биология ярыгин. Книга 1 Издание пятое, исправленное и дополненное
 Скачать 7.35 Mb. Скачать 7.35 Mb.
|

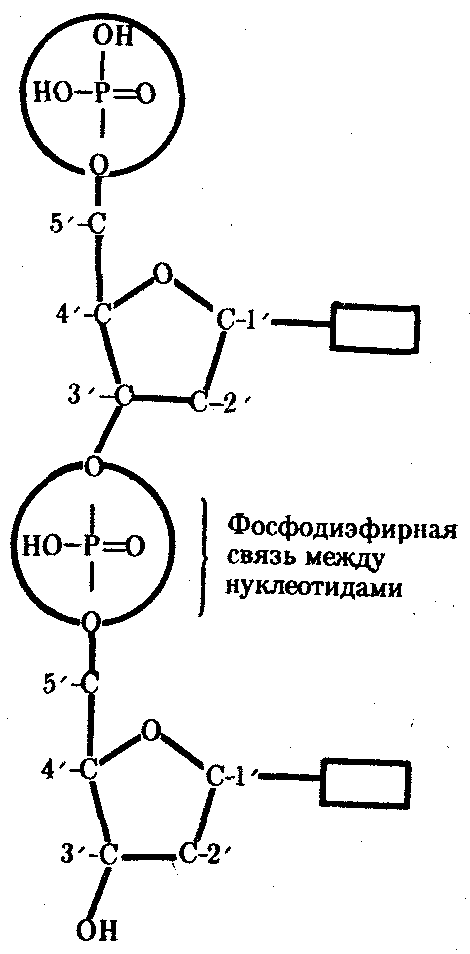
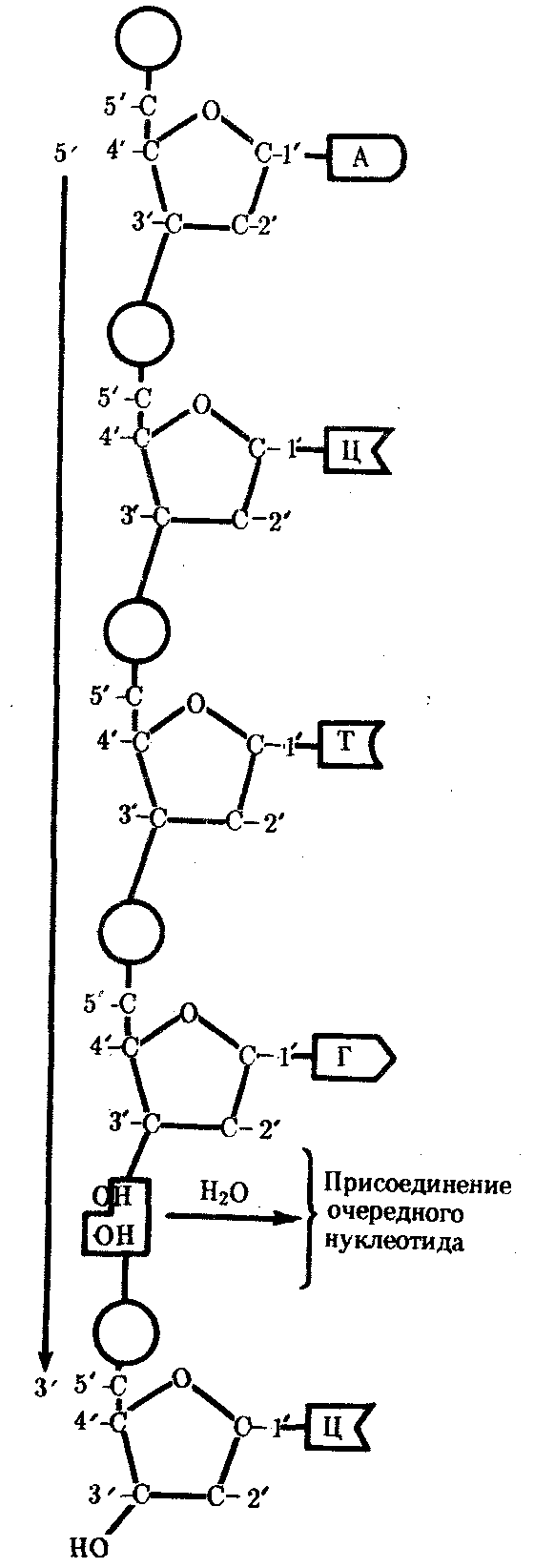
3.3. ОБЩИЕ СВОЙСТВА ГЕНЕТИЧЕСКОГО МАТЕРИАЛА И УРОВНИ ОРГАНИЗАЦИИ ГЕНЕТИЧЕСКОГО АППАРАТАНа основании приведенных выше определений наследственности и изменчивости можно предположить, каким требованиям должен отвечать материальный субстрат этих двух свойств жизни. Во-первых, генетический материал должен обладать способностью к самовоспроизведению, чтобы в. процессе размножения передавать наследственную информацию, на основе которой будет осуществляться формирование нового поколения. Во-вторых, для обеспечения устойчивости характеристик в ряду поколений наследственный материал должен сохранять постоянной свою организацию. В-третьих, материал наследственности и изменчивости должен обладать способностью приобретать изменения и воспроизводить их, обеспечивая возможность исторического развития живой материи в меняющихся условиях. Только в случае соответствия указанным требованиям материальный субстрат наследственности и изменчивости может обеспечить длительность и непрерывность существования живой природы и ее эволюцию. Современные представления о природе генетического аппарата позволяют выделить три уровня его организации: генный, хромосомный и геномный. На каждом из них проявляются основные свойства материала наследственности и изменчивости и определенные закономерности его передачи и функционирования. 3.4. ГЕННЫЙ УРОВЕНЬ ОРГАНИЗАЦИИ ГЕНЕТИЧЕСКОГО АППАРАТАЭлементарной функциональной единицей генетического аппарата, определяющей возможность развития отдельного признака клетки или организма данного вида, является ген (наследственный задаток, по Г. Менделю). Передачей генов в ряду поколений клеток или организмов достигается материальная преемственность — наследование потомками признаков родителей. Под признаком понимают единицу морфологической, физиологической, биохимической, иммунологической, клинической и любой другой дискретности организмов (клеток), т.