Клетка структур един всего живого клетки растит и живот организмов сходны по строению
 Скачать 0.91 Mb. Скачать 0.91 Mb.
|

|
Спиральное. Характеризуется утерей элементов симметрии уже на стадии четырех, (а иногда и двух) бластомеров. В результате наклонного положения митотического веретена все верхние бластомеры сдвигаются в одном направлении по отношению к нижним, т.е. бластомеры анимального полюса располагаются между бластомерами вегетативного полюса. Кольчатые черви, плоские черви, моллюски, немертины. • Билатеральное. На ранних этапах дробления проявляется билатеральная симметрия. При таком дроблении каждый бластомер правой половины эмбриона соответствует подобному бластомеру левой половины, т.е. образующиеся бластомеры располагаются по бокам исходного бластомера. Круглые черви (нематоды, коловратки), асцидии. • Анархическое. Беспорядочное расположение бластомеров. Некоторые медузы. Типы бластул: Целобластулавозникает в результате полного, равномерного дробления. Один слой клеток (бластодерма) расположен по периферии. Это бластула с однослойной стенкой (бластодермой) и обширным бластоцелем. Хар-на для ланцетника, для иглокожих (морские звезды, морские ежи). Амфибластула.Образуется в результате полного неравномерного дробления. Дно бластулы занимает все вегетативное полушарие, а бластоцель смещен в анимальную половину. Хар-на для амфибий и кольчатых червей. Дискобластула. Образуется в результате дискоидального дробления. В телолецитальных яйцах диск бластомеров выгибается над желтком, а между ними возникает полость, которую также называют бластоцелем. Дискоидальное дробление наблюдается лишь в области анимального полюса, где образуется дисковидное скопление клеток - бластодерма От желтка она отделена подзародышевой полостью. Хар-на для птиц и рептилий (пресмыкающихся). 4) Перибластула.Образуется при поверхностном дроблении. Это заключительная стадия дробления в центролецитальных яйцах членистоногих. Бластоцеля нет, он заполнен желтком. Характерна для насекомых, клещей и других членистоногих. 5) Морула. Бластоцель вообще не возникает, дробление заканчивается на стадии плотного комка клеток (напоминает тутовую ягоду). Это ранняя бластула у ланцетника (когда еще нет бластоцеля). Таким образом, у ланцетников морула является предшественником целобластулы. У млекопитающих стадия морулы предшествует бластоцисте. У некоторых животных (например, у некоторых кишечнополостных) морула - это самостоятельный тип бластулы. 6) Стерробластула - это бластула с однослойной стенкой и очень маленьким центрально расположенным бластоцелем. Бластомеры очень крупные, бластоцеля нет или очень маленький. Хар-на для некоторых членистоногих и моллюсков. 7) Плакула. Это двуслойная пластинка. Хар-на для плоских и кольчатых червей. 8) Бластоциста(бластула типа бластодермического пузырька). Хаар-на для млекопитающих. В ней выделяют наружный слой - трофобласт и внутреннюю массу - эмбриобласт. Трофобласт образует ворсинки, с помощью которых бластоциста имплантируется в слизистую матки. Способы гаструляции Инвагинация, или впячивание. Вегетативн полюс бластулы впячивается внутрь, едва не смыкаясь с противоположным. В результ бластоцель почти полностью исчезает и из шара образуется двуслойн зародыш. Внешн слой клеток - эктодерма (наружн листок), а внутрен - энтодерма (внутрен листок). Образующася полость наз полостью первичн кишки, или гастроцелем, вход в нее — первичным ртом, или бластопором. Края бластопора наз губами. Наблюд у животных с изолецитальным типом яиц (ланцетник). Иммиграция . Активн перемещением части клеток бластодермы в бластоцель с формирова¬нием из них внутреннего энтодермального слоя. Характерна для кишечно¬полостных. Эпиболия, или обрастание. Мелкие клетки анимальн полюса более активн делятся, обраст и покрыв снаружи крупные богатые желтком клетки вегетативн полюса, из котор и образ-ся внутрен слой. Хаар-но для умеренно телолецитальн яиц земноводн. Деламинация, или расслоение. Эмбриональн клетки делятся параллельн поверхности бластулы, в результат чего образ наруж и внутрен зародышевые листки. Встреч у птиц, пресмыкающихся и насекомых. 