биология ярыгин. Книга 1 Издание пятое, исправленное и дополненное
 Скачать 7.35 Mb. Скачать 7.35 Mb.
|

РАЗДЕЛ IIКЛЕТОЧНЫЙ И МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЙ УРОВНИ ОРГАНИЗАЦИИ ЖИЗНИ — ОСНОВА ЖИЗНЕДЕЯТЕЛЬНОСТИ ОРГАНИЗМОВГЛАВА 2КЛЕТКА — ЭЛЕМЕНТАРНАЯ ЕДИНИЦА ЖИВОГОКлетка представляет собой обособленную, наименьшую по размерам структуру, которой присуща вся совокупность свойств жизни и которая может в подходящих условиях окружающей среды поддерживать эти свойства в самой себе, а также передавать их в ряду поколений. Клетка, таким образом, несет полную характеристику жизни. Вне клетки не существует настоящей жизнедеятельности. Поэтому в природе планеты ей принадлежит роль элементарной структурной, функциональной и генетической единицы. Это означает, что клетка составляет основу строения, жизнедеятельности и развития всех живых форм — одноклеточных, многоклеточных и даже неклеточных. Благодаря заложенным в ней механизмам клетка обеспечивает обмен веществ, использование биологической информации, размножение, свойства наследственности и изменчивости, обусловливая тем самым присущие органическому миру качества единства и разнообразия. Занимая в мире живых существ положение элементарной единицы, клетка отличается сложным строением. При этом определенные черты обнаруживаются во всех без исключения клетках, характеризуя наиболее важные стороны клеточной организации как таковой. 2.1. КЛЕТОЧНАЯ ТЕОРИЯКлеточная теория сформулирована немецким исследователем, зоологом Т. Шванном (1839). Поскольку при создании этой теории Шванн широко пользовался работами ботаника М. Шлейдена, последнего по праву считают соавтором клеточной теории. Исходя из предположения о схожести (гомологичности) растительных и животных клеток, доказываемой одинаковым механизмом их возникновения, Шванн обобщил многочисленные данные в виде теории, согласно которой клетки являются структурной и функциональной основой живых существ. В конце XIX столетия немецкий патолог Р. Вирхов на основе новых фактов пересмотрел клеточную теорию. Ему принадлежит вывод о том, что клетка может возникнуть лишь из предсуществующей клетки. Им также создана вызвавшая критику концепция «клеточного государства», согласно которой многоклеточный организм состоит из относительно самостоятельных единиц (клеток), поставленных в своей жизнедеятельности в тесную зависимость друг от друга. Клеточная теория в современном виде включает три главных положения. Первое положение соотносит клетку с живой природой планеты в целом. Оно утверждает, что жизнь, какие бы сложные или простые (например, вирусы) формы она ни принимала, в ее структурном, функциональном и генетическом отношении обеспечивается в конечном итоге только клеткой. Выдающаяся роль клетки как первоисточника жизни обусловливается тем, что именно она является биологической единицей, с помощью которой происходит извлечение из внешней среды, превращение и использование организмами энергии и веществ. Непосредственно в клетке сохраняется и используется биологическая информация. Второе положение указывает, что в настоящих условиях единственным способом возникновения новых клеток является деление предсуществующих клеток. В обосновании клеточной природы жизни на Земле тезису о единообразии путей возникновения клеток принадлежит особая роль. Именно этот тезис был использован М. Шлейденом и Т. Шванном для обоснования представления о гомологии разных типов клеток1. Современная биология расширила круг доказательств этому. Независимо от индивидуальных структурно-функциональных особенностей все клетки одинаковым образом: а) хранят биологическую информацию, б) редуплицируют генетический материал с целью его передачи в ряду