е. отдельное качество или свойство, по которому они отличаются друг от друга. Большинство перечисленных выше особенностей организмов или клеток относится к категории сложных признаков, формирование которых требует синтеза многих веществ, в первую очередь белков со специфическими свойствами — ферментов, иммунопротеинов, структурных, сократительных, транспортных и других белков. Свойства белковой молекулы определяются аминокислотной последовательностью ее полипептидной цепи, которая прямо задается последовательностью нуклеотидов в ДНК соответствующего гена и является элементарным, или простым, признаком. Основные свойства гена как функциональной единицы генетического аппарата определяются его химической организацией, 3.4.1. Химическая организация генаИсследования, направленные на выяснение химической природы наследственного материала, неопровержимо доказали, что материальным субстратом наследственности и изменчивости являются нуклеиновые кислоты, которые были обнаружены Ф. Мишером (1868) в ядрах клеток гноя. Нуклеиновые кислоты являются макромолекулами, т.е. отличаются большой молекулярной массой. Это полимеры, состоящие из мономеров — нуклеотидов, включающих три компонента: сахар (пентозу), фосфат и азотистое основание (пурин или пиримидин). К первому атому углерода в молекуле пентозы С-1' присоединяется азотистое основание (аденин, гуанин, цитозин, тимин или урацил), а к пятому атому углерода С-5' с помощью эфирной связи — фосфат; у третьего атома углерода С-3' всегда имеется гидроксильная группа — ОН (рис. 3.1). Соединение нуклеотидов в макромолекулу нуклеиновой кислоты происходит путем взаимодействия фосфата одного нуклеотида с гидроксилом другого так, что между ними устанавливается фосфодиэфирная связь (рис. 3.2). В результате образуется полинуклеотидная цепь. Остов цепи состоит из чередующихся молекул фосфата и сахара. К молекулам пентозы в положении С-1' присоединено одно из перечисленных выше азотистых оснований (рис. 3.3). 