30. эмбрил период онтог. способы формир мезодермы. строен нейрулы. гисто и органогенез эмбриональный (период до вылупления из яйца или до рождения), подразделяющийся на стадии:- зигота; - дробление, завершающееся образованием бластулы; - гаструляция; - нейруляция (эта стадия характерна только для хордовых); - гисто- и органогенез Способы закладки мезодермы Телобластический. С двух сторон первичн кишки вблизи бластопора дифференцируется по 1ой крупн клетке — телобласту, затем кажд из них делится на мелк клетки, котор врастают межд экто- и энтодермой, формируя мезодерму. Хар-н для первичнорот, т.е. наблюдается у всех 3хслойных беспозвоночн животных, кроме иглокожих. Энтероцельный . Мезодерма вычленяется из первичной энтодермы. При этом с двух сторон от первичной кишки образуются выпячивания (карманы). Затем они полностью отделяются от первичной кишки и, разрастаясь между экто- и энтодермой, образуют мезодерму. Характерен для вторичноротых, т.е. наблюдается у иглокожих и хордовых животных. Для высших позвоночных характерен комбинированный способ. НЕЙРУЛА – заключит стадия гаструляции у высших хордовых животных. на этой стадии происход процесс гистогенеза. и орагногенеза. Производные зародышевых листков после гаструляции устанав основн план строен тела животных в виде 3 зародыш листков, и можн говорить о новом этапе в развит зародышей - об органогенезе. зародышевые пласты являются гомологичными образованиями. Производные эктодермы. Из материала наружн зародыш листка - эктодермы - образ нервная пластинка, дающая начало центральн и периферич нервной системе, а также ганглиозная пластинка, из котор формируются ганглии вегетативной нервной системы, клетки мозгового слоя надпочечников, пигментн клетки, задняя доля гипофиза и эпифиз. Из эктодермы развиваются также эпидермис кожи и его производные (волосы, когти, ногти, рога, перья, копыта, панцири, чешуи, чешуйки и т.д), кожн железы (сальные, потовые, молочные и т.д), компоненты органов чувств (органов зрения, слуха, обоняния), эмаль (поверхностный слой) зубов, эпителий ротовой полости и эпителий прямой кишки (т.е. у многих низших животных - эпителий передней и задней кишки). Производные энтодермы. развиваются - эпителий средн кишки, пищеварит железы (печень, поджелудочн железа), эпителий лёгких. Также образ - ротовая полость, передняя доля гипофиза, глоточная или жаберная область, язык, щитовидн железа, зачаток желчного пузыря. У зародыш человека и у всех высших позвоночн животн 1я пара жаберных мешков участв в возникн наружн слух отверстий, полости среднего уха и евстахиевой трубы. 2я пара глоточных щелей у зародыша человека приним участие в образ небных миндалин. За счет энтодермальн клеток, отделяющихся от стенок 3го, 4го и 5го жаберн мешков, развив железы внутрен секреции: тимус (вилочковая железа), околощитовидные железы. Производные мезодермы. В ходе органогенеза происх сложн преобразов клеточн материала мезодермы, приводящ к формиров хрящевого и костного скелета, соединительнотканного слоя кожи, скелетных мышц, а также органов кровенос, выделительн и пол систем организма. Из мезодермы образхорда. 31. гибридологич метод. законы Менделя. Гибридологический метод - Анализ закономерностей наследования отдельных свойств и признаков организмов при половом размножении, а также изменчивости генов и их комбинаторики. Метод разработан Г.Менделем. Принципы гибридологического метода: 1) использование в качестве исходных родительских форм гомозиготных по анализируемым признакам особей (т.е. чистые линии); 2) учет при скрещивании не всего многообразия признаков, а лишь одной или нескольких пар альтернативных вариантов признаков; 3) индивидуальный анализ потомства от каждой особи; 4) количественный учет проявлений изучаемых признаков у всех особей. законы Менделя 1)закон единообразия гибридов первого поколения (1865г) – при скрещивании гомозиготных особей, отличающихся по одной паре альтернативных признаков, все потомки в F1 имеют единый фенотип и генотип, и у всех гибридов проявляется доминантный вариант признака родителей. 2) закон расщепления гибридов второго поколения (1865г.) – при моногибридном скрещивании гетерозиготных особей во втором поколении наблюдается расщепление в отношении 3:1 (3/4 особей с доминантным признаком и 1/4 особей с рецессивным признаком.) по фенотипу, и 1:2:! по генотипу. 