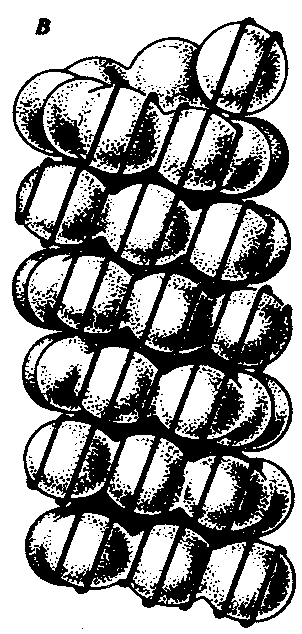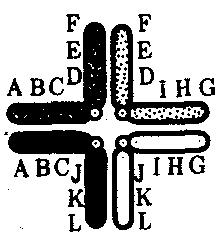биология ярыгин. Книга 1 Издание пятое, исправленное и дополненное
 Скачать 7.35 Mb. Скачать 7.35 Mb.
|

3.4.4. Функциональная характеристика генаВ процессе реализации наследственной информации, заключенной в гене, проявляется целый ряд его свойств. Определяя возможность развития отдельного качества, присущего данной клетке или организму, ген характеризуется дискретностью действия. Ввиду того что в гене заключается информация об аминокислотной последовательности определенного полипептида, его действие является специфичным. Однако в некоторых случаях одна и та же нуклеотидная последовательность может детерминировать синтез не одного, а нескольких полипептидов. Это наблюдается в случае альтернативного сплайсинга у эукариот и при перекрывают генов у фагов и прокариот. Очевидно, такую способность следует оценить как множественное, или плейотропное, действие гена (хотя традиционно под плейотропным действием гена принято понимать участие его продукта — полипептида — в разных биохимических процессах, имеющих отношение к формированию различных сложных признаков). Например, участие фермента (рис. 3.45) в ускорении определенной реакции (А → В), которая является звеном нескольких биохимических процессов, делает зависимыми результаты этих процессов (D и Е) от нормального функционирования гена а, кодирующего этот белок. Определяя возможность транскрибирования мРНК для синтеза конкретной полипептидной цепи, ген характеризуется дозированностью действия, т.е. количественной зависимостью результата его экспрессии от дозы соответствующего аллеля этого гена. Примером может служить зависимость степени нарушения транспортных свойств гемоглобина у человека при серповидно-клеточной анемии от дозы аллеля HbS. Наличие в генотипе человека двойной дозы этого аллеля, приводящего к изменению структуры β-глобиновых цепей гемоглобина, сопровождается грубым нарушением формы эритроцитов и развитием клинически выраженной картины анемии вплоть до гибели. У носителей только одного аллеля HbS при нормальном втором аллеле лишь незначительно изменяется форма эритроцитов и анемия не развивается, а организм характеризуется практически нормальной жизнеспособностью. 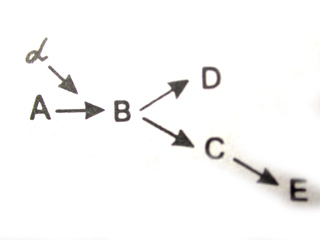 Рис. 3.45. Зависимость формирования нескольких признаков от нормального функционирования продукта гена Нарушение реакции А → В, катализируемой белком, в результате мутации гена ведет к выключению последующих этапов формирования признаков D и Е 3.4.5. Биологическое значение генного уровня организации наследственного материалаДискретность наследственного материала, предположение о которой высказал еще Г. Мендель, подразумевает делимость его на части, являющиеся элементарными единицами,—гены. В настоящее время ген рассматривают как единицу генетической функции. Он представляет собой минимальное количество наследственного материала, которое необходимо для синтеза тРНК, рРНК или полипептида с определенными свойствами. Ген несет ответственность за формирование и передачу по наследству отдельного признака или свойства клеток, организмов данного вида. Кроме того, изменение структуры гена, возникающее в разных его участках, в конечном итоге приводит к изменению соответствующего элементарного признака. Таким образом, на генном уровне организации наследственного материала обеспечиваются индивидуальное наследование и индивидуальное изменение отдельных признаков и свойств клеток, организмов данного вида. Реальное существование генного уровня организации наследственного материала дало возможность исследователям при анализе характера наследования отдельных признаков открыть главные закономерности, которые легли в основу наших представлений об организации материального носителя наследственности и изменчивости. 3.5. ХРОМОСОМНЫЙ УРОВЕНЬ ОРГАНИЗАЦИИ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА3.5.1. Некоторые положения хромосомной теории наследственностиТермин хромосома был предложен в 1888 г. немецким морфологом В. Вальдейером, который применил его для обозначения внутриядерных структур эукариотической клетки, хорошо окрашивающихся основными красителями (от греч. хрома — цвет, краска, и сома — тело). К началу XX в. углубленное изучение поведения этих структур в ходе самовоспроизведения клеток, при созревании половых клеток, при оплодотворении и раннем развитии зародыша обнаружило строго закономерные динамические изменения их организации. Это привело немецкого цитолога и эмбриолога Т. Бовери (1902—1907) и американского цитолога У. Сеттона (1902—1903) к утверждению тесной связи наследственного материала с хромосомами, что легло в основу хромосомной теории наследственности. Детальная разработка этой теории была осуществлена в начале XX в. школой американских генетиков, возглавляемой Т. Морганом. Работы Т. Моргана и его сотрудников не только подтвердили значение хромосом как основных носителей наследственного материала, представленного отдельными генами, но и установили линейность расположения их по длине хромосомы. Доказательством связи материального субстрата наследственности и изменчивости с хромосомами было, с одной стороны, строгое соответствие открытых Г. Менделем закономерностей наследования признаков поведению хромосом в ходе митоза, при мейозе и оплодотворении. С другой стороны, в лаборатории Т. Моргана был обнаружен особый тип наследования признаков, который хорошо объяснялся связью соответствующих генов с Х-хромосомой. Речь идет о сцепленном с полом наследовании окраски глаз у дрозофилы. Представление о хромосомах как носителях комплексов генов было высказано на основе наблюдения сцепленного наследования ряда родительских признаков друг с другом при передаче их в ряду поколений. Такое сцепление неальтернативных признаков было объяснено размещением соответствующих генов в одной хромосоме, которая представляет собой достаточно устойчивую структуру, сохраняющую состав генов в ряду поколений клеток и организмов. Согласно хромосомной теории наследственности, совокупность генов, входящих в состав одной хромосомы, образует группу сцепления. Каждая хромосома уникальна по набору заключенных в ней генов. Число групп сцепления в наследственном материале организмов данного вида определяется, таким образом, количеством хромосом в гаплоидном наборе их половых клеток. При оплодотворении образуется диплоидный набор, в котором каждая группа сцепления представлена двумя вариантами — отцовской и материнской хромосомами, несущими оригинальные наборы аллелей соответствующего комплекса генов. Представление о линейности расположения генов в каждой хромосоме возникло на основе наблюдения нередко возникающей рекомбинации (взаимообмена) между материнским и отцовским комплексами генов, заключенными в гомологичных хромосомах. Было установлено, что частота рекомбинации характеризуется определенным постоянством для каждой пары генов в данной группе сцепления и различна для разных пар. Это наблюдение дало возможность высказать предположение о связи частоты рекомбинации с последовательностью расположения генов в хромосоме и процессом кроссинговера, происходящим между гомологами в профазе I мейоза (см. разд. 3.6.2.3). Представление о линейном распределении генов хорошо объясняло зависимость частоты рекомбинации от расстояния между ними в хромосоме. Открытие сцепленного наследования неальтернативных признаков легло в основу разработки методики построения генетических карт хромосом с использованием гибридологического метода генетического анализа. Таким образом, в начале XX в. была неопровержимо доказана роль хромосом как основных носителей наследственного материала в эука-риотической клетке. Подтверждение этому было получено при изучении химического состава хромосом. 