срс Биомедицина различие струкртуры прокариот и эукариот. История открытия клетки. Что такое прокариот и эукариот
 Скачать 258.42 Kb. Скачать 258.42 Kb.
|

|
Содержание:
-Транскрипция у прокариот -Трансляция у прокариот
Почти все живые существа состоят из очень маленьких «кирпичиков», которые называются клетками. Большинство клеток настолько малы, что увидеть их можно лишь под мощным микроскопом. При этом каждая клетка может жить сама по себе, создавать похожие на себя клетки и развиваться. Некоторые из живых существ имеют только одну клетку. Это бактерии, одноклеточные животные и одноклеточные растения. Но большинство животных, растений и грибов — многоклеточные. Многоклеточные организмы сложены из множества групп различных клеток, которые имеют свою специализацию и образуют различные органы. Например, в человеческом организме более 220 видов клеток. Одной из важных классификаций в биологии клеток является их деление на прокариоты и эукариоты. Говоря об эволюции микробиологии, стоит отметить существенный вклад ученого Пастера, который был его основоположником. Именно благодаря этому человеку начали развиваться области иммунологии и биотехнологии. Он дал основное определение главным понятиям, относящимся к клетке, обосновал принципы и работу механизма по актуальности роли микроорганизмов во всех сферах жизнедеятельности организмов. Его деятельность продолжил Кох. В своей работе я разберу такие вопросы: какие организмы относятся к каждому из этих двух основных классов клеток. Какое строение имеют клетки и в чем их отличие? Какова классификация каждого из этих видов. Чем же они полезны для человека и биосферы, и каково их значение в целом? История открытия клетки Открытию клетки предшествовало изобретение микроскопа в конце XVI века (З. Янсен). Первым, кто увидел клетки был Р. Гук (1665 г.). С помощью увеличительного прибора он рассматривал срезы тканей живых организмов. На срезе растительной пробки он увидел ячеистую структуру и назвал отдельные ячейки клетками. Гук считал, что сами ячейки — это пустота, а содержимое живого организма заключено в каркасе (клеточной стенке). Чуть позже А. Левенгук, используя более совершенный микроскоп, увидел именно содержимое клеток, в том числе увидел бактерии. В 1827 г К. Бэром была обнаружена яйцеклетка, тем самым было доказано предположение, что все живые организмы развиваются из клетки. Через несколько лет было отрыто содержащееся в клетке ядро (Р. Броун). Обобщив ранее сделанные открытия, Т. Шванн разработал первый вариант клеточной теории, в которой доказывалось единство клеточного строения растений и животных. Однако в клеточной теории Шванна было одно ошибочное предположение, которое было заимствовано у другого исследователя клеток — М. Шлейдена. Оба ученых считали, что клетки могут образовываться из неклеточных структур и веществ. В середине XIX века Р. Вирхов доказал, что все клетки образуются только из других клеток путем их деления («каждая клетка из клетки»). В это же время возникает наука цитология, которая изучает строение и процессы в клетках. Во второй половине XIX века были открыты многие компоненты клетки, отмечена роль ядра в делении клетки. В первой половине XX века с помощью электронного микроскопа были открыты остальные более мелкие структуры клетки. Стало очевидно, что клетки разных организмов и разных тканей имеют много общего. Прокариоты Прокариоты - организмы, состоящие из клеток, которые не имеют клеточного ядра или любых мембранных органелл. Это означает, что генетический материал ДНК у прокариот не связан в ядре. Кроме того, ДНК прокариот менее структурировано, чем у эукариот. В прокариот, ДНК одноконтурный. ДНК эукариот организовано в хромосомы. Большинство прокариот состоят только из одной клетки (одноклеточные), но есть несколько и многоклеточных. Ученые разделяют прокариот на две группы: бактерии и археи. Типичная клетка прокариота содержит следующие части:
 Эукариоты Считается, что ядерные клеточные организмы появились около 1,5 миллиардов лет назад. Хотя в прошлые времена ученые слабо понимали суть явлений на клеточном уровне, но в своих трудах у них часто стали появляться приблизительные рисунки этой единицы организма. Эукариоты - живые организмы, клетки которых содержат ядро и мембранные органеллы. Генетический материал у эукариот находится в ядре, а ДНК организовано в хромосомы. Эукариотические организмы могут быть одноклеточными и многоклеточными. Все животные являются эукариотами. Также эукариоты включают растения, грибы и простейших. Типичная клетка эукариота содержит следующие части:
Существует несколько вариантов деления надцарства эукариот на царства. Первыми были выделены царства растений и животных. Затем было выделено царство грибов, которые, по мнению большинства биологов, не могут быть причислены ни к одному из этих царств из-за биохимических особенностей. Также некоторые авторы выделяют царства протистов или простейших и хромистов. Некоторые системы насчитывают до 20 царств. По системе Томаса Кавалье-Смита, все эукариоты подразделяются на два монофилетических таксона (поддомена) — униконтов (Unikonta) и биконтов (Bikonta). Положение таких эукариот, как Collodictyon и Diphylleia rotans, на данный момент не определено.  Характерные признаки прокариотических и эукариотических клеток
Строение клеток прокариот Клетки прокариотов имеют относительно простое строение. В прокариотической клетке нет настоящего ядра, ядрышка и хромосом. Вместо клеточного ядра есть его эквивалент – нуклеоид (подобное ядру образование), лишённый оболочки и состоящий из одной единственной кольцевой молекулы ДНК, связанной с очень небольшим количеством белка. Это скопление нуклеиновых кислот и белков, лежащих в цитоплазме, и не отделённых от неё мембраной. 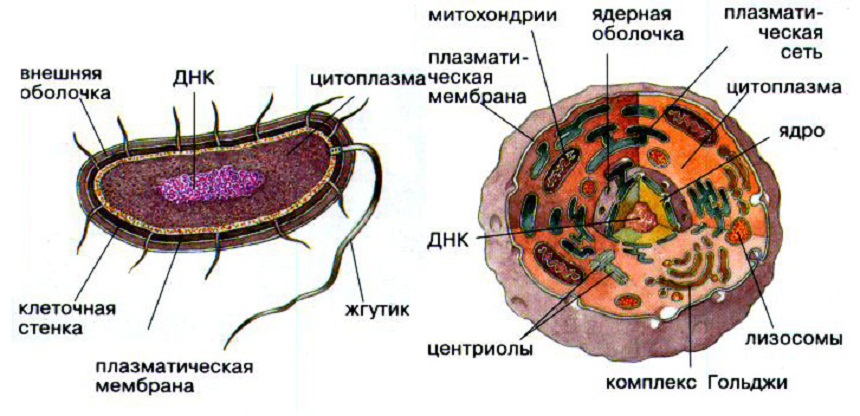 Прокариотические клетки не имеют внутренних мембран, кроме вмятин плазмолеммы. Это означает, что у них отсутствуют такие органеллы как митохондрии, эндоплазматическая сеть, хлоропласты, лизосомы и комплекс Гольджи, которые окружены мембраной и присутствуют в эукариотических клетках. Нет также вакуоль. Из органелл там есть лишь более мелкие, чем у клеток эукариот, рибосомы. Клетки прокариот покрыты плотной клеточной стенкой и часто слизистой капсулой. В состав клеточной стенки входит муреин. Его молекула состоит из параллельно расположенных полисахаридных цепей, сшитых друг с другом короткими цепями пептидов. Плазматическая мембрана может прогибаться внутрь цитоплазмы, образуя мезосомы. На мембранах мезосом расположены окислительно – восстановительные ферменты, а у фотосинтезирующих прокариот – ещё и соответствующие пигменты (бактериохлорофилл у бактерий, хлорофилл и фикобилины у цианобактерий). Благодаря этому такие мембраны способны выполнять функции митохондрий, хлоропластов и других органелл. Бесполое размножение прокариот осуществляется простым делением клетки пополам. Форма клеток бактерий изменчива, однако можно выделить несколько основных морфологических типов: Кокки – шаровидные формы. К коккам относятся: микрококки – одиночные клетки, диплококки – парные кокки; стрептококки – колонии в виде цепочек; стафилококки – гроздевидные колонии; сарцины – колонии кубической формы. Палочки. К палочкам относятся: бактерии (которые, как правило, не образуют споры), бациллы, клостридии (которые образуют споры). Споры у бактерий служат не для размножения, а для перенесения неблагоприятных условий – одна клетка образует одну спору. Споры могут образовываться в центральной части клетки или на одном из концов палочки. Извитые формы. К извитым формам относятся одноклеточные бактерии: спириллы (клетки в виде длинной спирали), вибрионы (клетки, изгиб которых составляет 1/4 спирали). Нитевидные формы. К нитевидным формам относятся как одноклеточные, так и многоклеточные прокариоты. Тело нитевидных прокариот может быть неразветвленным и разветвленным. Многие прокариоты способны к активному движению, которое, как правило, осуществляется с помощью жгутиков. Жгутики прокариот построены на основе белков флагеллинов. Строение клеток эукариот . Клетки эукариот более сложного строения, хотя содержат те же основные структурные компоненты (клеточная стенка, плазмолемма, цитоплазма), что и прокариотические клетки. Все эукариотические клетки разделены на компартменты - реакционные пространства – многочисленными мембранами. В этих отсеках независимо друг от друга одновременно происходят различные химические реакции. В клетке главные функции распределены между ядром и различными органеллами - митохондриями, рибосомами, комплексом Гольджи и пр. Ядро, пластиды и митохондрии отграничены от цитоплазмы двумембранной оболочкой. Ядро клетки содержит генетический материал. Хлоропласты растений в основном выполняют функцию улавливания солнечной энергии и превращают её на химическую энергию углеводов в процессе фотосинтеза, а митохондрии вырабатывают энергию расщепляя углеводы, жиры, белки и другие органические соединения. К мембранным системам цитоплазмы клеток эукариот относятся эндоплазматическая сетка и комплекс Гольджи, необходимые для осуществления жизненных процессов клетки. Лизосомы, пероксисомы и вакуоли так же выполняют специфические функции. Только хромосомы, рибосомы, микротрубочки и микрофиламенты немембранного происхождения. Делятся эукариотические клетки путём митоза. Генетический аппарат прокариот. Прокариоты – это организмы, в клетках которых отсутствует оформленное ядро. Функции ядра выполняет нуклеоид (то есть «подобный ядру»); в отличие от ядра, нуклеоид не имеет собственной оболочки. Основу генома прокариот составляют кольцевые молекулы ДНК: прокариотические хромосомы и плазмиды. Множество молекул ДНК образует две взаимосвязанные подсистемы: хромосомную и плазмидную. Хромосомная подсистема прокариотического генома Основу хромосомной подсистемы прокариотического генома составляет прокариотическая (бактериальная) хромосома (генофор), входящая в состав нуклеоида – ядерноподобной структуры. Нуклеоид по морфологии напоминает соцветие цветной капусты и занимает примерно 30% объема цитоплазмы. Бактериальная хромосома представляет собой кольцевую двуспиральную правозакрученную молекулу ДНК, которая свернута во вторичную спираль. Вторичная структура хромосомы поддерживается с помощью гистоноподобных (основных) белков и РНК. Точка прикрепления бактериальной хромосомы к мезосоме (складке плазмалеммы) является точкой начала репликации ДНК (эта точка носит название сайта OriC). Бактериальная хромосома удваивается перед делением клетки. Репликация ДНК идет в две стороны от сайта OriC и завершается в точке TerC. Молекулы ДНК, способные себя воспроизводить путем репликации, называются репликоны. Длина прокариотической хромосомы составляет несколько миллионов нуклеотидных пар (мпн); например, минимальная длина ДНК прокариотической хромосомы E. coli штамма MG1655 составляет 4639221 пн (физическая длина около 1,5 мм). У типичных прокариот в неделящейся клетке имеется одна бактериальная хромосома. Поэтому прокариоты в целом являются гаплоидами (гаплобионтами). Плазмиды и эписомы - это небольшие фрагменты ДНК, отличающейся от основной массы ДНК. Они часто реплицируются вместе с ДНК хозяина, но не нужны для выживания его клетки. Сначала было принято различать эписомы и плазмиды: эписомы внедряются в ДНК хозяина, а плазмиды - нет. К эписомам относятся F-факторы и так называемые умеренные фаги. Сейчас обе группы называют одним общим термином "плазмиды".Плазмиды широко распространены в природе, и в последние годы их считают внутриклеточными паразитами или симбионтами, устроенными еще проще, чем вирусы. Что касается плазмид, то здесь дело обстоит еще сложнее - ведь они представляют собой только молекулы ДНК. Плазмиды придают своим клеткам-хозяевам целый ряд особых свойств. Некоторые плазмиды являются "факторами резистенции" (R-плазмиды, или R-факторы: от англ. R = resistance - устойчивость), т.