поколений, в) используют информацию для осуществления своих функций на основе синтеза белка, г) хранят и переносят энергию, д) превращают энергию в работу, е) регулируют обмен веществ. Третье положение клеточной теории соотносит клетку с многоклеточными организмами, для которых характерен принцип целостности и системной организации. Для системы свойственно наличие новых качеств благодаря взаимному влиянию и взаимодействию единиц, составляющих эту систему. Структурно-функциональными единицами многоклеточных существ являются клетки. Вместе с тем многоклеточный организм характеризуется рядом особых свойств, которые нельзя свести к свойствам и качествам отдельных клеток. В третьем положении клеточной теории мы встречаемся с проблемой соотношения части и целого. Системный подход как научное направление используется в биологических исследованиях с начала прошлого столетия. Системный характер организации и функционирования свойствен не только организму, но и другим главным биологическим образованиям — геному, клетке, популяции, биогеоценозу, биосфере. 2.2. ТИПЫ КЛЕТОЧНОЙ ОРГАНИЗАЦИИВ природе существует значительное разнообразие клеток, различающихся по размерам, форме, химическим особенностям. Число же главных типов клеточной организации ограничено двумя. Выделяют прокариотический и эукариотический типы с подразделением второго на подтип, характерный для простейших организмов, и подтип, характерный для многоклеточных. Клеткам прокариоттеского типа (рис. 2.1) свойственны малые размеры (не более 0,5—3,0 мкм в диаметре или по длине), отсутствие обособленного ядра, так что генетический материал в виде ДНК не отграничен от цитоплазмы оболочкой. В клетке отсутствует развитая система мембран. Генетический аппарат представлен ДНК единственной кольцевой хромосомы, которая лишена основных белков — гистонов (гистоны являются белками клеточных ядер). Благодаря значительному количеству диаминокислот аргинина и лизина гистоны имеют щелочной характер. Различия прокариотических и эукариотических клеток по наличию гистонов указывают на разные механизмы регуляции функции генетического материала. В прокариотических клетках отсутствует клеточный центр. Не типичны внутриклеточные перемещения цитоплазмы и амебоидное движение. Время, необходимое для образования двух дочерних клеток из материнской (время генерации), сравнительно мало и исчисляется десятками минут. К прокариотическому типу клеток относятся бактерии и синезеленые водоросли. 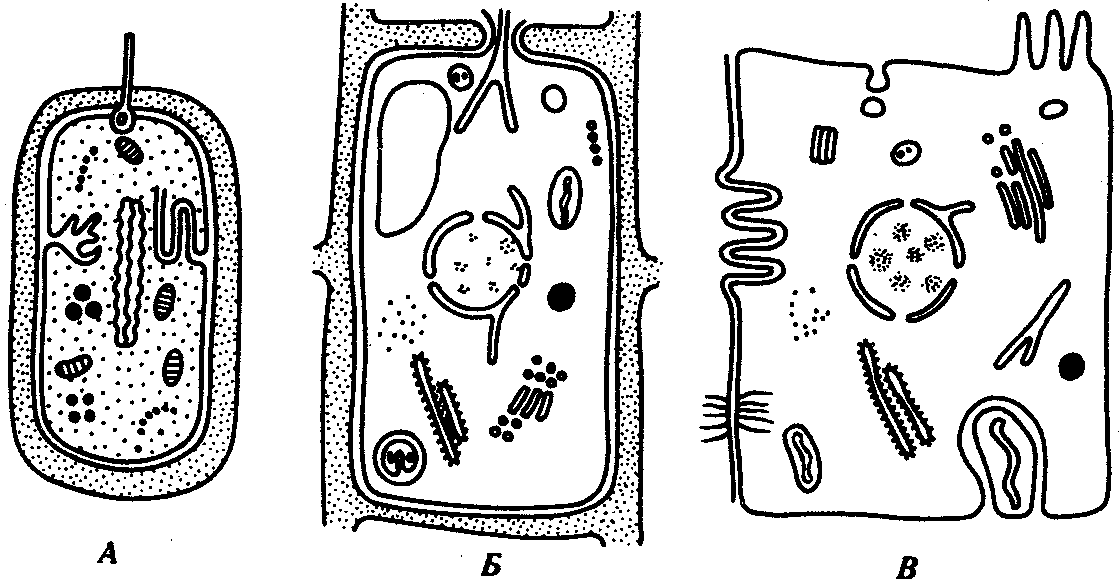 Рис. 2.1. Типичные черты структурной организации клеток. А -Б — эукариотическая растительная; В — животная 2.3. СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ЭУКАРИОТИЧЕСКОЙ КЛЕТКИЭукариотический тип клеточной организации представлен двумя подтипами. Особенностью организмов простейших (рис. 2.2) является то, что они (исключая колониальные формы) соответствуют в структурном отношении уровню одной клетки, а в физиологическом — полноценной особи. В связи с этим одной из черт клеток части простейших является наличие в цитоплазме миниатюрных образований, выполняющих на клеточном уровне функции жизненно важных органов многоклеточного организма. Таковы (например, у инфузорий) цитостом, цитофарингс и порошица, аналогичные пищеварительной системе, и сократительные вакуоли, аналогичные выделительной системе. В традиционном изложении клетку растительного или животного организма описывают как объект, отграниченный оболочкой, в котором выделяют ядро и цитоплазму. В ядре наряду с оболочкой и ядерным соком обнаруживаются ядрышко и хроматин. Цитоплазма представлена ее основным веществом (матриксом, гиалоплазмой), в котором распределены включения и органеллы. 