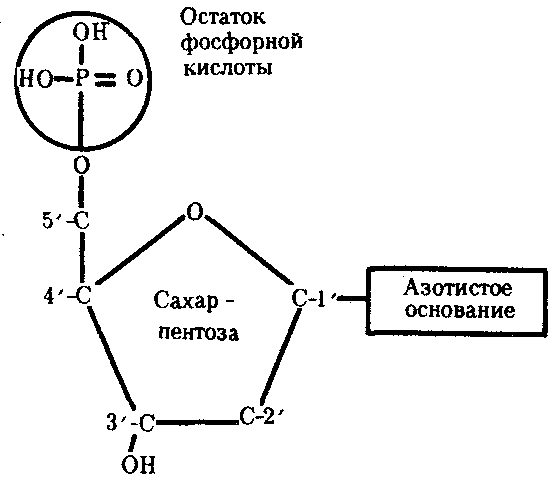 Рис. 3.1. Схема строения нуклеотида Объяснение см. в тексте; обозначения компонентов нуклеотида, использованные в этом рисунке, сохраняются во всех последующих схемах нуклеиновых кислот Сборка полинуклеотидной цепи осуществляется при участии фермента полимеразы, который обеспечивает присоединение фосфатной группы следующего нуклеотида к гидроксильной группе, стоящей в положении 3', предыдущего нуклеотида (рис. 3.3). Благодаря отмеченной специфике действия названного фермента наращивание полинуклеотидной цепи происходит только на одном конце: там, где находится свободный гидроксил в положении 3'. Начало цепи всегда несет фосфатную группу в положении 5'. Это позволяет выделить в ней 5' и 3 '-концы. Среди нуклеиновых кислот различают два вида соединений: дезоксирибонуклеиновую (ДНК) и рибонуклеиновую (РНК) кислоты. Изучение состава основных носителей наследственного материала — хромосом — обнаружило, что их наиболее химически устойчивым компонентом является ДНК, которая представляет собой субстрат наследственности и изменчивости.
3.4.1.1. Структура ДНК. Модель Дж. Уотсона и Ф. КрикаДНК состоит из нуклеотидов, в состав которых входят сахар — дезоксирибоза, фосфат и одно из азотистых оснований — пурин (аденин или гуанин) либо пиримидин (тимин или цитозин). Особенностью структурной организации ДНК является то, что ее молекулы включают две полинуклеотидные цепи, связанные между собой определенным образом. В соответствии с трехмерной моделью ДНК, предложенной в 1953 г. американским биофизиком Дж. Уотсоном и английским биофизиком и генетиком Ф. Криком, эти цепи соединяются друг с другом водородными связями между их азотистыми основаниями по принципу комплементарности. Аденин одной цепи соединяется двумя водородными связями с тимином другой цепи, а между гуанином и цитозином разных цепей образуются три водородные связи. Такое соединение азотистых оснований обеспечивает прочную связь двух цепей и сохранение равного расстояния между ними на всем протяжении. 