3.5.2. Физико-химическая организация хромосом эукариотической клетки3.5.2.1. Химический состав хромосомИзучение химической организации хромосом эукариотических клеток показало, что они состоят в основном из ДНК и белков, которые образуют нуклеопротеиновый комплекс—хроматин, получивший свое название за способность окрашиваться основными красителями. Как было доказано многочисленными исследованиями (см. § 3.2), ДНК является материальным носителем свойств наследственности и изменчивости и заключает в себе биологическую информацию — программу развития клетки, организма, записанную с помощью особого кода. Количество ДНК в ядрах клеток организма данного вида постоянно и пропорционально их плоидности. В диплоидных соматических клетках организма ее вдвое больше, чем в гаметах. Увеличение числа хромосомных наборов в полипловдных клетках сопровождается пропорциональным увеличением количества ДНК в них. Белки составляют значительную часть вещества хромосом. На их долю приходится около 65% массы этих структур. Все хромосомные белки разделяются на две группы: гистоны и негистоновые белки. Гистоны представлены пятью фракциями: HI, Н2А, Н2В, НЗ, Н4. Являясь положительно заряженными основными белками, они достаточно прочно соединяются с молекулами ДНК, чем препятствуют считыванию заключенной в ней биологической информации. В этом состоит их регуляторная роль. Кроме того, эти белки выполняют структурную функцию, обеспечивая пространственную организацию ДНК в хромосомах (см. разд. 3.5.2.2). Число фракций негистоновых белков превышает 100. Среди них ферменты синтеза и процессинга РНК, редупликации и репарации ДНК. Кислые белки хромосом выполняют также структурную и регуляторную роль. Помимо ДНК и белков в составе хромосом обнаруживаются также РНК, липиды, полисахариды, ионы металлов. РНК хромосом представлена отчасти продуктами транскрипции, еще не покинувшими место синтеза. Некоторым фракциям свойственна регуляторная функция. Регуляторная роль компонентов хромосом заключается в «запрещении» или «разрешении» списывания информации с молекулы ДНК. Массовые соотношения ДНК: гистоны: негистоновые белки: РНК: липиды — равны 1:1:(0,2—0,5):(0,1—0,15):(0,01—-0,03). Другие компоненты встречаются в незначительном количестве. 3.5.2.2. Структурная организация хроматинаСохраняя преемственность в ряду клеточных поколений, хроматин в зависимости от периода и фазы клеточного цикла меняет свою организацию. В интерфазе при световой микроскопии он выявляется в виде глыбок, рассеянных в нуклеоплазме ядра. При переходе клетки к митозу, особенно в метафазе, хроматин приобретает вид хорошо различимых отдельных интенсивно окрашенных телец — хромосом. Интерфазную и метафазную формы существования хроматина расценивают как два полярных варианта его структурной организации, связанных в митотическом цикле взаимопереходами. В пользу такой оценки свидетельствуют данные электронной микроскопии о том, что в основе как интерфазной, так и метафазной формы лежит одна и та же элементарная нитчатая структура. В процессе электронно-микроскопических и физико-химических исследований в составе интерфазного хроматина и метафазных хромосом были выявлены нити (фибриллы) диаметром 3,0—5,0, 10, 20—30 нм. Полезно вспомнить, что диаметр двойной спирали ДНК составляет примерно 2 нм, диаметр нитчатой структуры интерфазного хроматина равен 100—200, а диаметр одной из сестринских хроматид метафазной хромосомы — 500— 600 нм. Наиболее распространенной является точка зрения, согласно которой хроматин (хромосома) представляет собой спирализованную нить. При этом выделяется несколько уровней спирализации (компак-тизации) хроматина (табл. 3.2). Таблица 3.2. Последовательные уровни компактизации хроматина
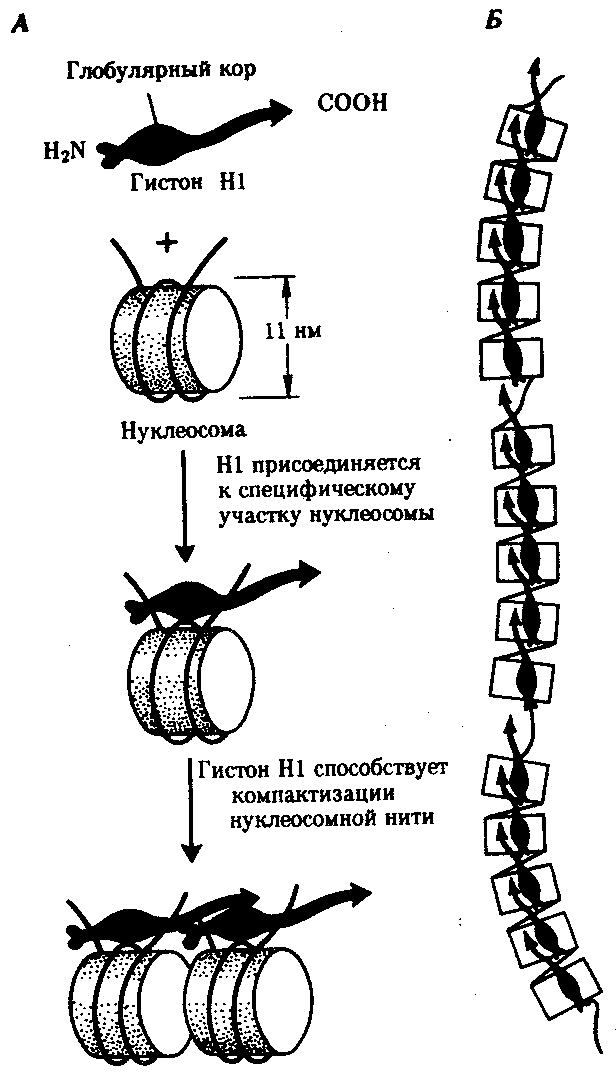
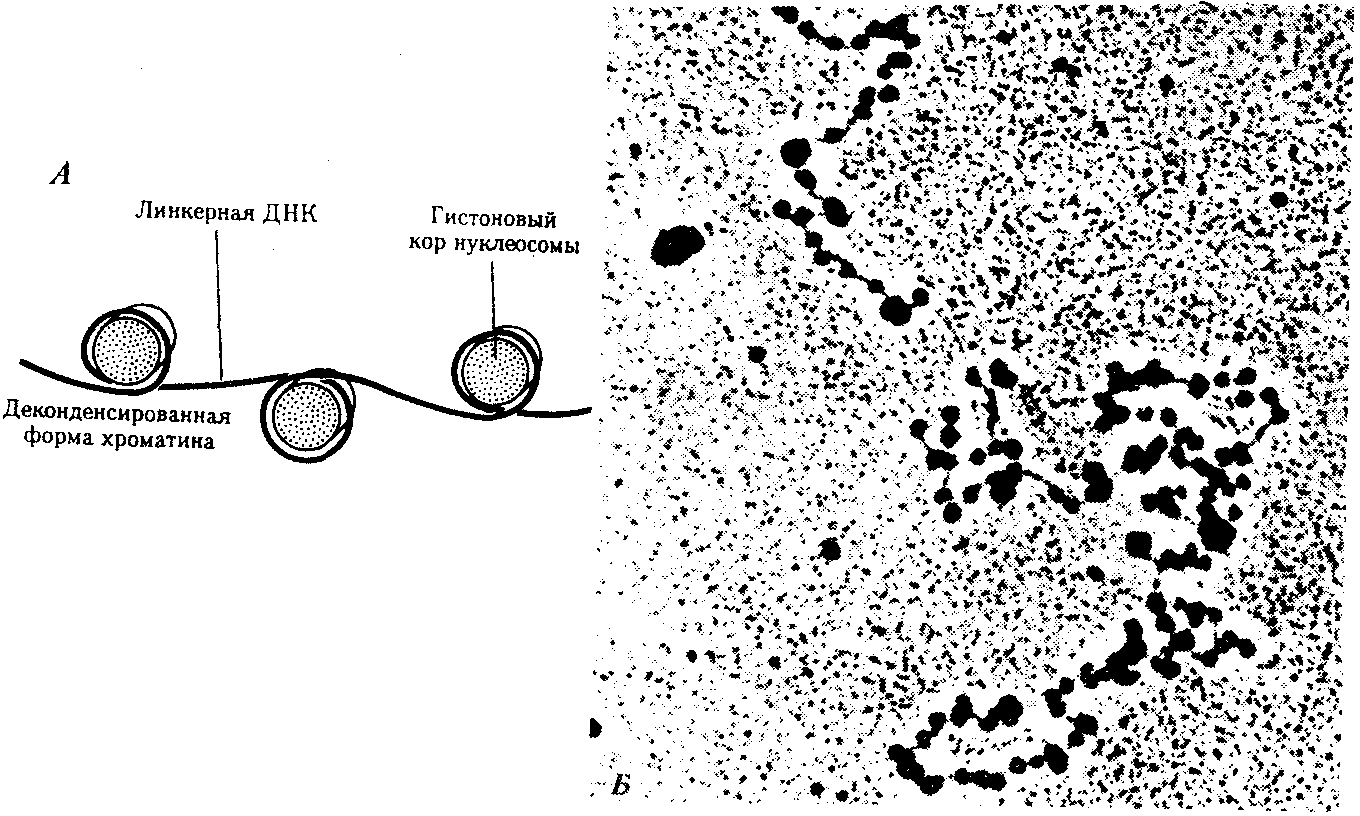 Рис. 3.46. Нуклеосомная организация хроматина. А — деконденсированная форма хроматина; Б — электронная микрофотография эукариотического хроматина: А — молекула ДНК накручена на белковые коры; Б — хроматин представлен нуклеосомами, соединенными линкерной ДНК Нуклеосомиая нить. Этот уровень организации хроматина обеспечивается четырьмя видами нуклеосомных гистонов: Н2А, Н2В, НЗ, Н4. Они образуют напоминающие по форме шайбу белковые тела — коры, состоящие из восьми молекул (по две молекулы каждого вида гистонов) (рис. 3.46). Молекула ДНК комплектируется с белковыми корами, спирально накручиваясь на них. При этом в контакте с каждым кором оказывается участок ДНК, состоящий из 146 пар нуклеотидов (п.н.). Свободные от контакта с белковыми телами участки ДНК называют связующими или линкерными. Они включают от 15 до 100 п.н. (в среднем 60 п.н.) в зависимости от типа клетки. Отрезок молекулы ДНК длиной около 200 п. н. вместе с белковым кором составляет нуклеосому. Благодаря такой организации в основе структуры хроматина лежит нить, представляющая собой цепочку повторяющихся единиц — нуклеосом (рис. 3.46, Б). В связи с этим геном человека, состоящий из 3 · 109 п. н., представлен двойной спиралью ДНК, упакованной в 1,5 · 107 нуклеосом. Вдоль нуклеосомной нити, напоминающей цепочку бус, имеются области ДНК, свободные от белковых тел. Эти области, расположенные с интервалами в несколько тысяч пар нуклеотидов, играют важную роль в дальнейшей упаковке хроматина, так как содержат нуклеотидные последовательности, специфически узнаваемые различными негистоновыми белками. В результате нуклеосомной организации хроматина двойная спираль ДНК диаметром 2 нм приобретает диаметр 10—11 нм. Хроматиновая фибрилла. Дальнейшая компактизация нуклеосомной нити обеспечивается пистоном HI, который, соединяясь с линкерной ДНК и двумя соседними белковыми телами, сближает их друг с другом. В результате образуется более компактная структура, построенная, возможно, по типу соленоида. Такая Хроматиновая фибрилла, называемая также элементарной, имеет диаметр 20—30 нм (рис. 3.47).