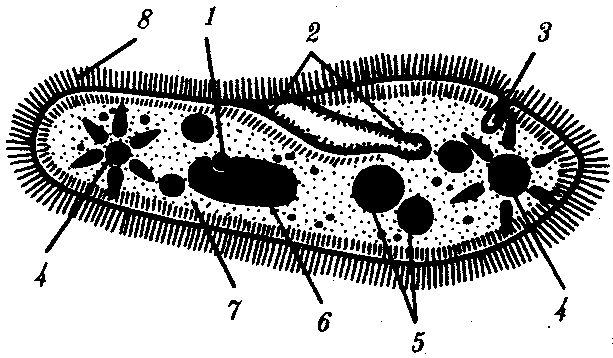 Рис. 2.2. Структурная организация одноклеточного организма (инфузория): 1—генеративное ядро, 2—цигостом с цитофарингсом, 3—порошица, 4—сократительные вакуоли, 5—пищеварительные вакуоли, 6—вегетативное ядро, 7—гиалоплазма, 8— реснички 2.3.1. Принцип компартментации.Биологическая мембранаВысокая упорядоченность внутреннего содержимого эукариотиче-ской клетки достигается путем компартментации ее объема — подразделения на «ячейки», отличающиеся деталями химического (ферментного) состава. Компартментация (рис. 2.3) способствует пространственному разделению веществ и процессов в клетке. Отдельный компартмент представлен органеллой (лизосома) или ее частью (пространство, отграниченное внутренней мембраной митохондрии). 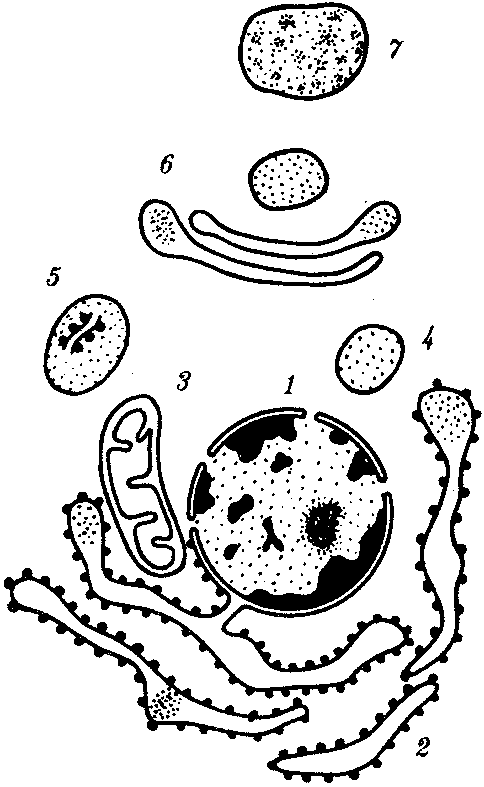 Рис. 2.3. Компартментация объема клетки с помощью мембран: 1—ядро, 2—шероховатая цитоплазматическая есть, 3—митохондрия, 4—транспортный цитоплазматический пузырек, 5—лизосома, 6—пластинчатый комплекс, 7 — гранула секрета 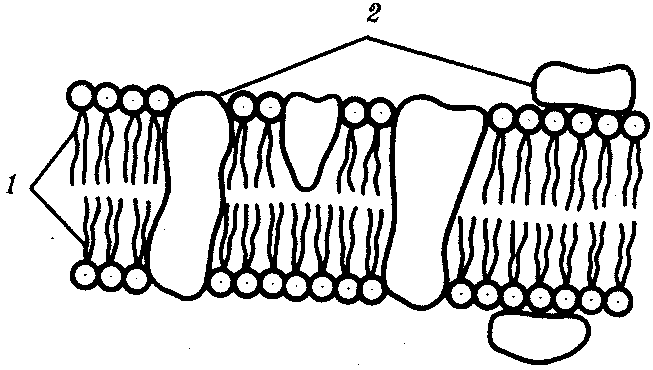 Рис. 2.4. Молекулярная организация биологической мембраны: 1 — бимолекулярный слой липидов, 2 — белки Предложено несколько схем взаимоотношения в мембране основных химических компонентов — белков и липидов, а также веществ, размещаемых на мембранной поверхности. В настоящее время принята точка зрения, согласно которой мембрана составлена из бимолекулярного слоя липидов. Гидрофобные участки их молекул повернуты друг к другу, а гидрофильные — находятся на поверхности слоя. Разнообразные белковые молекулы встроены в этот слой или размещены на его поверхностях (рис. 2.4). Благодаря компартментации клеточного объема в эукариотической клетке наблюдается разделение функций между разными структурами. Одновременно различные структуры закономерно взаимодействуют друг с другом. 2.3.2. Строение типичной клетки многоклеточного организмаКлетки многоклеточных организмов, как животных, так и растительных, обособлены от своего окружения оболочкой. Клеточная оболочка, или плазмалемма, животных клеток образована мембраной, покрытой снаружи слоем гликокаликса толщиной 10—20 нм. Основными составляющими гликокаликса служат комплексы полисахаридов с белками (гликопротеины) и жирами (гликолипиды). Изнутри к мембране примыкает кортикальный (корковый) слой цитоплазмы толщиной 0,1—0,5 мкм, в котором не встречаются рибосомы и пузырьки, но в значительном количестве находятся микротрубочки и микрофиламенты, имеющие в своем составе сократимые белки. Плазмалемма выполняет отграничивающую, барьерную, транспортную и рецепторную функции. Благодаря свойству избирательной проницаемости она регулирует химический состав внутренней среды клетки. В плазмалемме размещены молекулы рецепторов, которые избирательно распознают определенные биологически активные вещества (гормоны). В удержании (заякоривании) этих веществ на клеточной поверхности участвуют белки кортикального слоя. Наличие в оболочке рецепторов дает клеткам возможность воспринимать сигналы извне, чтобы целесообразно реагировать на изменения в окружающей их среде или состоянии организма. В пластах и слоях соседние клетки удерживаются благодаря наличию разного вида контактов, которые представлены участками плазмалеммы, имеющими особое строение. В клетке выделяют ядро и цитоплазму. Клеточное ядро состоит из оболочки, ядерного сока, ядрышка и хроматина. Функциональная роль ядерной оболочки заключается в обособлении генетического материала (хромосом) эукариотической клетки от цитоплазмы с присущими ей многочисленными метаболическими реакциями, а также регуляции двусторонних взаимодействий ядра и цитоплазмы. Ядерная оболочка состоит из двух мембран, разделенных околоядерным (перинуклеарным) пространством. Последнее может сообщаться с канальцами цитоплазматической сети. Ядерная оболочка пронизана порожу диаметром 80—90 нм. Область поры или поровый комплекс с диаметром около 120 нм имеет определенное строение, что указывает на сложный механизм регуляции ядерно-цитоплазматических перемещений веществ и структур. Количество пор зависит от функционального состояния клетки. Чем выше синтетическая активность в клетке, тем больше их число. Подсчитано, что у низших позвоночных животных в эритробластах, где интенсивно образуется и накапливается гемоглобин, на 1 мкм2 ядерной оболочки приходится около 30 пор. В зрелых эритроцитах названных животных, сохраняющих ядра, на 1 мк»г оболочки остается до пяти пор, т.е. в 6 раз меньше. В области перового комплекса начинается так называемая плотная пластинка — белковый слой, подстилающий на всем протяжении внутреннюю мембрану ядерной оболочки. Эта структура выполняет прежде всего опорную функцию, так как при ее наличии форма ядра сохраняется даже в случае разрушения обеих мембран ядерной оболочки. Предполагают также, что закономерная связь с веществом плотной пластинки способствует упорядоченному расположению хромосом в интерфазном ядре. Основу ядерного сока, или матрикса, составляют белки. Ядерный сок образует внутреннюю среду ядра, в связи с чем он играет важную роль в обеспечении нормального функционирования генетического материала. В составе ядерного сока присутствуют нитчатые, или фибриллярные, белки, с которыми связано выполнение опорной функции: в матриксе находятся также первичные продукты транскрипции генетической информации — гетероядерные РНК (гя-РНК), которые здесь же подвергаются процессингу, превращаясь в м-РНК (см. 3.4.3.2). Ядрышко представляет собой структуру, в которой происходит образование и созревание рибосомальных РНК (рРНК). Гены рРНК занимают определенные участки (в зависимости от вида животного) одной или нескольких хромосом (у человека 13—15 и 21—22 пары) — ядрышковые организаторы, в области которых и образуются ядрышки. Такие участки в метафазных хромосомах выглядят как сужения и называются вторичными перетяжками. С помощью электронного микроскопа в ядрышке выявляют нитчатый и зернистый компоненты. Нитчатый (фибриллярный) компонент представлен комплексами белка и гигантских молекул РНК-предшественниц, из которых затем образуются более мелкие молекулы зрелых рРНК. В процессе созревания фибриллы преобразуются в рибонуклеопротеиновые зерна (гранулы), которыми представлен зернистый компонент. Хроматиновые структуры в виде глыбок, рассеянных в нуклеоплазме, являются интерфазной формой существования хромосом клетки (см. 3.5.2.2). В цитоплазме различают основное вещество (матрикс, гиалоплазма), включения и органеллы. Основное вещество цитоплазмы заполняет пространство между плазмалеммой, ядерной оболочкой и другими внутриклеточными структурами. Обычный электронный микроскоп не выявляет в нем какой-либо внутренней организации. Белковый состав гиалоплазмы разнообразен. Важнейшие из белков представлены ферментами гаиколиза, обмена Сахаров, азотистых оснований, аминокислот и липидов. Ряд белков гиалоплазмы служит субъединицами, из которых происходит сборка таких структур, как микротрубочки. Основное вещество цитоплазмы образует истинную внутреннюю среду клетки, которая объединяет все внутриклеточные структуры и обеспечивает взаимодействие их друг с другом. Выполнение матриксом объединяющей, а также каркасной функции может быть связано с выявляемой с помощью сверхмощного электронного микроскопа микротрабекулярной сети, образованной тонкими фибриллами толщиной 2—3 нм и пронизывающей всю цитоплазму. Через гиалоплазму осуществляется значительный объем