е. факторами, придающими устойчивость к антибиотикам. Примером может служить пенициллиназная плазмида стафилококков, которая трансдуцируется различными бактериофагами. В этой плазмиде содержится ген, кодирующий фермент пенициллиназу, которая разрушает пенициллин и, таким образом, придает устойчивость к пенициллину. Передача и распространение таких факторов среди бактерий (в результате полового размножения) очень мешают врачам. Другие плазмидные гены определяют устойчивость к дезинфицирующим средствам; способствуют таким заболеваниям, как стафилококковая импетиго; помогают молочнокислым бактериям превращать молоко в сыр; придают способность усваивать такие сложные вещества, как углеводороды, что можно использовать для борьбы с загрязнениями океана или для получения кормового белка из нефти. Реализация наследственной информации в процессе жизненного цикла (онтогенеза) организма -- двухступенчатый процесс. Сначала с определенных участков ДНК информация подписывается (транскрибируется) в виде комплементарных нулеотидных последовательностей молекул иРНК, которая перемещается в цитоплазму, связывается с рибосомами и в рибосоме с иРНК осуществляется перевод (трансляция) генетической информации в определенную последовательность аминокислотных остатков молекулы белка. Процесс транскрипции находится в клетке под строгим контролем, поэтому имеет место как неодинаковое транскрибирование во времени разных участков ДНК (генов), так и неодинаковая скорость, с которой гены могут транскрибироваться. В результате количество молекул мРНК в клетке, комплементарных разным генам, сильно различается. Хотя в целом механизмы синтеза ДНК и РНК сходны, процесс транскрипции не обладает той степенью точности, которая характерна для репликации ДНК. Однако поскольку мРНК не способна к самовоспроизведению, возникающие при ее синтезе ошибки в последующих клеточных генерациях не воспроизводятся и, следовательно, не могут наследоваться. Матричные РНК служат матрицами для синтеза различных белковых молекул. Перевод генетической информации с "языка" нуклеотидов на "язык" аминокислот--сложный многостадийный процесс, включающий активацию аминокислот, образование ими комплексов с особым видом РНК (транспортными РНК, или тРНК), взаимодействие этих комплексов с мРНК, связанной с рибосомой, приводящее в конечном итоге к формированию полипептидной цепи, аминокислотный состав которой изначально запрограммирован в определенном участке ДНК. В осуществлении каждой из стадий, ведущих к синтезу молекулы белка, участвует несколько различных ферментов. Хотя механизм трансляции отличается высокой точностью, вероятность ошибки в целом выше, чем в случае синтеза молекул ДНК и РНК. Наиболее уязвимый этап -- "узнавание" с помощью фермента аминокислоты соответствующей молекулой тРНК. По имеющимся данным, частота возникновения ошибок на этом этапе порядка 10-4, что и определяет, возможно, уровень точности процесса синтеза белка в целом. Однако, как и в случае синтеза РНК, ошибки в процессе трансляции, приводящие к синтезу измененной молекулы белка, не воспроизводятся, если они не закодированы исходно в генетическом материале. Транскрипция у прокариот.
Трансляция у прокариот.
Таким образом, процессы транскрипции и трансляции, служащие для выражения в онтогенезе генетической информации, не приводят к наследованию изменений, возникающих при их функционировании. Только изменения, происходящие в молекулах ДНК, могут сохраняться в ряду поколений, поскольку они воспроизводятся в процессе репликации. Следовательно, в основе эволюции прокариот лежит способность к изменению только их генетического материала. У прокариот весь генетический материал, необходимый для жизнедеятельности, локализован в одной хромосоме, т. е. бактериальная клетка гаплоидна. В определенных условиях в клетках бактерий может содержаться по нескольку копий хромосомы. Деление прокариотических клеток — процесс образования дочерних прокариотических клеток из материнской. Ключевыми событиями клеточного цикла как прокариот, так и эукариот являются репликация ДНК и деление клетки. Отличительной чертой деления прокариотических клеток является непосредственное участие реплицированной ДНК в процессе деления. В подавляющем большинстве случаев прокариотические клетки делятся с образованием двух одинаковых по размеру дочерних клеток, поэтому этот процесс ещё иногда называют бинарным делением. Так как чаще всего прокариотические клетки имеют клеточную стенку, бинарное деление сопровождается образованием септы — перегородки между дочерними клетками, которая затем расслаивается посередине. При размножении бактерий в искусственных условиях (в ограниченном объеме питательной среды) в развитии культуры выделяется 4 периода, или фазы.
|