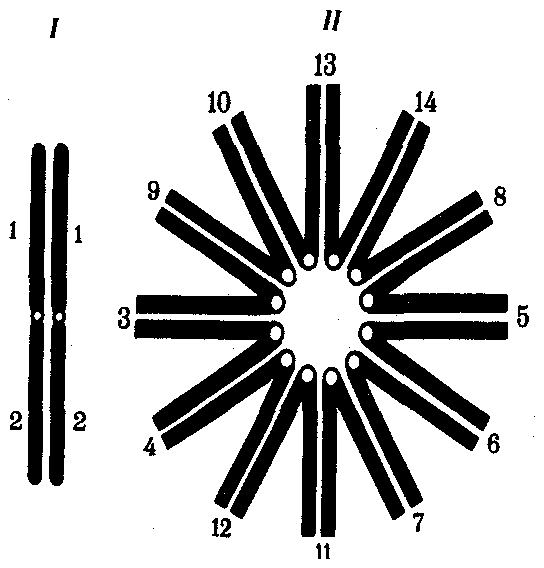
Интерфазная хромонема. Следующий уровень структурной организации генетического материала обусловлен укладкой хроматиновой фибриллы в петли. В их образовании, по-видимому, принимают участие негистоновые белки, которые способны узнавать специфические нуклеотидные последовательности вненуклеосомной ДНК, отдаленные друг от друга на расстояние в несколько тысяч пар нуклеотидов. Эти белки сближают указанные участки с образованием петель из расположенных между ними фрагментов хроматиновой фибриллы (рис. 3.48). Участок ДНК, соответствующий одной петле, содержит от 20 000 до 80 000 п. н. Возможно, каждая петля является функциональной единицей генома. В результате такой упаковки Хроматиновая фибрилла диаметром 20—30 нм преобразуется в структуру диаметром 100—200 нм, называемую интерфазной хромонемой. Отдельные участки интерфазной хромонемы подвергаются дальнейшей компактизации, образуя структурные блоки, объединяющие соседние петли с одинаковой организацией (рис. 3.49). Они выявляются в интерфазном ядре в виде глыбок хроматина. Возможно, существование таких структурных блоков обусловливает картину неравномерного распределения некоторых красителей в метафазных хромосомах, что используют в цитогенетических исследованиях (см. разд. 3.5.2.3 и 6.4.3.6). 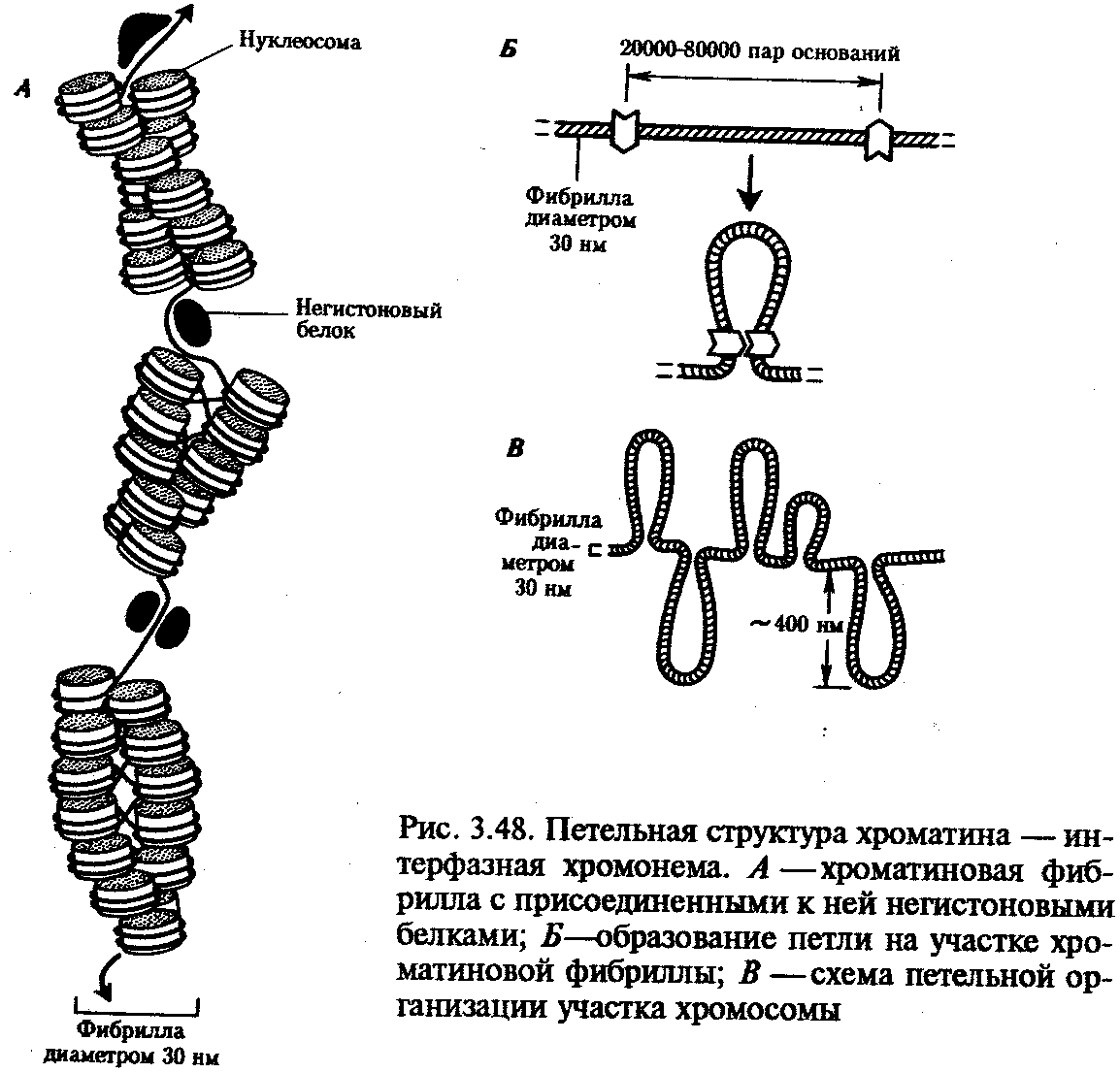 Неодинаковая степень компактизации разных участков интерфазных хромосом имеет большое функциональное значение. В зависимости от состояния хроматина выделяют эухроматиновые участки хромосом, отличающиеся меньшей плотностью упаковки в неделящихся клетках и потенциально транскрибируемые, и гетерохроматиновые участки, характеризующиеся компактной организацией и генетической инертностью. В их пределах транскрипции биологической информации не происходит. Различают конститутивный (структурный) и факультативный гетерохроматин. Конститутивный гетерохроматин содержится в околоцентромерных и теломерных участках всех хромосом, а также на протяжении некоторых внутренних фрагментов отдельных хромосом (рис. 3.50). Он образован только нетранскрибируемой ДНК. Вероятно, его роль заключается в поддержании общей структуры ядра, прикреплении хроматина к ядерной оболочке, взаимном узнавании гомологичных хромосом в мейозе, разделении соседних структурных генов, участии в процессах регуляции их активности. 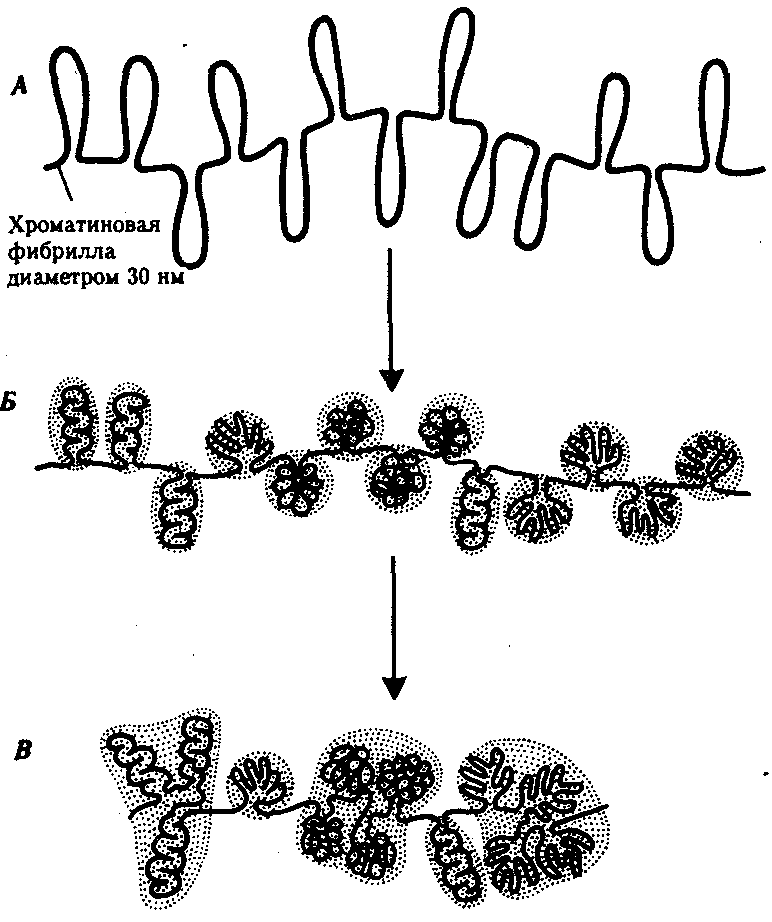 Рис. 3.49. Структурные блоки в организации хроматина. А — петельная структура хроматина; Б — дальнейшая конденсация хроматиновых петель; В — объединение петель, имеющих сходную структуру, в блоки с образованием окончательной формы интерфазной хромосомы  Рис. 3.50. Конститутивный гетерохроматин в метафазных хромосомах человека Примером факультативного гетерохроматина служит тельце полового хроматина, образуемое в норме в клетках организмов гомогаметного пола (у человека гомогаметным является женский пол) одной из двух Х-хромосом. Гены этой хромосомы не транскрибируются. Образование факультативного гетерохроматина за счет генетического материала других хромосом сопровождает процесс клеточной дифференцировки и служит механизмом выключения из активной функции групп генов, транскрипция которых не требуется в клетках данной специализации. В связи с этим рисунок хроматина ядер клеток из разных тканей и органов на гистологических препаратах различается. Примером может служить гетерохроматизация хроматина в ядрах зрелых эритроцитов птиц. Перечисленные уровни структурной организации хроматина обнаруживаются в неделящейся клетке, когда хромосомы еще недостаточно компактизованы, чтобы быть видимыми в световой микроскоп как отдельные структуры. Лишь некоторые их участки с более высокой плотностью упаковки выявляются в ядрах в виде хроматиновых глыбок (рис. 3.51). 