внутриклеточных перемещений веществ и структур. Основное вещество цитоплазмы следует рассматривать так же, как сложную коллоидную систему, способную переходить из золеобразного (жидкого) состояния в гелеобразное. В процессе таких переходов совершается работа. О функциональном значении таких переходов см. разд. 2.3.8. Включениями (рис. 2.5) называют относительно непостоянные компоненты цитоплазмы, которые служат запасными питательными веществами (жир, гликоген), продуктами, подлежащими выведению из клетки (гранулы секрета), балластными веществами (некоторые пигменты). Органеллы — это постоянные структуры цитоплазмы, выполняющие в клетке жизненно важные функции. Выделяют органеллы общего значения и специальные. Последние в значительном количестве присутствуют в клетках, специализированных к выполнению определенной функции, но в незначительном количестве могут встречаться и в других типах клеток. К ним относят, например, микроворсинки всасывающей поверхности эпителиальной клетки кишечника, реснички эпителия трахеи и бронхов, синаптические пузырьки, транспортирующие вещества — переносчики нервного возбуждения с одной нервной клетки на другую или клетку рабочего органа, миофибриллы, от которых зависит сокращение мышцы. Детальное рассмотрение специальных органелл входит в задачу курса гистологии. К органеллам общего значения относят элементы канальцевой и вакуолярной системы в виде шероховатой и гладкой цитоплазматической сети, пластинчатый комплекс, митохондрии, рибосомы и полисомы, лизосомы, пероксисомы, микрофибриллы и микротрубочки, центриоли клеточного центра. В растительных клетках выделяют также хлоропласта, в которых происходит фотосинтез. Канальцевая и вакуолярная системы образованы сообщающимися или отдельными трубчатыми или уплощенными (цистерна) полостями, ограниченными мембранами и распространяющимися по всей цитоплазме клетки. Нередко цистерны имеют пузыревидные расширения. В названной системе выделяют шероховатую и гладкую цитоплазматическую сети (см. рис. 2.3). Особенность строения шероховатой сети состоит в прикреплении к ее мембранам полисом. В силу этого она выполняет функцию синтеза определенной категории белков, преимущественно удаляемых из клетки, например секретируемых клетками желез. В области шероховатой сети происходит образование белков и липидов цитоплазматических мембран, а также их сборка. Плотно упакованные в слоистую структуру цистерны шероховатой сети являются участками наиболее активного белкового синтеза и называются эргастоплазмой. Мембраны гладкой цитоплазматической сети лишены полисом. Функционально эта сеть связана с обменом углеводов, жиров и других веществ небелковой природы, например стероидных гормонов (в половых железах, корковом слое надпочечников). По канальцам и цистернам происходит перемещение веществ, в частности секретируемого железистой клеткой материала, от места синтеза в зону упаковки в гранулы. В участках печеночных клеток, богатых структурами гладкой сети, разрушаются и обезвреживаются вредные токсические вещества, некоторые лекарства (барбитураты). В пузырьках и канальцах гладкой сети поперечно-полосатой мускулатуры сохраняются (депонируются) ионы кальция, играющие важную роль в процессе сокращения.  