновой оболочкой, являются невозбудимыми; возбуждение может возникать только в участках мембраны, расположенных в области перехватов Ранвье, где находится максимальное количество управляемых потенциалчувствительных натриевых каналов - 12 000 на 1 мкм. Время проведения возбуждения по миелиновому волокну обратно пропорционально длине между перехватами. При развитии ПД в одном из перехватов Ранвье происходит реверсия заряда мембраны. Между электроотрицательными и электроположительными участками мембраны возникает электрический ток, который раздражает соседние участки мембраны. Однако в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье. Таким образом, возбуждение распространяется по мембране скачкообразно ( сальтаторно) от одного перехвата Ранвье к другому. "Перепрыгивание" потенциала действия через участок между перехватами возможно потому, что амплитуда ПД в 5-6 раз превышает пороговую величину, необходимую для возбуждения соседнего перехвата. ПД может "перепрыгивать" не только через один, но и через два межперехватных промежутка. Это явление может наблюдаться при снижении возбудимости соседнего перехвата под действием какого-либо фармакологического вещества, например, новокаина, кокаина и др. Сальтаторное проведение возбуждения является высокоскоростным, высокоэкономичным. Нервные волокна по скорости проведения возбуждения, диаметру волокна, длительности различных фаз потенциала действия и строению принято подразделять на три типа: А, В, С. Волокна типа А в свою очередь делятся на подтипы: А-альфа, А-бета, А-гамма, А-дельта. 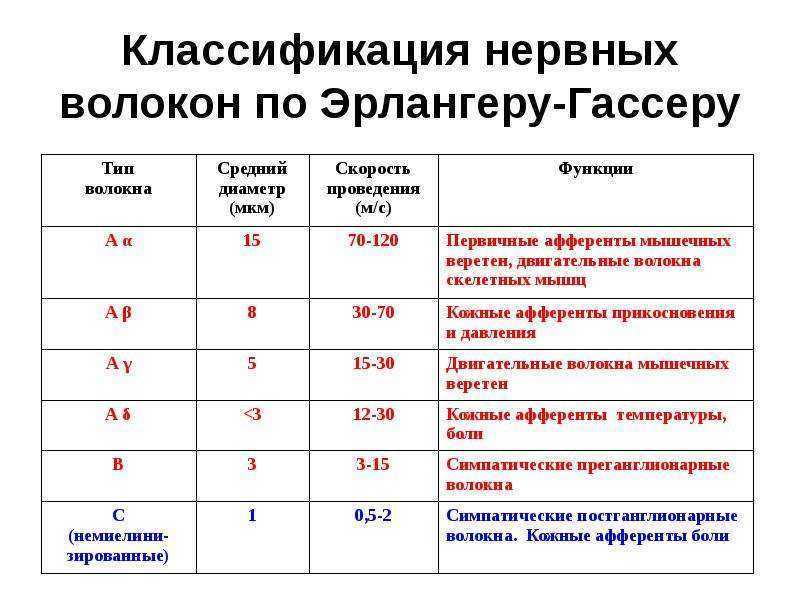 Проведение возбуждения по нервному волокну не нарушается в течение длительного ( многочасового) времени. Это свидетельствует о малой утомляемости нервного волокна. Считают, что нервное волокно относительно неутомляемо вследствие того, что процессы ресинтеза энергии в нем идут с достаточно большой скоростью и успевают восстановить траты энергии, происходящие при прохождении возбуждения. В момент возбуждения энергия нервного волокна тратится на работу натрий-калиевого насоса. Особенно большие траты энергии происходят в перехватах Ранвье вследствие большой плотности здесь натрий-калиевых каналов. Распространение потенциала действия в дендрите представляет собой гораздо более сложный процесс, чем в аксоне. Во-первых, в аксоне справедливо допущение, что подпороговые изменения потенциала не влияют на пассивные свойства мембраны. В дендритах, напротив, это допущение невозможно благодаря наличию целого ряда потенциалзависимых проводимостей, кроме тех, что обычно участвуют в генерации потенциала действия. Положение более усложняется тем, что в дендритном дереве ПД соседствуют с синаптическими потенциалами (ВПСП и ТПСП). Законы проведения возбуждения в нервных волокнах 1. Закон двустороннего проведения - возбуждение, возникающее в одном участке нерва, распространяется в обе стороны от места своего возникновения. Это можно доказать, если на нервное волокно наложить регистрирующие электроды на некотором расстоянии друг от друга, а между ними нанести раздражение. Возбуждение зафиксируют электроды по обе стороны от места раздражения. В организме возбуждение всегда распространяется по аксону от тела клетки (ортодромно). 2. Закон анатомической и физиологической целостности - возбуждение может распространяться по нервному волокну только в случае его морфологической и функциональной целостности. Различные факторы, воздействующие на нервное волокно (наркотические вещества, охлаждение, перевязка и т. д.) приводят к нарушению физиологической целостности, т. е. к нарушению механизмов передачи возбуждения. Несмотря на сохранение его анатомической целостности, проведение возбуждения в таких условиях нарушается. Н. Е. Введенский обнаружил, что если участок нерва подвергнуть альтерации (т. е. воздействию повреждающего агента) посредством, например, отравления или повреждения, то лабильность такого участка резко снижается. Восстановление исходного состояния нервного волокна после каждого потенциала действия в поврежденном участке происходит медленно. При действии