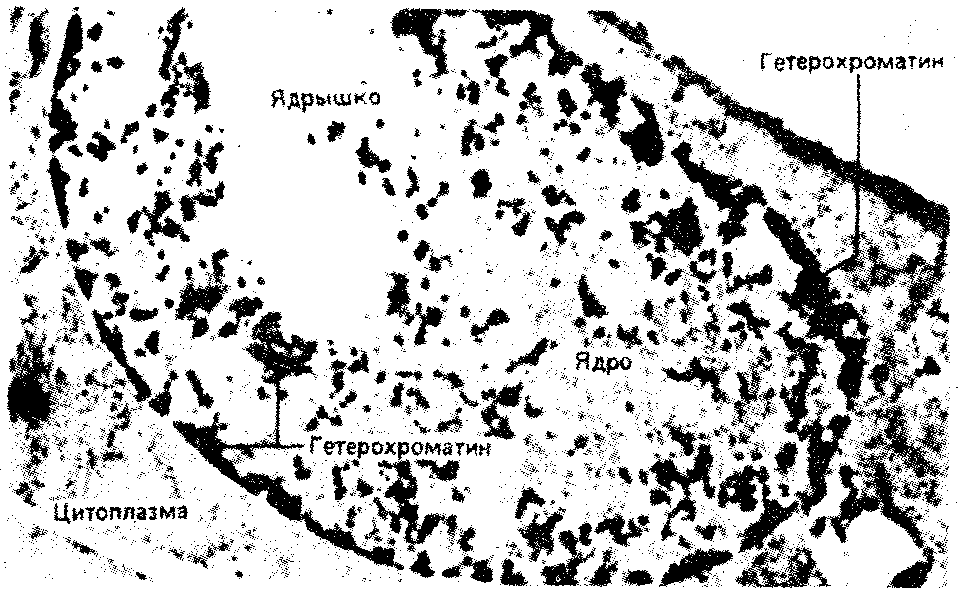 Рис. 3.51. Гетерохроматин в интерфазном ядре Компактные участки гетерохроматина сгруппированы около ядрышка и ядерной мембраны Метафазная хромосома. Вступление клетки из интерфазы в митоз сопровождается суперкомпактизацией хроматина. Отдельные хромосомы становятся хорошо различимы. Этот процесс начинается в профазе, достигая своего максимального выражения в метафазе митоза и анафазе (см. разд. 2.4.2). В телофазе митоза происходит декомпак-тизация вещества хромосом, которое приобретает структуру интерфазного хроматина. Описанная митотическая суперкомпактизация облегчает распределение хромосом к полюсам митотического веретена в анафазе митоза. Степень компактизации хроматина в разные периоды митотического цикла клетки можно оценить по данным, приведенным в табл. 3.2. 3.5.2.3. Морфология хромосомМитотическая суперкомпактизация хроматина делает возможным изучение внешнего вида хромосом с помощью световой микроскопии. В первой половине митоза они состоят из двух хроматид, соединенных между собой в области первичной перетяжки (центромеры или кинетохора) особым образом организованного участка хромосомы, общего для обеих сестринских хроматид. Во второй половине митоза происходит отделение хроматид друг от друга. Из них образуются однонитчатые дочерние хромосомы, распределяющиеся между дочерними клетками. В зависимости от места положения центромеры и длины плеч, расположенных по обе стороны от нее, различают несколько форм хромосом: равноплечие, или метацентрические (с центромерой посередине), неравноплечие, или субметацентрические (с центромерой, сдвинутой к одному из концов), палочковидные, или акроцентрические (с центромерой, расположенной практически на конце хромосомы), и точковые —очень небольшие, форму которых трудно определить (рис. 3.52). При рутинных методах окраски хромосом они различаются по форме и соотносительным размерам. При использовании методик дифференциальной окраски выявляется неодинаковая флуоресценция или распределение красителя по длине хромосомы, строго специфические для каждой отдельной хромосомы и ее гомолога (рис. 3.53). Таким образом, каждая хромосома индивидуальна не только по заключенному в ней набору генов, но и по морфологии и характеру дифференциального окрашивания. 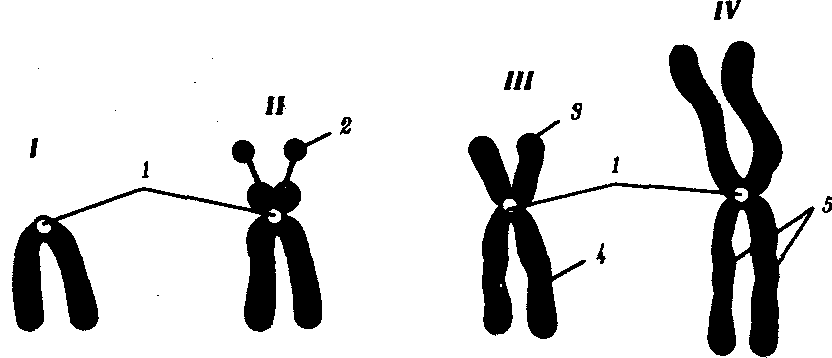 Рис. 3.52. Формы хромосом: I — телоцентрическая, II — акроцентрическая, III—субметацентрическая, IV—метацентрическая; 1 — центромера, 2 — спутник, 3 — короткое плечо, 4 — длинное плечо, 5 — хроматиды 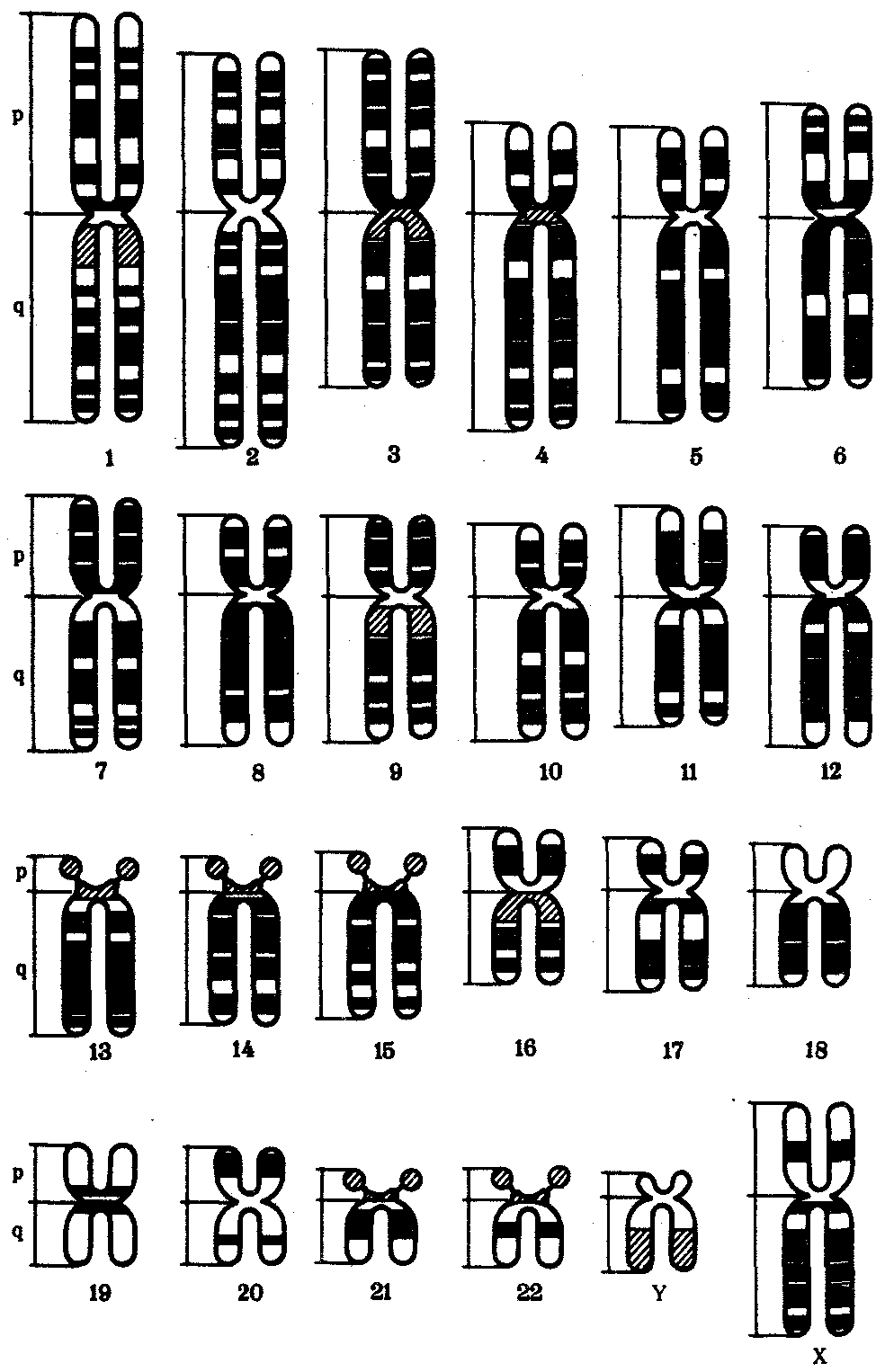 Рис. 3.53. Расположение локусов в хромосомах человека при их дифференциальном окрашивании: р — короткое плечо, q — длинное плечо; 1—22 — порядковый номер хромосомы; XY — половые хромосомы 3.5.2.4. Особенности пространственной организациигенетического материала в прокариотической клеткеОписанная организация наследственного материала эукариот, связанная с расположением его функциональных единиц в ядерных структурах—хромосомах, у прокариот отсутствует. Основная часть генетического материала заключена у них в единственной кольцевой молекуле ДНК длиной около 1 мм (Е. coli), причем у прокариот не обнаружено гистонов, обеспечивающих нуклеосомную организацию хроматина эукариот. Однако кольцевой молекуле ДНК прокариот также присуща укладка в виде петель. ДНК прокариот тоже комплексируется с некоторыми негастоновыми белками, образуя нуклеоид, который гораздо менее устойчив по сравнению с хроматином эукариот (рис. 3.54). 