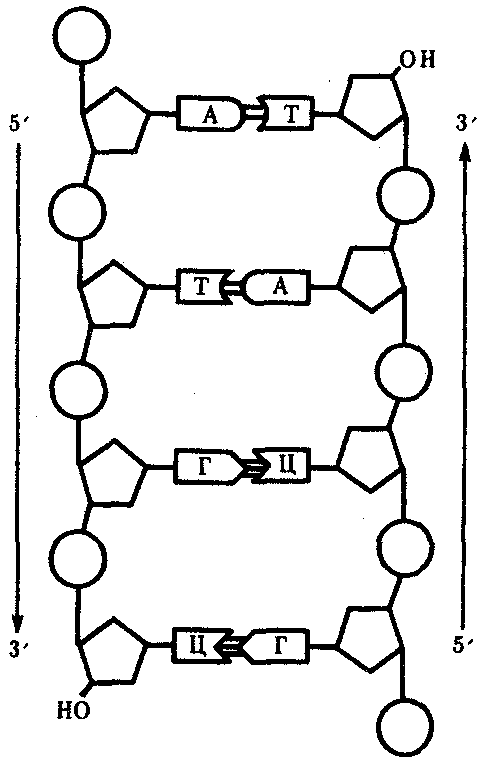 Рис. 3.4. Схема строения молекулы ДНК Стрелками обозначена антилараллельность целей Другой важной особенностью объединения двух полинуклеотидных цепей в молекуле ДНК является их антипараллельность: 5'-конец одной цепи соединяется с 3'-концом другой, и наоборот (рис. 3.4). Данные рентгеноструктурного анализа показали, что молекула ДНК, состоящая из двух цепей, образует спираль, закрученную вокруг собственной оси. Диаметр спирали составляет 2 нм, длина шага — 3, 4 нм. В каждый виток входит 10 пар нуклеотидов. Чаще всего двойные спирали являются правозакрученными — при движении вверх вдоль оси спирали цепи поворачиваются вправо. Большинство молекул ДНК в растворе находится в правозакрученной — В-форме (В-ДНК). Однако встречаются также левозакрученные формы (Z-ДНК). Какое количество этой ДНК присутствует в клетках и каково ее биологическое значение, пока не установлено (рис. 3.5). 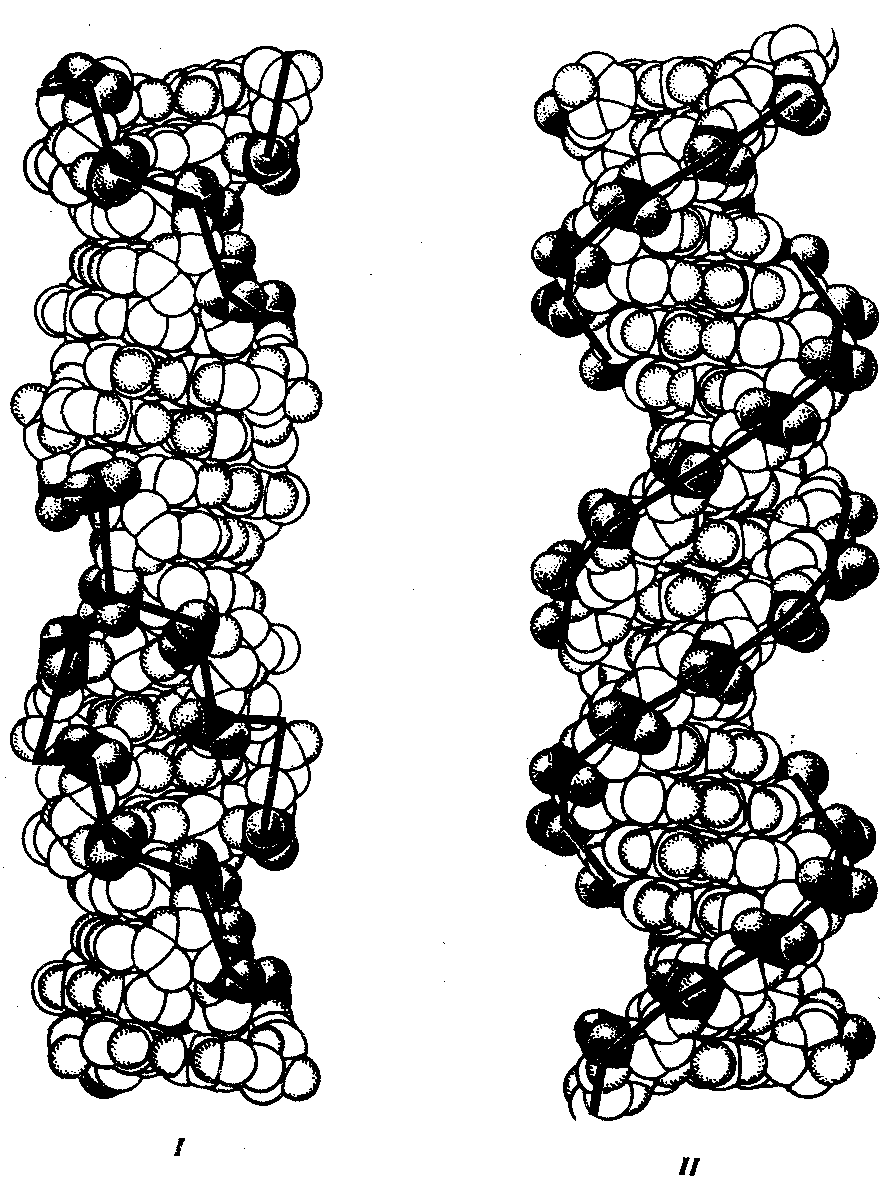 Рис. 3.5. Пространственные модели левоэакрученной Z-формы (I) и правозакрученной В-формы (II) ДНК Таким образом, в структурной организации молекулы ДНК можно выделить первичную структуру —полинуклеотидную цепь, вторичную структуру—две комплементарные друг другу и антипараллельные полинуклеотидные цепи, соединенные водородными связями, и третичную структуру — трехмерную спираль с приведенными выше пространственными характеристиками. 3.4.1.2. Способ записи генетической информации в молекуле ДНК. Биологический код и его свойстваПервично все многообразие жизни обусловливается разнообразием белковых молекул, выполняющих в клетках различные биологические функции. Структура белков определяется набором и порядком расположения аминокислот в их пептидных цепях. Именно эта последовательность аминокислот в пептидах зашифрована в молекулах ДНК с помощью биологического (генетического) кода. Относительная примитивность структуры