на этот участок частых раздражителей он не в состоянии воспроизвести заданный ритм раздражения, и поэтому проведение импульсов блокируется. Такое состояние пониженной лабильности было названо Н. Е. Введенским парабиозом. В развитии состояния парабиоза можно отметить три, последовательно сменяющих друг друга, фазы: уравнительную, парадоксальную, тормозную. В уравнительную фазу происходит уравнивание величины ответной реакции на частые и редкие раздражители. В нормальных условиях функционирования нервного волокна величина ответной реакции иннервируемых им мышечных волокон подчиняется закону силы: на редкие раздражители ответная реакция меньше, а на частые раздражители - больше. При действии парабиотического агента и при редком ритме раздражении (например, 25 Гц) все импульсы возбуждения проводятся через парабиотический участок, так как возбудимость после предыдущего импульса успевает восстановиться. При высоком ритме раздражении (100 Гц) последующие импульсы могут поступать в тот момент, когда нервное волокно еще находится в состоянии относительной рефрактерности, вызванной предыдущим потенциалом действия. Поэтому часть импульсов не проводится. Если проводится только каждое четвертое возбуждение (т. е. 25 импульсов из 100) , то амплитуда ответной реакции становится такой же, как на редкие раздражители (25 Гц) - происходит уравнивание ответной реакции. В парадоксальную-фазу происходит дальнейшее снижение лабильности. При этом на редкие и частые раздражители ответная реакция возникает, но на частые раздражители она значительно меньше, т. к. частые раздражители еще больше снижают лабильность, удлиняя фазу абсолютной рефрактерности. Следовательно, наблюдается парадокс - на редкие раздражители ответная реакция больше, чем на частые. В тормозную фазу лабильность снижается до такой степени, что и редкие, и частые раздражители не вызывают ответной реакции. При этом мембрана нервного волокна деполяризована и не переходит в стадию реполяризации, т. е. не восстанавливается ее исходное состояние. Явление парабиоза лежит в основе медикаментозного локального обезболивания. Влияние анестезирующих веществ также связано с понижением лабильности и нарушением механизма проведения возбуждения по нервным волокнам. Парабиоз - явление обратимое. Если парабиотическое вещество действует недолго, то после прекращения его действия нерв выходит из состояния парабиоза через те же фазы, но в обратной последовательности. Механизм развития парабиотического состояния сводится к следующему. При воздействии на нервное волокно парабиотического фактора нарушается способность мембраны увеличивать натриевую проницаемость в ответ на раздражение. В участке альтерации инактивация натриевых каналов, вызванная повреждающим агентом, суммируется с инактивацией, вызываемой нервным импульсом, и возбудимость снижается настолько, что проведение следующего импульса блокируется. 3. Закон изолированного проведения - возбуждение, распространяющееся по волокну, входящему в состав нерва, не передается на соседние нервные волокна. Способность нервного волокна к изолированному проведению возбуждения обусловлена наличием оболочек, а также тем, что сопротивление жидкости, заполняющей межволоконные пространства, значительно ниже, чем сопротивления мембраны волокна. Поэтому ток, выйдя из возбужденного волокна, шунтируется в жидкости и оказывается слабым для возбуждения соседних волокон. Основная часть тока, возникающего между возбужденным и невозбужденным участками нервного волокна, проходит по межклеточным щелям, не действуя на рядом расположенные нервные волокна. Изолированное проведение возбуждения имеет важное значение. Нерв содержит большое количество нервных волокон ( чувствительных, двигательных, вегетативных), которые иннервируют различные по структуре и функциям эффекторы (клетки, ткани, органы). Если бы возбуждение внутри нерва распространялось с одного нервного волокна на другое, то нормальное функционирование органов было бы невозможно 4. Закон бездекрементного проведения - амплитуда потенциала действия не изменяется с увеличением расстояния от места его возникновения. ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ. Значение нервной системы – это быстрая и точная передача информации и её интеграция, н.с. обеспечивает взаимосвязь между органами и системами органов, функционирование организма как единого целого, его взаимодействие с внешней средой. Она регулирует и координирует деятельность различных органов, приспосабливает деятельность всего организма как целостной системы к изменяющимся условиями внешней и внутренней среды. С помощью нервной системы осуществляется приём и анализ разнообразных сигналов из окружающей среды и с внутренних органов, формируются ответные реакции на эти сигналы. С деятельностью высших отделов н.с. связано осуществление психических функций – осознание сигналов окружающего мира, их запоминание, принятие решения и организация целенаправленного поведения, абстрактное мышление и речь. Все эти сложные функции осуществляются огромным количеством нервных клеток – нейронов, объединённых в сложнейшие нейронные цепи и центры. Нейроны и глиальные клетки – структурные элементы ЦНС. Нейрон - структурная и функциональная единица нервной системы, приспособленная для осуществления приёма, обработки, хранения, передачи и интеграции информации. Эта клетка состоит из тела, или сомы, и отростков разного типа аксонов – длинные отростки и дендритов короткие отростки. Аксон - прямой отросток, большой длины до 1,5 м, начинающийся от тела клетки. Конец аксона сильно ветвится, образует кисточку называемую окончанием аксона или терминали. Аксон является проводящей частью нейрона, он осуществляет проведение возбуждения от рецептора к нервным клеткам, а от нейрона к исполнительному органу (мышце, железе). Аксон покрыт миелиновой оболочкой, но не сплошь, а отдельными перехватами, что обеспечивает огромную скорость проведения импульса. Аксон покрытый оболочкой называют нервным волокном. Дендриты - короткие, сильно ветвящиеся отростки. От одной клетки может отходить от 1 до 1000 дендритов и они не выходят за пределы Ц.Н.С. У них имеются выросты или ответвления, так называемые шипики. Их очень много только в коре человека, отличительной их особенностью является то, что они подходят близко друг к другу, но не образуют между собой контактов. Но они увеличивают поверхность дендрита и создают условия для размещения на них большого числа контактов с другими нервными клетками. 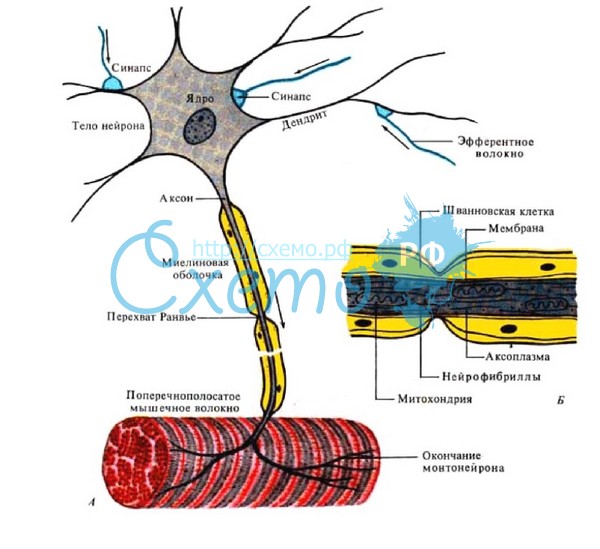 А – строение нейрона, Б – строение нервного волокна (аксона) Классификация нейронов. Нейроны классифицируются по нескольким признакам: 1) по форме тела – звездчатые, веретенообразные, пирамидные и др.; 2) по локализации – центральные (расположены в ЦНС) и периферические (расположены вне ЦНС, а в спинномозговых, черепно-мозговых и вегетативных ганглиях, сплетениях, внутри органов); 3) по числу отростков – униполярные, биполярные и мультиполярные (рис. 3.3.2); 4) по функциональному признаку – рецепторные, эфферентные, вставочные. 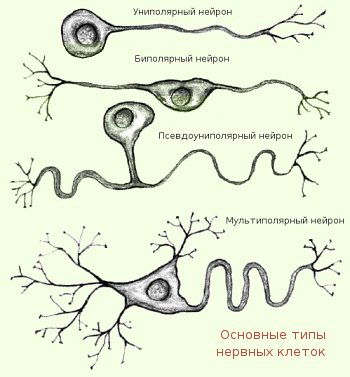 Рецепторные (афферентные, чувствительные) нейроны проводят возбуждение (нервные импульсы) от рецепторов в ЦНС. Тела этих нейронов расположены в спинальных ганглиях, от тела отходит один отросток, который Т-образно делится на две ветви: аксон и дендрит. Дендрит (ложный аксон) – длинный отросток, покрыт миелиновой оболочкой, отходит от тела на периферию, разветвляется, подходя к рецепторам. Эфферентные нейроны (командные по Павлову И.П.) проводят импульсы из ЦНС к органам, эту функцию выполняют длинные аксоны нейронов (длина может достигать 1,5 м.). Их тела располагаются в передних рогах (мотонейроны) и боковых рогах (вегетативные нейроны) спинного мозга. Вставочные (контактные, интернейроны) нейроны – самая многочисленная группа, которые воспринимают нервные импульсы от афферентных нейронов и передают их на эфферентные нейроны. Различают возбуждающие и тормозящие вставочные нейроны. Возрастные особенности. Нервная система формируется на 3-й неделе эмбрионального развития из дорсальной части наружного зародышевого листка – эктодермы. На ранних стадиях развития нейрон имеет