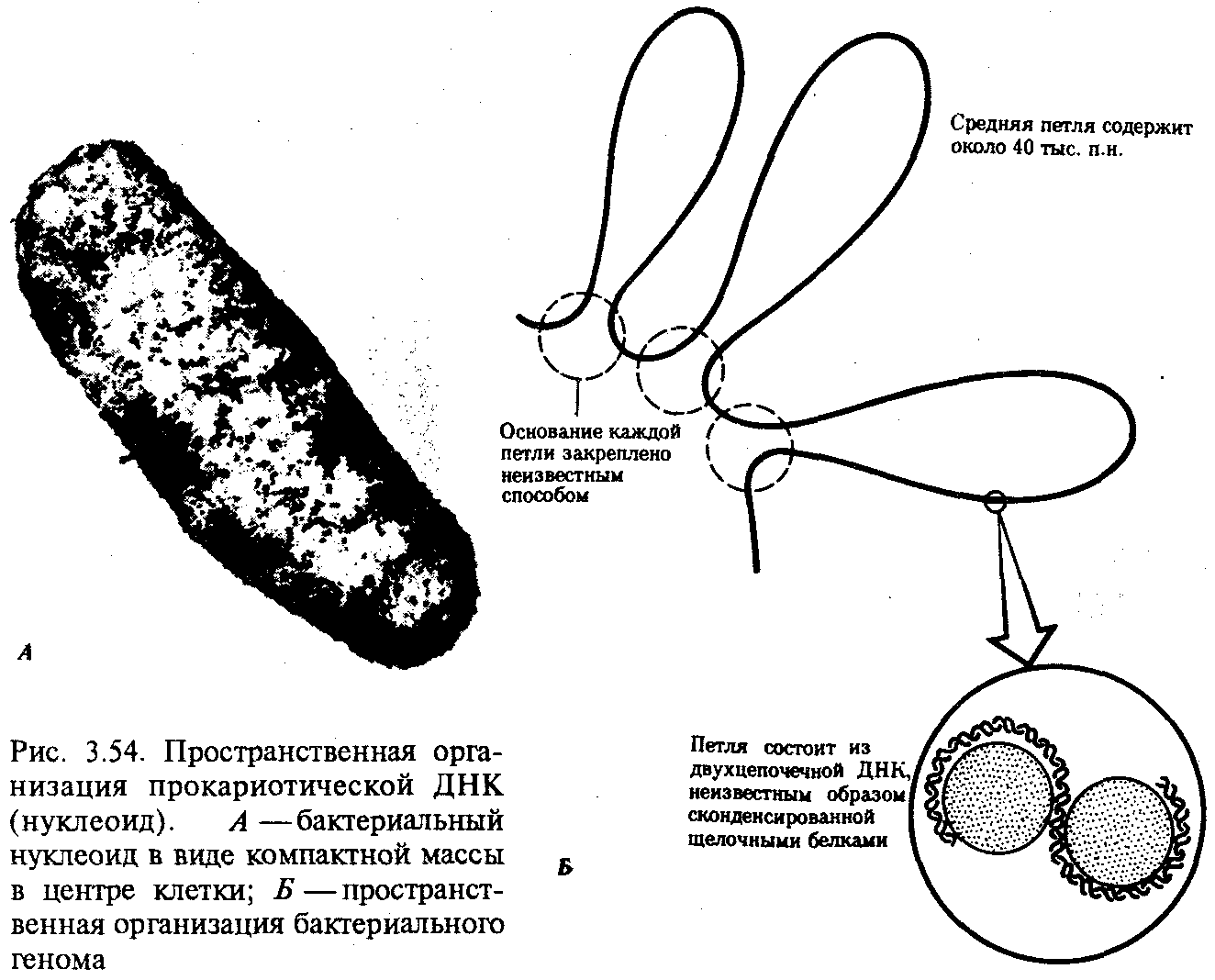 3.5.3. Проявление основных свойств материала наследственности и изменчивости на хромосомном уровне его организацииНа хромосомном уровне организации, который появляется в процессе эволюции у эукариотических клеток, генетический аппарат должен удовлетворять всем требованиям, предъявляемым к субстрату наследственности и изменчивости: обладать способностью к самовоспроизведению, поддержанию постоянства своей организации и приобретению изменений, которые могут передаваться новому поколению клеток. 3.5.3.1. Самовоспроизведение хромосомв митотическом цикле клеток Рис. 3.55. Возникновение двунитчатой хромосомы из однонитчатой: I — интерфазная хромосома до репликации ДНК, II — интерфазная хромосома после репликации ДНК, III — метафазная хромосома Процесс самовоспроизведения — редупликации хромосом осуществляется в период подготовки клетки к митозу — в интерфазе. Вследствие этого в ходе после- дующего деления клетка передает дочернему поколению полноценный комплекс генов, заключенный в каждой отдельной хромосоме. В основе самовоспроизведения хромосомы лежит репликация ДНК с образованием из одной материнской спирали двух дочерних молекул, на базе которых образуются две сестринские хроматиды (рис. 3.55). Благодаря высокой точности репликации ДНК (см. разд. 3.4.2.2 ) сестринские хроматиды несут практически одинаковую информацию, записанную в ее дочерних молекулах. В каждой хроматиде ДНК находится в комплексе с гистонами, и ей присущи все описанные выше уровни компактизации хроматина, свойственные интерфазной клетке. Таким образом, готовясь к самовоспроизведению, клетка удваивает содержание ДНК в каждой хромосоме. При этом последняя приобретает двунитчатую структуру. Известны случаи, когда репликация ДНК, причем неоднократная, не сопровождается формированием хроматид на базе дочерних спиралей. При этом образуются так называемые политенные хромосомы, содержащие многие сотни копий ДНК. Такие хромосомы обнаруживаются, например, в неделящихся клетках слюнных желез личинок некоторых насекомых при обычной световой микроскопии (рис. 3.56). 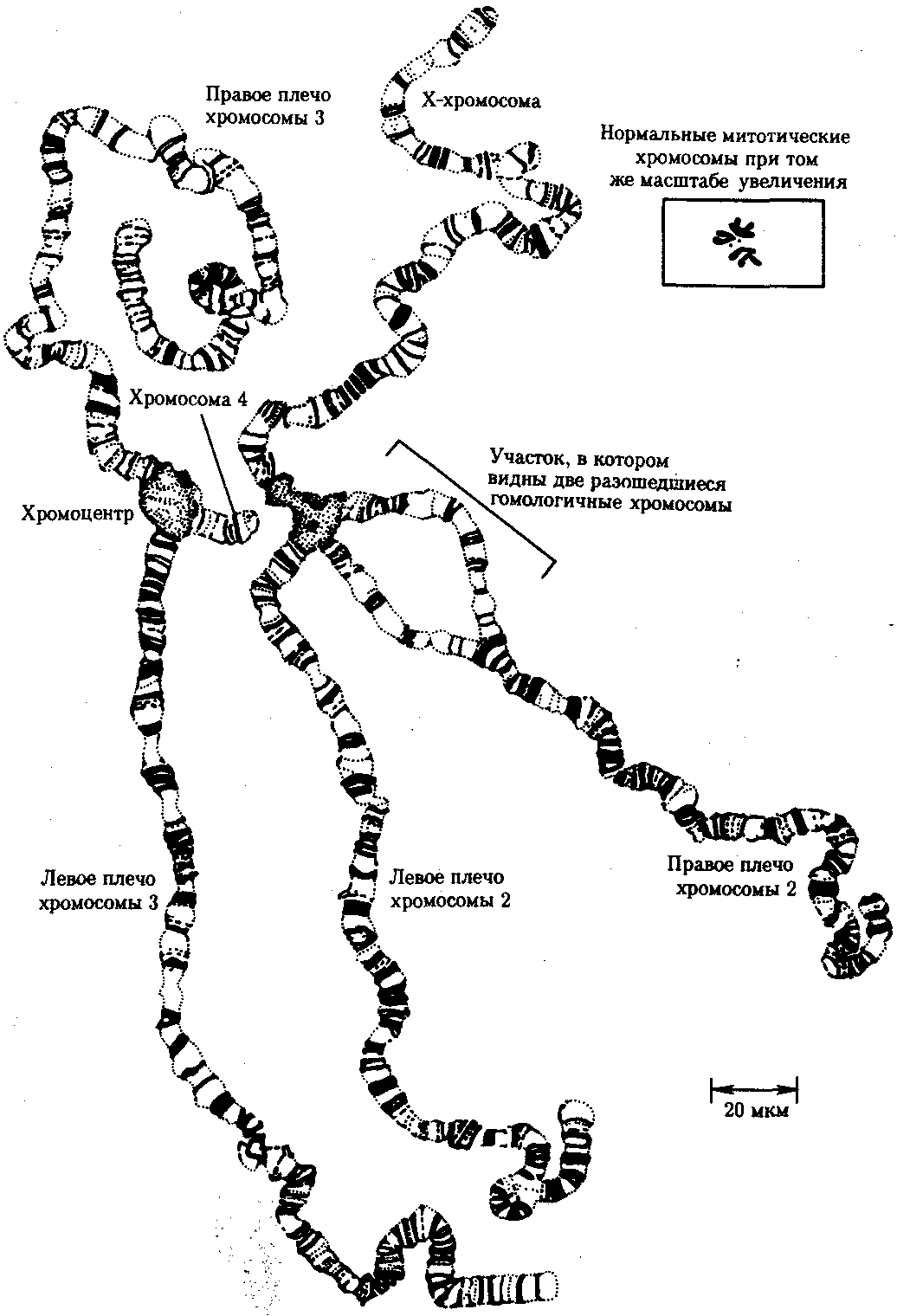 Рис. 3.56. Полный набор политенных хромосом из клетки слюнной железы дрозофилы 3.5.3.2. Распределение материала материнских хромосоммежду дочерними клетками в митозеВ ходе митотического деления обеспечивается закономерное распределение сестринских хроматид каждой хромосомы между дочерними клетками. В составе дочерних хромосом (бывших сестринских хроматид) каждая клетка нового поколения получает одну из двух молекул ДНК, образовавшихся в результате репликации материнской двойной спирали. Следовательно, новое поколение клеток получает одинаковую генетическую информацию в составе каждой группы сцепления. Таким образом, процессы, происходящие с хромосомами при подготовке клеток к делению и в самом делении, обеспечивают самовоспроизведение и постоянство их структуры в ряду клеточных поколений (см. разд. 3.6.2.1). После митоза хромосомы дочерней клетки представлены одной молекулой ДНК, компактно упакованной с помощью белков в одну хроматиновую нить, т.е. имеют такую же структуру, какую имели хромосомы материнской клетки до начала процесса репликации ДНК. Если вновь образованная клетка выбирает путь подготовки к делению, то в ней должны произойти все описанные выше события, связанные с динамикой структурной организации ее хромосом. 