3) закон независимого наследования признаков (1865г.) – аллели 2х или нескольких генов, если они располагаются в негомологичных хромосомах, и определяемые ими признаки передаются потомству независимо друг от друга, комбинируются во всевозможных сочетаниях. 9:3:3:1 32. сцепленное наследование. Опыты Моргана. хромосомн теор наследственности. кроссинговер. карты хромосом. Кроссинго́вер -(другое название в биологии перекрёст) — явление обмена участками гомологичных хромосом во время конъюгации при мейозе. Хромосомная теория Основн полож: 1. Материальными носителями наследственной инф является хромосомы с расположенными в них генами. 2. Гены располагаются в хромосомах в линейной последовательностисти. 3. Аллели 1 гена располагаются в идентичных локусах гомологичных хромосом. 4. В гомологичных хромосомах могут располагаться различные аллели одного и того же гена. 5. Набор генов в каждой из негомологичных хромосом уникален. 6. Каждый биологический вид характеризуется определен набором хромосом - кариотипом. 7. Гены, локализованые в одной хромосоме, образуют группу сцепления и, как правило, наследуются вместе. 8.Числ групп сцепления у представит гомогаметного пола (XX) равно n, у представителей гетерогаметного пола (XY) n+1 (n - гаплоидный набор хромосом). 9. После конъюгации в профазе I делен мейоза происходит обмен участками гомологичных хромосом -кроссинговер, что привод к генетической рекомбинации. 10. Сила сцепления между генами обратно пропорциональна расстоянию между ними (чем больше расстояние между генами, тем больше вероятность кроссинговера). 11. За единицу расстояния между генами принимается 1 морганида, которая означает вероятность 1% кроссинговеров. сцепленное наследование сцепл наслед Морган изучал на мухах дрозофилах. изучал наследование 2х признаков: цвет тела: В-серое, в-черное; второй признак длинна крыльев: Wg-длинные, wg-короткие. Морган скрещивал 2 чистые линии: одна с двумя доминантными аллелими, другая-сдвумя рецессивными. Гибриды первого поколения в соответствии с первым законом Менделя – были единообразные. Затем Морган провёл анализирующее скрещивание гибридной самки. он ожидал получить расщепление 1:1:1:1, но получилось 4е фенотипических класса: 2 из которых большие с родительским сочетанием признаков, и два минорных: с новым сочетанием признаков. Такое расщепление происходит в том случае если изучаемые гены располагаются в одной хромосоме образуя группу сцепления. самка образ 4 сорта гамет: 2 не кроссинговерных, 2 кроссинговерных. 33. человек как объект ген исслед. достоинства человека как объекта для ген исслед: 1) высокая численность (особей) доступных для изучения популяции человека; 2)значительное число и разнообразие известных у человека мутации и хромосом анамалий. 3) высокий уровень изученности биохимии и физиологии человека в норме и при различных заболеваниях. недостатки человека как объекта для ген исслед: 1) невозможность целенаправленного подбора родительских пар и экспериментальных браков. 2) низкая плодовитость. 3) высок продолжительности жизни, и как следствие – медленная смена поколений (продолжит жизни поколения равна 25-30 годам, аминимал период жизни до наступлен полов зрелости составл 13-35 лет, что позволяет проследить одному исследователю лишь 2, 3 поколения.) 4) большое число групп сцеплений – 23 и значительно наследованное разнообразие. 5) большая фенотипическая изменчивость людей. Моногенными или менделирующими признаками наз – такие признаки, развитие которых контролируется одним геном. Наследование моногенных признаков, как привило осуществляется в соответствии с законами Менделя. Однако, при этом следует иметь в виду, что мутации разных генов могут приводить в формированию сходных фенотипических проявлений. (признаков) Пигментная КСЕРОДЕРМА развив в резулт нарушений сисемму репарации ДНК, которая устраняет изменения в структуре ДНК (тиминовые димеры), возникающие под действием солнечных лучей. Эта система включ несколько генов, кодирующих синтез ферментов участвующих в репарации ДНК.: 1)Геликаза – разрывает двойные связи а молек ДНК; 2)эндонуклеаза – вырезает поврежденный участок. 3) экзонуклеаза – удаляет поврежденный участок; 4) ДНК-полимераза строит новую копию; 5) лигаза сшивает разрыв сахаро-фосфатного остова Мутация любого из семи генов системы репарации ДНК приводит к развитию заболевания. 