большое ядро, окруженное небольшим количеством нейроплазмы, затем оно постепенно уменьшается. На 3-м месяце начинается рост аксона по направлению к периферии и когда он достигает органа, тот начинает функционировать еще во внутриутробном периоде. Дендриты вырастают позднее, начинают функционировать после рождения. По мере роста и развития ребенка увеличивается количество разветвлений на дендритах, на них появляются шипики, что увеличивает количество связей между нейронами. Количество образующихся шипиков прямо пропорционально интенсивности обучения ребенка. У новорожденных количество нейронов больше, чем клеток нейроглии. С возрастом количество глиальных клеток увеличивается и к 20–30 годам соотношение нейронов и нейроглии составляет 50:50. В пожилом и старческом возрасте количество глиальных клеток преобладает в связи с постепенным разрушением нейронов). С возрастом нейроны уменьшаются в размерах, в них уменьшается количество РНК, необходимой для синтеза белков и ферментов. Глиальные клетки – клетки ЦНС, поддерживающие нормальное функционирование нейронов за счёт выполнения механической защиты, опорной, буферной, фагоцитарной, заместительной, изолирующей функции, а также участие в обмене медиаторов. Эти клетки более многочисленны, чем нейроны, занимают объёма ЦНС. Глиальные клетки находятся в непосредственном контакте с нейронами и другими клеточными элементами (мозговыми оболочками, церебральными сосудами). При этом элементы нейроглии образуют оптимальную для нейронов среду. Подобная система строения служит опорой, питанием и разграничением нервных клеток, а также выполняет секреторные функции. Количество глиальных структур значительно больше, чем остальных клеток нервной системы (коэффициент глиоцит/нейрон равен 8-10). Нарушение такого соотношения в одну или другую стороны приводит к развитию различных патологий нервной системы. Особенности происхождения глиальных элементов легли в основу их деления на макроглию (собственно нейроглию) и микроглию. Макроглия неоднородна в морфо-функциональном отношении. К ней относят следующие типы клеток: Эпендимальные; Олигодендроциты; Астроциты. При этом каждая из групп также имеет свои виды клеток. Эпендимальные клетки представлены эпендимоцитами I-го и II-го типов, а также таницитами. Располагаются они в один слой, образуют выстилку мягкой мозговой оболочки (I тип), внутренней поверхности желудочков, цереброспинального канала (II тип) и дно третьего желудочка (танициты). Такое строение обеспечивает выполнение барьерной функции. Олигодендроциты представлены в центральной и в периферической нервной системе. Макроглия наиболее многочисленно представлена именно этими клетками. Виды олигодендроцитов: Центральные глиоциты; Сателиты; Леммоциты. Астроциты – нейроглиальные элементы звездчатой формы с многочисленными отростками. К их особенности относят то, что они представлены только в центральной нервной системе как в белом веществе (протоплазматическая астроглия), так и в сером (волокнистая астроглия). В понятие «нейроглия» также входят микроглиальные клетки или глиальные макрофаги. Они имеют отличное от макроглии не только строение, но и происхождение. Эти особые виды многоотросчатых клеток разбросаны по всему веществу головного мозга и имеют способность к фагоцитозу (такой особенностью обладает и ряд других нейроглиальных элементов). Основная роль глиальных макрофагов состоит в защите церебральных структур от патологических агентов. Глиальные клетки имеют различное происхождение. В зависимости от того, какие клетки явились предшественниками нейроглиальных элементов, выделяют макро- и микроглию. Макроглия развивается из эктодермы (наружного эмбрионального листка), т.е. имеет общих с нейронами предшественников. Микроглиальные макрофаги имеют мезодермальное происхождение (из среднего зародышевого листка). По сути элементы микроглии формируются из структур крови (эритромиелоидов, примитивных макрофагов и других клеток гемоцитарного ростка), заселяющие мозг на ранних этапах эмбриогенеза. В последующем число церебральных макрофагов поддерживается в результате пролиферации. |