ДНК, представляющей чередование всего лишь четырех различных нуклеотидов, долгое время мешала исследователям рассматривать это соединение как материальный субстрат наследственности и изменчивости, в котором должна быть зашифрована чрезвычайно разнообразная информация. В 1954 г. Г. Гамовым было высказано предположение, что кодирование информации в молекулах ДНК должно осуществляться сочетаниями нескольких нуклеотидов. В многообразии белков, существующих в природе, было обнаружено около 20 различных аминокислот. Для шифровки такого их числа достаточное количество сочетаний нуклеотидов может обеспечить лишь триплетный код, в котором каждая аминокислота шифруется тремя стоящими рядом нуклеотидами. В этом случае из четырех нуклеотидов образуется 43 = 64 триплета. Код, состоящий из двух нуклеотидов, дал бы возможность зашифровать только 42 = 16 различных аминокислот. Полная расшифовка генетического кода проведена в 60-х гг. нашего столетия. Из 64 возможных триплетов ДНК 61 кодирует различные аминокислоты; оставшиеся 3 получили название бессмысленных, или «нонсенс-триплетов». Они не шифруют аминокислот и выполняют функцию знаков препинания при считывании наследственной информации. К ним относятся АТТ, АЦТ, АТЦ. Обращает на себя внимание явная избыточность кода, проявляющаяся в том, что многие аминокислоты шифруются несколькими триплетами (рис. 3.6). Это свойство триплетного кода, названное вырожденностью, имеет очень важное значение, так как возникновение в структуре молекулы ДНК изменений по типу замены одного нукле-отида в полинуклеотидной цепи может не изменить смысла триплета. Возникшее таким образом новое сочетание из трех нуклеотидов кодирует ту же самую аминокислоту. В процессе изучения свойств генетического кода была обнаружена его специфичность. Каждый триплет способен кодировать только одну определенную аминокислоту. Интересным фактом является полное соответствие кода у различных видов живых организмов. Такая универсальность генетического кода свидетельствует о