Рис. 2.5. Цитопдазматические включения эукариотической клетки. А — липофусцин; Б — липидная капля; В — гранулы секрета Рибосома — это округлая рибонуклеопротеиновая частица диаметром 20—30 нм. Она состоит из малой и большой субъединиц, объединение которых происходит в присутствии матричной (информационной) РНК (мРНК). Одна молекула мРНК обычно объединяет несколько рибосом наподобие нитки бус. Такую структуру называют полисомой. Полисомы свободно располагаются в основном веществе цитоплазмы или прикреплены к мембранам шероховатой цитоплазматической сети. В обоих случаях они служат местом активного синтеза белка. Сравнение соотношения количества свободных и прикрепленных к мембранам полисом в эмбриональных недифференцированных и опухолевых клетках, с одной стороны, и в специализированных клетках взрослого организма —с другой, привело к заключению, что на полисомах гиалоплазмы образуются белки для собственных нужд (для «домашнего» пользования) данной клетки, тогда как на полисомах гранулярной сети синтезируются белки, выводимые из клетки и используемые на нужды организма (например, пищеварительные ферменты, белки грудного молока). Пластинчатый комплекс Голъджи образован совокупностью диктиосом числом от нескольких десятков (обычно около 20) до нескольких сотен и даже тысяч на клетку. Диктиосома (рис. 2.6, А) представлена стопкой из 3—12 уплощенных дискообразных цистерн, от краев которых отшнуровываются пузырьки (везикулы). Ограниченные определенным участком (локальные) расширения цистерн дают более крупные пузырьки (вакуоли). В дифференцированных клетках позвоночных животных и человека диктиосомы обычно собраны в околоядерной зоне цитоплазмы. В пластинчатом комплексе образуются секреторные пузырьки или вакуоли, содержимое которых составляют белки и другие соединения, подлежащие выводу из клетки. При этом предшественник секрета (просекрет), поступающий.в диктиосому из зоны синтеза, подвергается в ней некоторым химическим преобразованиям. Он также обособляется (сегрегируется) в виде «порций», которые здесь же одеваются мембранной оболочкой. В пластинчатом комплексе образуются лизосомы. В диктиосомах синтезируются полисахариды, а также их комплексы с белками (гликопротеины) и жирами (гликолипиды), которые затем можно обнаружить в гликокаликсе клеточной оболочки. Митохондрии (рис. 2.6, Б) — это структуры округлой или палочко-видной, нередко ветвящейся формы толщиной 0,5 мкм и длиной обычно до 5—10 мкм. В большинстве животных клеток количество митохондрий колеблется от 150 до 1500, однако в женских половых клетках их число достигает нескольких сотен тысяч. В сперматозоидах нередко присутствует одна гигантская митохондрия, спирально закрученная вокруг осевой части жгутика. Одна разветвленная митохондрия обнаружена в клетке такого паразита человека, как трипаносома. Оболочка митохондрий состоит из двух мембран, различающихся по химическому составу, набору ферментов и функциям. Внутренняя мембрана образует впячивания листовидной (кристы) или трубчатой (тубулы) формы. Пространство, ограниченное внутренней мембраной, составляет матрикс органеллы. В нем с помощью электронного микроскопа обнаруживаются зерна диаметром 20—40 нм. Они накапливают ионы кальция и магния, а также полисахариды, например гликоген. 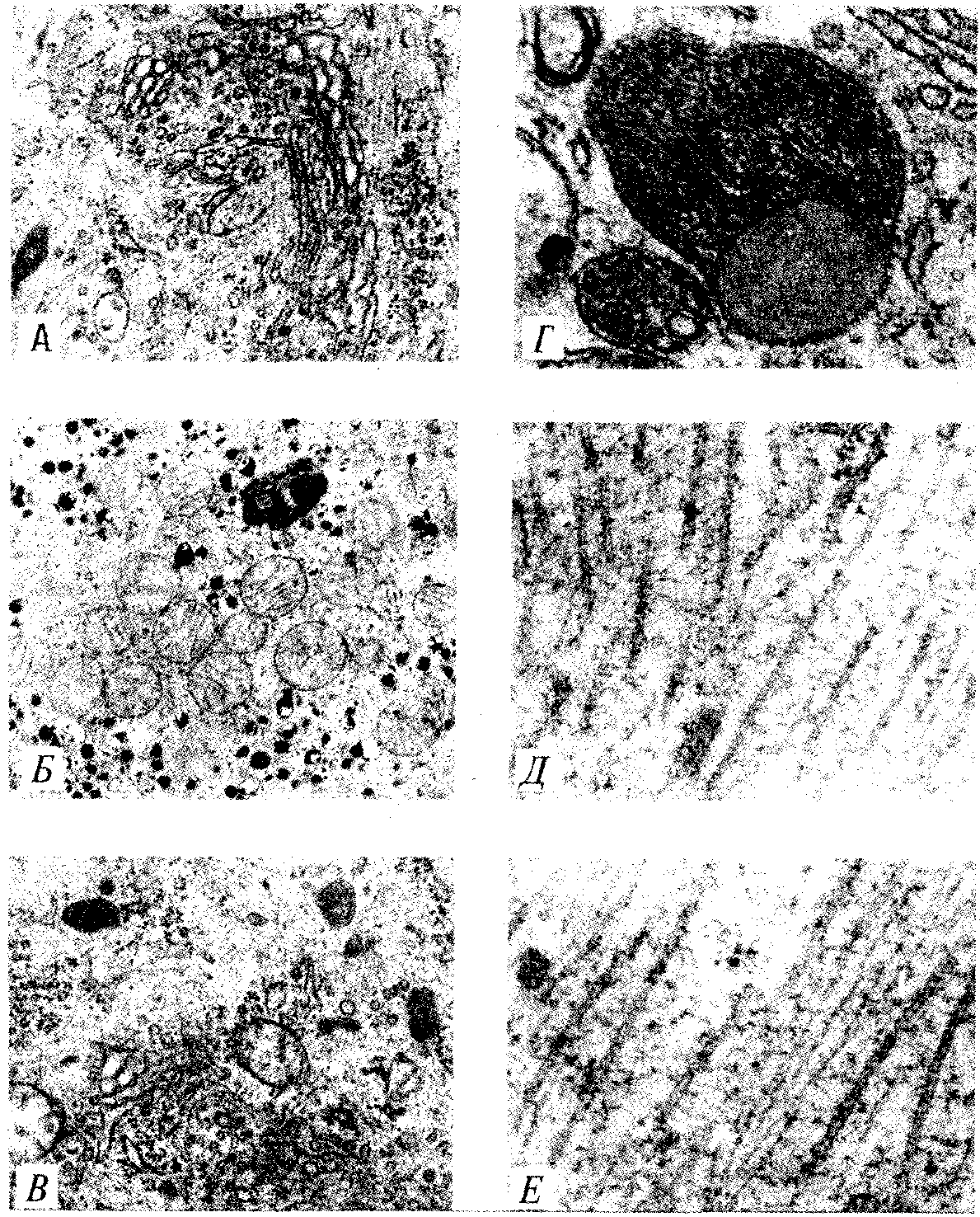 Рис. 2.6. Общеклеточные органеллы Объяснение см. в тексте В матриксе размещен собственный аппарат биосинтеза белка органеллы. Он представлен 2—б копиями кольцевой и лишенной гистонов (как у прокариот) молекулы ДНК, рибосомами, набором транспортных РНК (тРНК), ферментами редупликации ДНК, транскрипции и трансляции наследственной информации. По основным свойствам: размерам и структуре рибосом, организации собственного наследственного материала — этот аппарат сходен с таковым у прокариот и отличается от аппарата биосинтеза белка цитоплазмы эукариотической клетки (чем подтверждается симбиотическая гипотеза происхождения митохондрий; см. § 1.5). Гены собственной ДНК кодируют нуклеотидные последовательности митохондриальных рРНК и тРНК, а также последовательности аминонокислот некоторых белков органеллы, главным образом ее внутренней мембраны. Аминокислотные