3.5.3.3. Изменения структурной организации хромосом. Хромосомные мутацииНесмотря на эволюционно отработанный механизм, позволяющий сохранять постоянной физико-химическую и морфологическую организацию хромосом в ряду клеточных поколений, под влиянием различных воздействий эта организация может изменяться. В основе изменения структуры хромосомы, как правило, лежит первоначальное нарушение ее целостности — разрывы, которые сопровождаются различными перестройками, называемыми хромосомными мутациями или аберрациями. Разрывы хромосом происходят закономерно в ходе кроссинговера, когда они сопровождаются обменом соответствующими участками между гомологами (см. разд. 3.6.2.3). Нарушение кроссинговера, при котором хромосомы обмениваются неравноценным генетическим материалом, приводит к появлению новых групп сцепления, где отдельные участки выпадают — делении — или удваиваются — дупликации (рис. 3.57). При таких перестройках изменяется число генов в группе сцепления. Разрывы хромосом могут возникать также под влиянием различных мутагенных факторов, главным образом физических (ионизирующего и других видов излучения), некоторых химических соединений, вирусов. 128 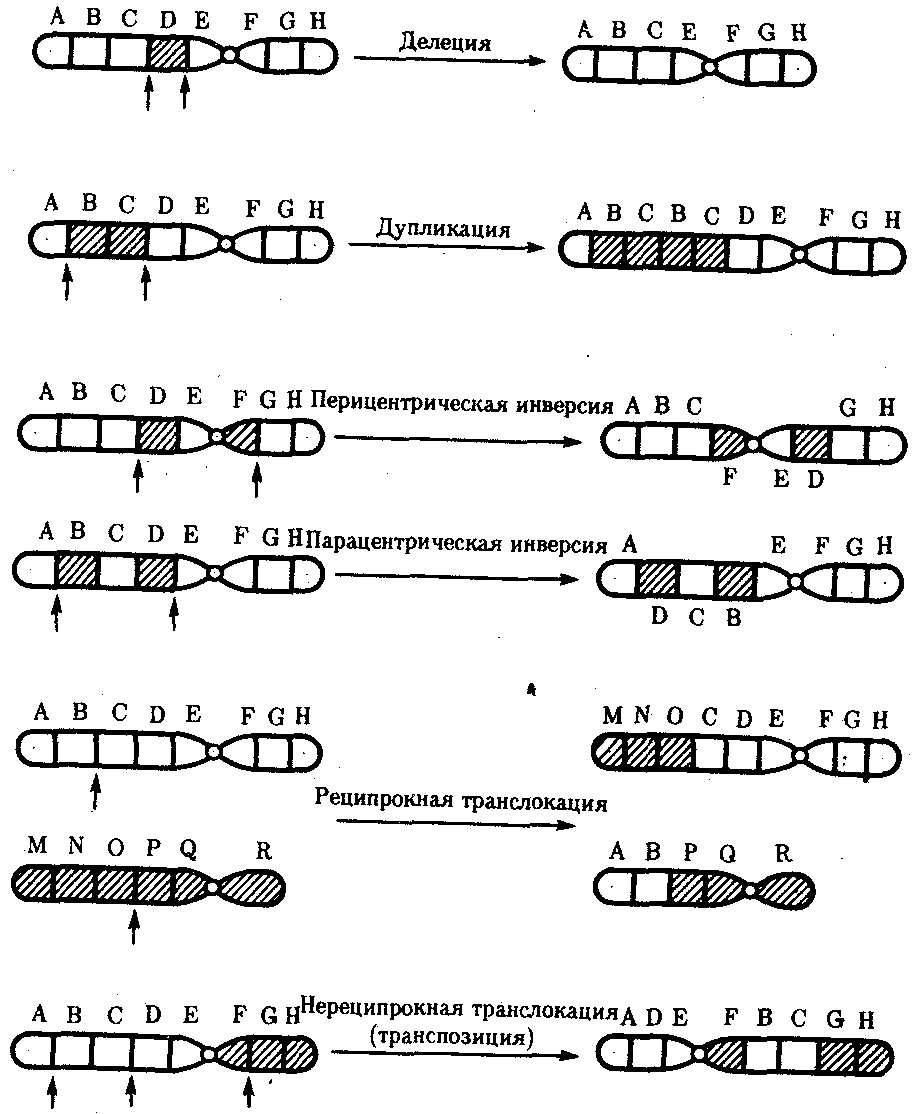 Рис. 3.57. Виды хромосомных перестроек Нарушение целостности хромосомы может сопровождаться поворотом ее участка, находящегося между двумя разрывами, на 180° — инверсия. В зависимости от того, включает ли данный участок область центромеры или нет, различают перицентрические и парацентрические инверсии (рис. 3.57). Фрагмент хромосомы, отделившийся от нее при разрыве, может быть утрачен клеткой при очередном митозе, если он не имеет центромеры. Чаще такой фрагмент прикрепляется к одной из хромосом — транслокация. Нередко две поврежденные негомологичные хромосомы взаимно обмениваются оторвавшимися участками —ре-ципрокная транслокация (рис. 3.57). Возможно присоединение фрагмента к своей же хромосоме, но в новом месте — транспозиция (рис. 3.57). Таким образом, различные виды инверсий и транслокаций характеризуются изменением локализации генов. Хромосомные перестройки, как правило, проявляются в изменении морфологии хромосом, что можно наблюдать в световой микроскоп. Метацентрические хромосомы превращаются в субметацентрические и акроцентрические и наоборот (рис. 3.58), появляются кольцевые и полицентрические хромосомы (рис. 3.59). Особую категорию хромосомных мутаций представляют аберрации, связанные с центрическим слиянием или разделением хромосом, когда две негомологичные структуры объединяются в одну — робертсоновская транслокация, или одна хромосома образует две самостоятельные хромосомы (рис. 3.60). При таких мутациях не только появляются хромосомы с новой морфологией, но и изменяется их количество в кариотипе. 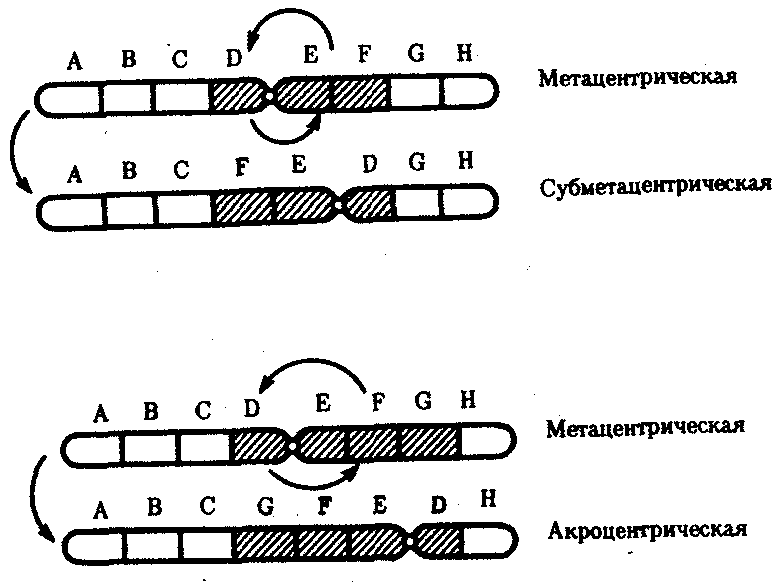 Рис. 3.58. Изменение формы хромосом в результате перицентрических инверсий 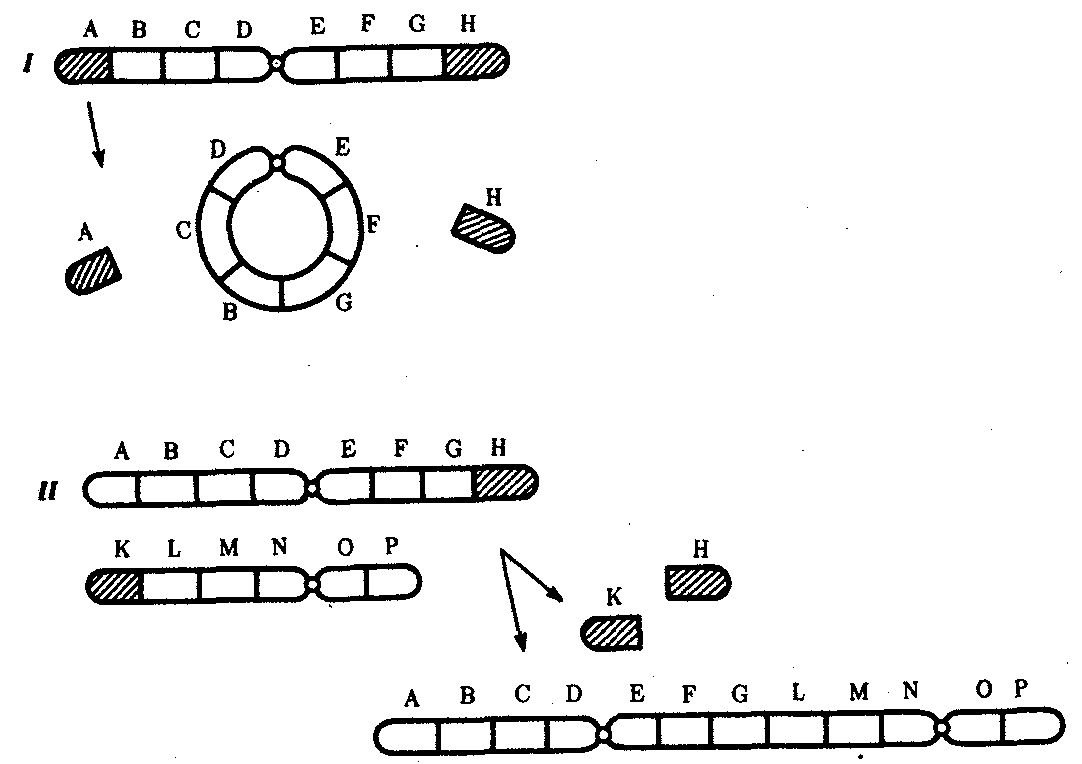 Рис. 3.59. Образование кольцевых (I) и полицентрических (II) хромосом 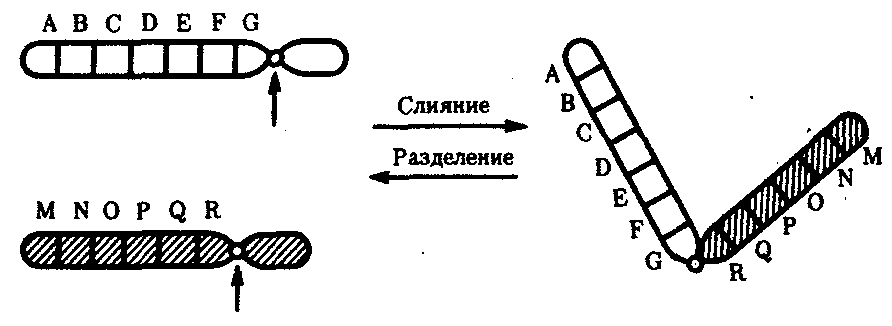 Рис. 3.60. Хромосомные перестройки, связанные с центрическим слиянием или разделением хромосом являются причиной изменения числа хромосом в кариотипе 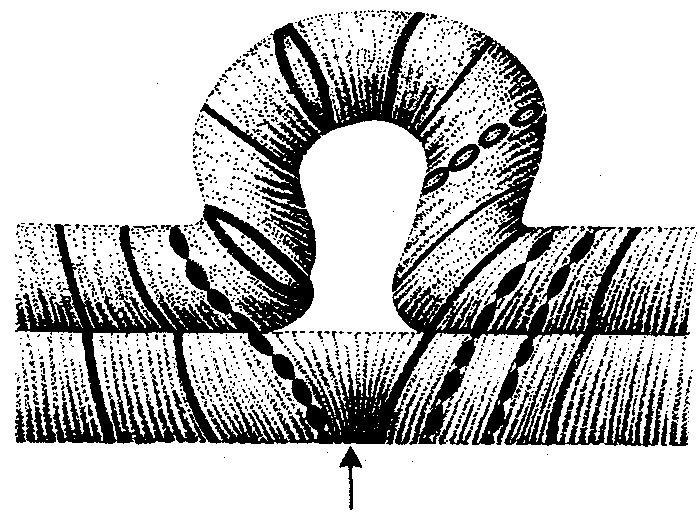 Рис. 3.61. Петля, образующаяся при конъюгации гомологичных хромосом, которые несут неравноценный наследственный материал в соответствующих участках в результате хромосомной перестройки Описанные структурные изменения хромосом, как правило, сопровождаются изменением генетической программы, получаемой клетками нового поколения после деления материнской клетки, так как изменяется количественное соотношение генов (при делениях и дупликациях), меняется характер их функционирования в связи с изменением взаимного расположения в хромосоме (при инверсии и транспозиции) или с переходом в другую группу сцепления (при транслокации). Чаще всего такие структурные изменения