34. семейно-генеалогический метод. семейно-генеалогический метод основан на изучении наследования признака в семьях на протяжении ряда поколений. Этот метод применим в том случае, когда сведения об анализирующем признаке сохраняются в памяти у членов семьи или каких то документах. С-г метод получил широк применен в разработке след теоретич и прикладных вопросов: 1) установления наследственного характера признака 2) определения типа наследования; 3) опред пенентрантности 4)изучения сцепления генов и картирования хромосом 5)исследования интенсивности мутацион процесса 6) изучения взаимодействия генов. 7) уточнения генетической природы патологических состояний и составления прогнозов здоровья при медико-генетическом консультировании. При анализе родословных для аутосомно-доминантного типа наследования хар-ны след особенности: 1) признак обнаруж в каждом поколении 3)редкий признак наследуется примерно половиной детей (причина – гетерозиготность) 3) потомки мужского и женского генотипа наследуют одинаково часто 4)оба родителя в равной степени передают признак детям. В родословных где прослеживается наследование аутосом-доминант заболевания, неполная пенетрантность гена, вызывающего это заболевание, будет проявятся так называемым пропуском поколения. Пенетрантность – вероятность появления определенного фенотипа при данном генотипе. Если пенетрантность меньше 100%, говорят о неполной пенетрантности. Неполная пенетрантность может быть результатом межгенных взаимодействий или воздействия факторов окруж среды. Для оценки пенетрантности гена используют метод трехпоколенных цепей. Для этого учитывают все трехпоколенные семьи, происходищии от больных и имеющие больных в 3-м поколении так, чтобы в них не было общих промежуточных предков. Доля промежуточных предков, у которых проявилось заболевание, от общего числа промежуточных предков даёт оценку пенетрантности гена. Варьирующая экспрессивность – понятие, определяющее изменчивость количественного выражения признака у разных индивидуумов-носителей соответствующего аллеля. Аутосомно-рецессивный: 1) при достаточном числе потомков признак может отсутствовать в поколении детей, но появлятся в поколении внуков. 2) признак может развиваться при его отсутствии у родителей. вероятность его проявления составляет 25%. 3) признак наследуется всеми детьми, ели оба родителя его имеют. 4) признак наслед потомками мужского женского пола одинаково часто. Аутосомно-доминантный Геноипы больных: А_(АА или Аа). Генотип здоровых аа . Все одинаково болеют. Нет пропусков поколении. У здоров родит не мог родится больные дети. Вероятность рожд: есл хотябы один из родит гомозиготен по патологическому аллелю. Все дети родж больн независ от генотипа другого родит. Вероятность рожд здоров детей у больных родит составляет 25% если оба родит гетерозигоны по патологическому аллелю. Полидактилия (шестипалость). Брахидактилия (короткопалость). хорея гентингтона, болезнь морфана. семейная гиперхолестеринемия. 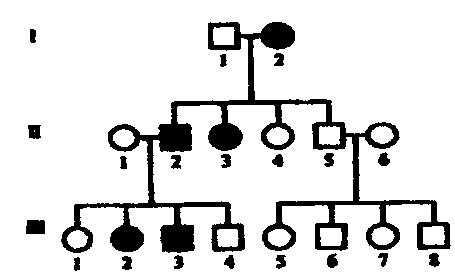 Аутосомно-рецессивный А_(АА-генотипически здоров, Аа-фенотипически здоровый носитель) Одинак часто бол и м и ж. есть пропуски поколен. У здоровых родителей могут родиться больные дети, если оба родителя-носители. Если хотябы один из родит генотипически здоров, все дети будут фенотипически здоровы независимо от генотипа другого родителя.. Если оба родит скрытые носители (здоровы) могут родит больные дети с вероятностью 25%. Вероятн рождения больного ребенка составляе 50% если 1 из род болен. А 2 скрыт носит. Галактоземия, фенилкетонурия, муковисцедоз, синдром тея-сакса, серповид клет анемия. 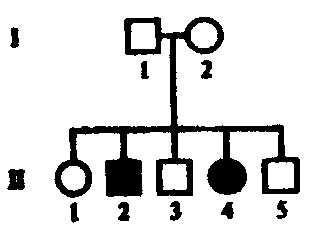 35. хар-ка Х-сцепл доминантного, рецессив и У-сцепл наслед. Х-сцеп доминантный Генотип бол Генотип здоров Чаще бол ж, так как они мог унаслед патологич аллель как от отца, так и от матери, а муж — тольк от матери. Нет пропуск в поколен. Есл хотяб один из родит болен то рожд больн дети. От отца к сыну признак не перед. Больн мать перед палог аллель 100% детей есл она гомозиготна и 50% детей независ от пола, есл она гетерозиготна. 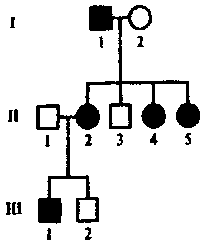 |