единстве происхождения всего многообразия живых форм на Земле в процессе биологической эволюции. Незначительные отличия генетического кода обнаружены в ДНК митохондрий некоторых видов. Это не противоречит в целом положению об универсальности кода, но свидетельствует в пользу определенной дивергентности в его эволюции на ранних этапах существования жизни. Расшифровка кода в ДНК митохондрий различных видов показала, что во всех случаях в митохондриальных ДНК отмечается общая особенность: триплет АЦТ читается как АЦЦ, и поэтому из нонсенс-триплета превращается в шифр аминокислоты триптофана. 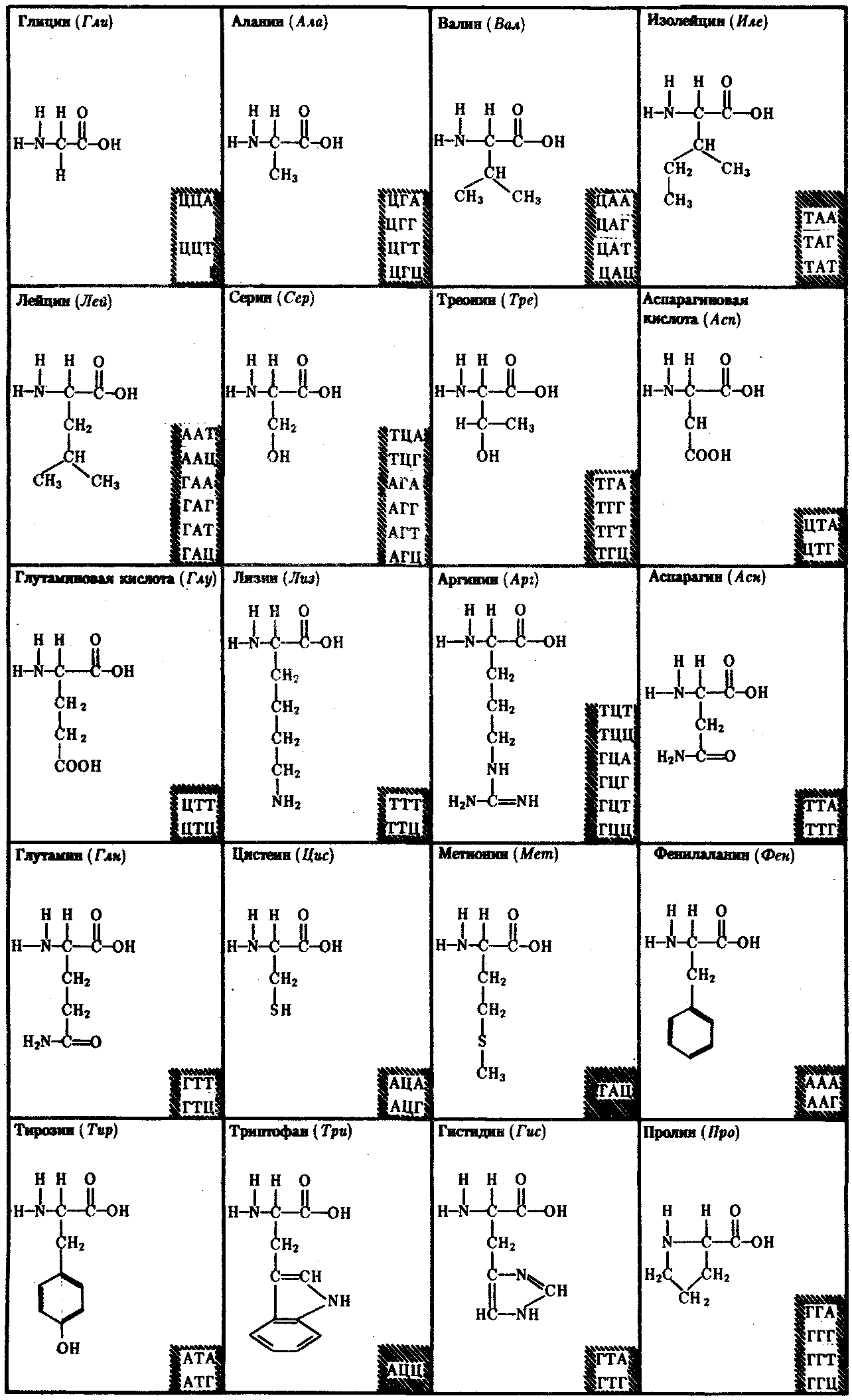 Рис. 3.6. Аминокислоты и кодирующие их триплеты ДНК Другие особенности являются специфичными для различных видов организмов. У дрожжей триплет ГАТ и, возможно, все семейство ГА кодирует вместо аминокислоты лейцина треонин. У млекопитающих триплет ТАГ имеет то же значение, что и ТАЦ, и кодирует аминокислоту метионин вместо изолейцина. Триплеты ТЦГ и ТЦЦ в ДНК митохондрий некоторых видов не кодируют аминокислот, являясь нонсенс-триплетами. Наряду с триплетностью, вырожденностью, специфичностью и универсальностью важнейшими характеристиками генетического кода являются его непрерывность и неперекрываемость кодонов при считывании. Это означает, что последовательность нуклеотидов считывается триплет за триплетом без пропусков, при этом соседние триплеты не перекрывают друг друга, т.е. каждый отдельный нуклеотид входит в состав только одного триплета при заданной рамке считывания (рис. 3.7). Доказательством неперекрываемости генетического кода является замена только одной аминокислоты в пептиде при замене одного нуклеотида в ДНК. В случае включения нуклеотида в несколько перекрывающихся триплетов его замена влекла бы за собой замену 2—3 аминокислот в пептидной цепи. 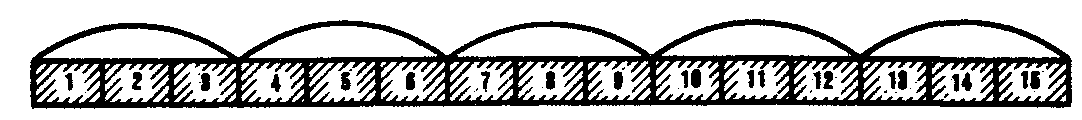 Рис. 3.7. Непрерывность и непререкаемость генетического кода при считывании наследственной информации Цифрами обозначены нуклеотиды |