хромосом отрицательно сказываются на жизнеспособности отдельных соматических клеток организма, но особенно серьезные последствия имеют хромосомные перестройки, происходящие в предшественниках гамет. Изменения структуры хромосом в предшественниках гамет сопровождаются нарушением процесса конъюгации гомологов в мейозе и их последующего расхождения. Так, делении или дупликации участка одной из хромосом сопровождаются при конъюгации образованием петли гомологом, имеющим избыточный материал (рис. 3.61). Реципрокная транслокация между двумя негомологичными хромосомами приводит к образованию при конъюгации не бивалента, а квадривалента, в котором хромосомы образуют фигуру креста благодаря притягиванию гомологичных участков, расположенных в разных хромосомах (рис. 3.62). Участие в реципрокных транслокациях большего числа хромосом с образованием поливалента сопровождается формированием еще более сложных структур при конъюгации (рис. 3.63).
В случае инверсии бивалент, возникающий в профазе I мейоза, образует петлю, включающую взаимно инвертированный участок (рис. 3.64). Конъюгация и последующее расхождение структур, образованных измененными хромосомами, приводит к появлению новых хромосомных перестроек. В результате гаметы, получая неполноценный наследственный материал, не способны обеспечить формирование нормального организма нового поколения. Причиной этой является нарушение соотношения генов, входящих в состав отдельных хромосом, и их взаимного расположения. Однако, несмотря на неблагоприятные, как правило, последствия хромосомных мутаций, иногда они оказываются совместимыми с жизнью клетки и организма и обеспечивают возможность эволюции структуры хромосом, лежащей в основе биологической эволюции. Так, небольшие по размеру делении могут сохраняться в гетерозиготном состоянии в ряду поколений. Менее вредными, чем делении, являются дупликации, хотя большой объем материала в увеличенной дозе (более 10% генома) приводит к гибели организма. 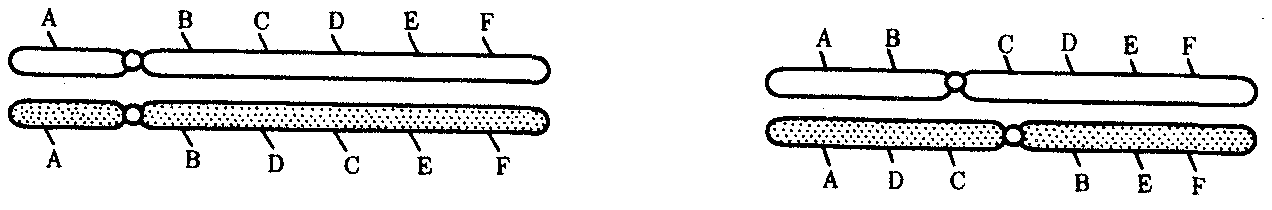 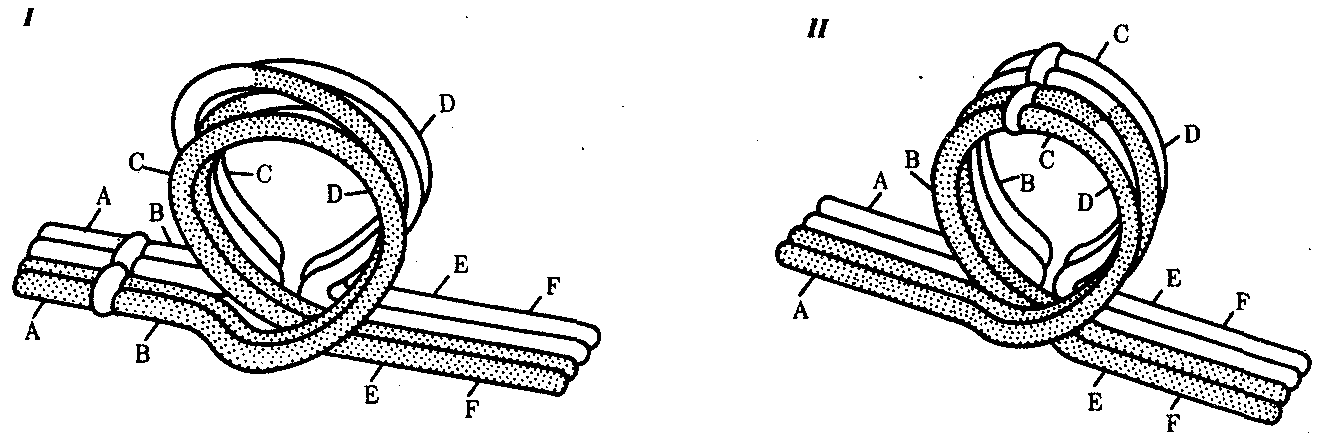 Рис. 3.64. Конъюгация хромосом при инверсиях: I — парацентрическая инверсия в одном из гомологов, II — перидентрическая инверсия в одном из гомологов Нередко жизнеспособными оказываются робертсоновские транслокации, часто не связанные с изменением объема наследственного материала. Этим можно объяснить варьирование числа хромосом в клетках организмов близкородственных видов. Например, у разных видов дрозофилы количество хромосом в гаплоидном наборе колеблется от 3 до 6, что объясняется процессами слияния и разделения хромосом. Возможно, существенным моментом в появлении вида Homo sapiens были структурные изменения хромосом у его обезьяноподобного предка. Установлено, что два плеча крупной второй хромосомы человека соответствуют двум разным хромосомам современных человекообразных обезьян (12-й и 13-й —шимпанзе, 13-й и-14-й —гориллы и орангутана). Вероятно, эта человеческая хромосома образовалась в результате центрического слияния по типу робертсоновской транслокации двух обезьяньих хромосом. К существенному варьированию морфологии хромосом, лежащему в основе их эволюции, приводят транслокации, транспозиции и инверсии. Анализ хромосом человека показал, что его 4, 5, 12 и 17-я хромосомы отличаются от соответствующих хромосом шимпанзе перицентрическими инверсиями. Таким образом, изменения хромосомной организации, чаще всего оказывающие неблагоприятное воздействие на жизнеспособность клетки и организма, с определенной вероятностью могут быть перспективными, наследоваться в ряду поколений клеток и организмов и создавать предпосылки для эволюции хромосомной организации наследственного материала. 