последовательности (первичная структура) большинства белков митохондрий закодированы в ДНК клеточного ядра и образуются вне органеллы в цитоплазме. Главная функция митохондрий состоит в ферментативном извлечении из определенных химических веществ энергии (путем их окисления) и накоплении энергии в биологически используемой форме (путем синтеза молекул аденозинтрифосфата —АТФ). В целом этот процесс называется окислительным (расформированием. В энергетической функции митохондрий активно участвуют компоненты матрикса и внутренняя мембрана. Именно с этой мембраной связаны цепь переноса электронов (окисление) и АТФ-синтетаза, катализирующая сопряженное с окислением фосфорилирование АДФ в АТФ. Среди побочных функций митохондрий можно назвать участие в синтезе стероидных гормонов и некоторых аминокислот (глутаминовая). Лизосомы (рис. 2.6, В) представляют собой пузырьки диаметром обычно 0,2—0,4 мкм, которые содержат набор ферментов кислых гидролаз, катализирующих при низких значениях рН гидролитическое (в водной среде) расщепление нуклеиновых кислот, белков, жиров, полисахаридов. Их оболочка образована одинарной мембраной, покрытой иногда снаружи волокнистым белковым слоем (на электронограммах «окаймленные» пузырьки). Функция лизосом — внутриклеточное переваривание оазличных химических соединений и структур. Первичными лизосомами (диаметр 100 нм) называют неактивные органеллы, вторичными — органеллы, в которых происходит процесс переваривания. Вторичные лизосомы образуются из первичных. Они подразделяются на гетеролизосомы (фаголизосомы) и аутолизосомы (цитолизосомы). В первых (рис. 2.6, Г) переваривается материал, поступающий в клетку извне путем пиноцитоза и фагоцитоза, во вторых разрушаются собственные структуры клетки, завершившие свою функцию. Вторичные лизосомы, в которых процесс переваривания завершен, называют остаточными тельцами (телолизосомы). В них отсутствуют гидролазы и содержится непереваренный материал. Микротельца составляют сборную группу органелл. Это ограниченные одной мембраной пузырьки диаметром 0,1—1,5 мкм с мелкозернистым матриксом и нередко кристаллоидными или аморфными белковыми включениями. К этой группе относят, в частности, пероксисомы. Они содержат ферменты оксидазы, катализирующие образование пероксида водорода, который, будучи токсичным, разрушается затем под действием фермента пероксидазы. Эти реакции включены в различные метаболические циклы, например в обмен мочевой кислоты в клетках печени и почек. В печеночной клетке число пероксисом достигает 70—100. К органеллам общего значения относят также некоторые постоянные структуры цитоплазмы, лишенные мембран. Микротрубочки (рис. 2.6, Д) — трубчатые образования различной длины с внешним диаметром 24 нм, шириной просвета 15 нм и толщиной стенки около 5 нм. Встречаются в свободном состоянии в цитоплазме клеток или как структурные элементы жгутиков, ресничек, митотического веретена, центриолей. Свободные микротрубочки и микротрубочки ресничек, жгутиков и центриолей имеют разную устойчивость к разрушающим воздействиям, например химическим (колхицин). Микротрубочки строятся из стереотипных субьединиц белковой природы путем их полимеризации. В живой клетке процессы полимеризации протекают одновременно с процессами деполимеризации. Соотношением этих процессов определяется количество микротрубочек. В свободном состоянии микротрубочки выполняют опорную функцию, определяя форму клеток, а также являются факторами направленного перемещения внутриклеточных компонентов. Микрофиламентами (рис. 2.6, Е) называют длинные, тонкие образования, иногда образующие пучки и обнаруживаемые по всей цитоплазме. Существует несколько разных типов микрофиламентов. Актиновые