3.5.4. Значение хромосомной организации в функционированиии наследовании генетического аппаратаХромосомный уровень организации наследственного материала обеспечивает в эукариотической клетке не только определенный характер функционирования отдельных генов, тип их наследования, но и регуляцию их активности. Хромосома как комплекс генов представляет собой эволюционно сложившуюся структуру, свойственную всем особям данного вида. Взаимное расположение генов в составе хромосомы играет немаловажную роль в характере их функционирования. Расположение гена в той или иной хромосоме определяет тип наследования соответствующего признака (см. разд. 6.3.1). Принадлежность генов к одной хромосоме обусловливает сцепленный характер наследования детерминируемых ими признаков, а расстояние между генами влияет на частоту рекомбинации этих признаков в потомстве (правило Т. Моргана). Расположение генов в разных хромосомах служит основой независимого наследования признаков (закон независимого наследования признаков Г. Менделя). Образуя в хромосоме устойчивый комплекс с гистонами, ДНК эукариотической клетки оказывается недоступной для других белков, осуществляющих транскрипцию (РНК-полимераза) и выполняющих регуляторные функции (см. разд. 3.6.6.4). У прокариот кольцевая молекула ДНК, формирующая неустойчивый комплекс с негистоновыми белками, остается доступной для белков, активирующих отдельные гены путем прямого взаимодействия с определенными нуклеотидными последовательностями ДНК. Таким образом, гистоны, участвующие в пространственной организации ДНК в хромосоме, регулируют генную активность, угнетая ее. Процесс транскрипции с участка ДНК эукариотической клетки предполагает необходимость предварительной декомпактизации хроматина путем временного ослабления связи ДНК с этими белками. Примером такой декомпактизации хроматина являются пуфы (вздутия), наблюдаемые в участках политенных хромосом и соответствующие активно функционирующим генам (рис. 3.65) или обнаруженные в овоцитах амфибий хромосомы типа ламповых щеток, в которых активные участки ДНК формируют петлеобразные структуры с меньшей степенью компактизации (рис. 3.66). 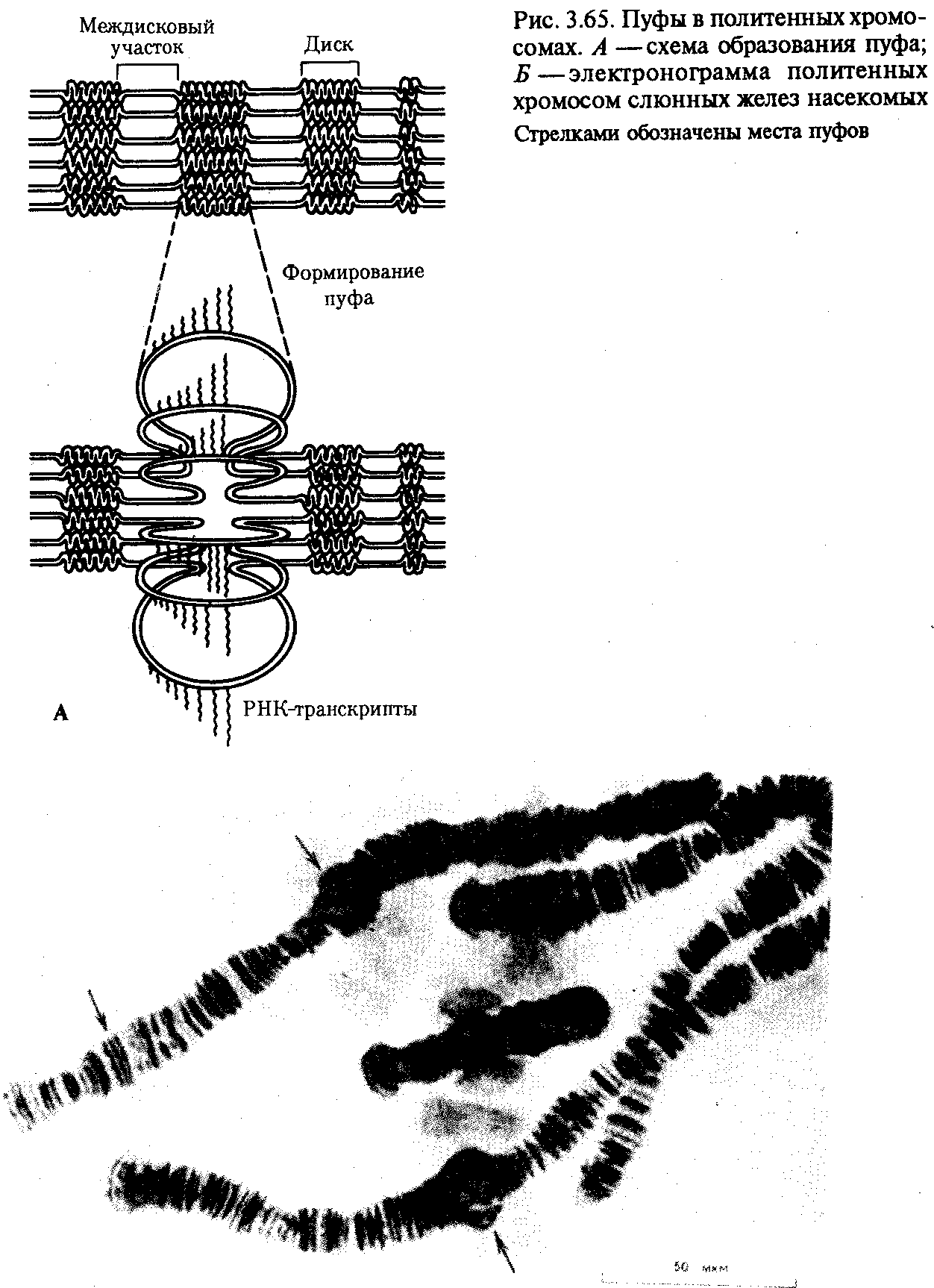 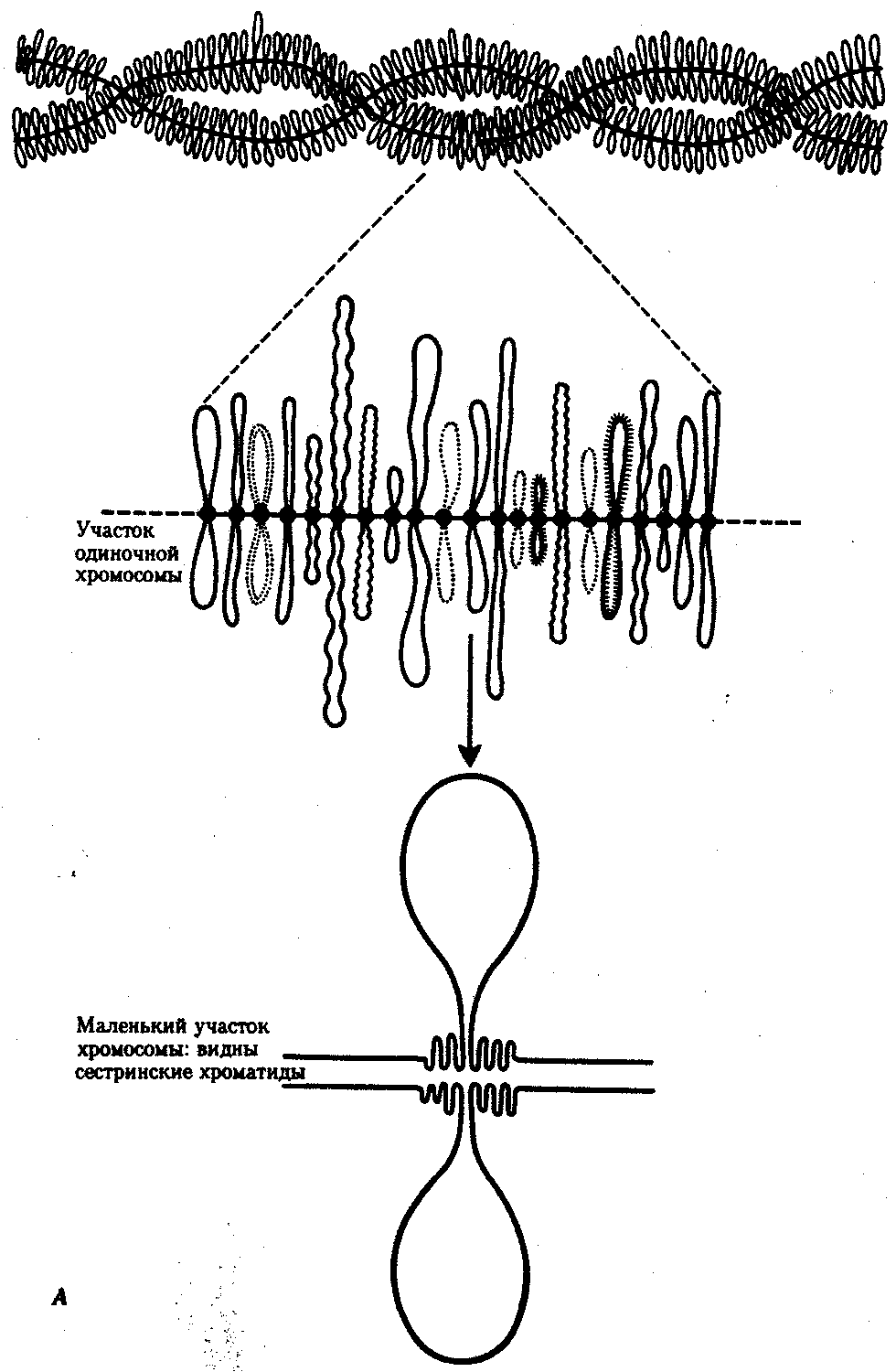 Рис. 3.66. Хромосомы типа ламповых щеток. А— схема строения; Б— хромосомы из овоцита амфибий  Рис. 3.66. Продолжение 3.5.5. Биологическое значение хромосомного уровня организации наследственного материалаВозникновение хромосомной организации наследственного материала в эукариотической клетке тесно связано с большим его объемом по сравнению с прокариотической клеткой. Распределение основной массы генетического материала в ограниченном количестве ядерных структур — хромосом — обеспечивает упорядоченность его пространственной организации по группам сцепления. Относительная самостоятельность хромосом в процессах репликации ДНК и распределения ее молекул между дочерними клетками позволяет закономерно воспроизводить и передавать обширную информацию в ряду клеточных поколений, сохраняя постоянство ее организации. Наконец, распределение генов по группам сцепления допускает возможность рекомбинации генетического материала гомологичных и негомологичных хромосом при мейозе и оплодотворении (см. разд. 3.6.2.3). Эффективность рекомбинации наследственного материала, возрастающая у эукариот благодаря его хромосомной организации, существенно увеличивает степень комбинативной изменчивости у данных организмов. Это является важным эволюционным фактором, обеспечивающим разнообразный исходный материал для естественного отбора. | ||||||||||||||||||||||||||||||