микрофиламенты благодаря присутствию в них сократимых белков (актин) рассматривают в качестве структур, обеспечивающих клеточные формы движения, например амебоидные. Им приписывают также каркасную роль и участие в организации внутриклеточных перемещений органелл и участков гиалоплазмы. По периферии клеток под плазмалеммой, а также в околоядерной зоне обнаруживаются пучки микрофиламентов толщиной 10 нм — промежуточные филстенты. В эпителиальных, нервных, глиальных, мышечных клетках, фибробластах они построены из разных белков. Промежуточные филаменты выполняют, по-видимому, механическую, каркасную функцию. Актиновые микрофибриллы и промежуточные филаменты, как и микротрубочки, построены из субъединиц. В силу этого их количество зависит от соотношения процессов полимеризации и деполимеризации. Для животных клеток, части клеток растений, грибов и водорослей характерен клеточный центр, в состав которого входят центриоли. Центриолъ (под электронным микроскопом) имеет вид «полого» цилиндра диаметром около 150 нм и длиной 300—500 нм. Ее стенка образована 27 микротрубочками, сгруппированными в 9 триплетов. В функцию центриолей входит образование нитей митотического веретена, которые также образованы микротрубочками. Центриоли поляризуют процесс деления клетки, обеспечивая расхождение сестринских хроматид (хромосом) в анафазе митоза. 2.3.3. Поток информацииЖизнедеятельность клетки как единицы биологической активности обеспечивается совокупностью взаимосвязанных, приуроченных к определенным внутриклеточным структурам, упорядоченных во времени и пространстве обменных (метаболических) процессов. Эти процессы образуют три потока: информации, энергии и веществ. Благодаря наличию потока информации клетка на основе многовекового эволюционного опыта предков приобретает структуру, отвечающую критериям живого, поддерживает ее во времени, а также передает в ряду поколений. В потоке информации участвуют ядро (конкретно ДНК хромосом), макромолекулы, переносящие информацию в цитоплазму (мРНК), цитоплазматический аппарат трансляции (рибосомы и полисомы, тРНК, ферменты активации аминокислот). На завершающем этапе этого потока полипептиды, синтезированные на полисомах, приобретают третичную и четвертичную структуры и используются в качестве катализаторов или структурных белков (рис. 2.7). Кроме основного по объему заключенной информации ядерного генома в эукариотических клетках функционируют также геномы митохондрий, а в зеленых растениях — и хлоропластов. 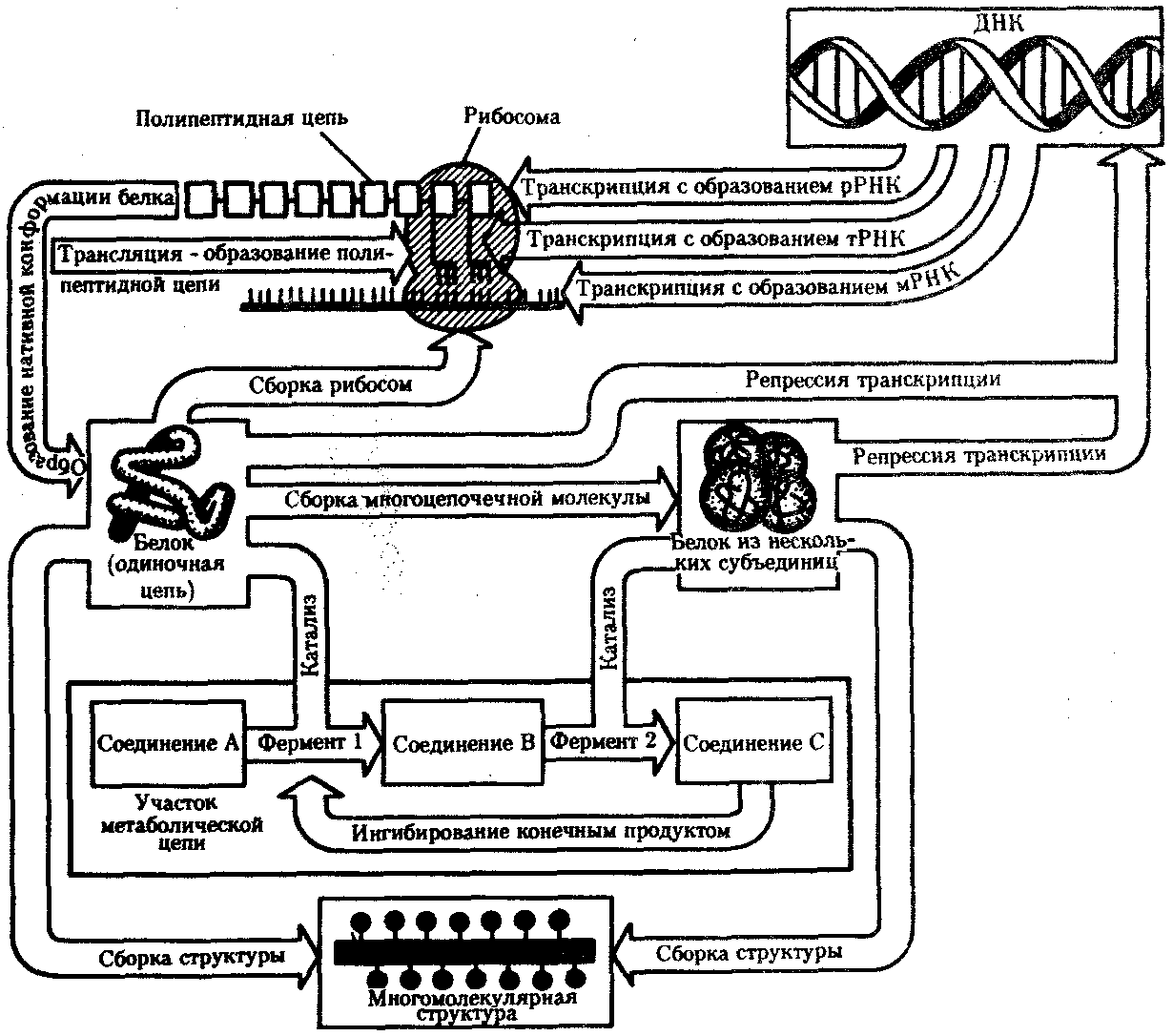 Рис. 2.7. Поток биологической информации в клетке 2.3.4. Внутриклеточный поток энергииПоток энергии у представителей разных групп организмов обеспечивается механизмами энергоснабжения —брожением, фото- или хемосинтезом, дыханием. Центральная роль в биоэнергетике клеток животных принадлежит дыхательному обмену. Он включает реакции расщепления низкокалорийного органического «топлива» в виде глюкозы, жирных кислот, аминокислот, а также использование выделяемой энергии для образования высококалорийного клеточного «топлива» в виде аденозинтрифосфата (АТФ). Энергия АТФ, непосредственно или будучи перенесена на другие макроэргические соединения (например, креатинфосфат), в разнообразных процессах преобразуется в тот или иной вид работы — химическую (синтезы), осмотическую (поддержание перепадов концентрации веществ), электрическую, механическую, ре-гуляторную. Макроэргическим называют соединение, в химических связях которого запасена энергия в форме, доступной для использования в биологических процессах. Универсальным соединением такого рода служит АТФ. Основное количество энергии заключено в связи, присоединяющей третий остаток фосфорной кислоты. 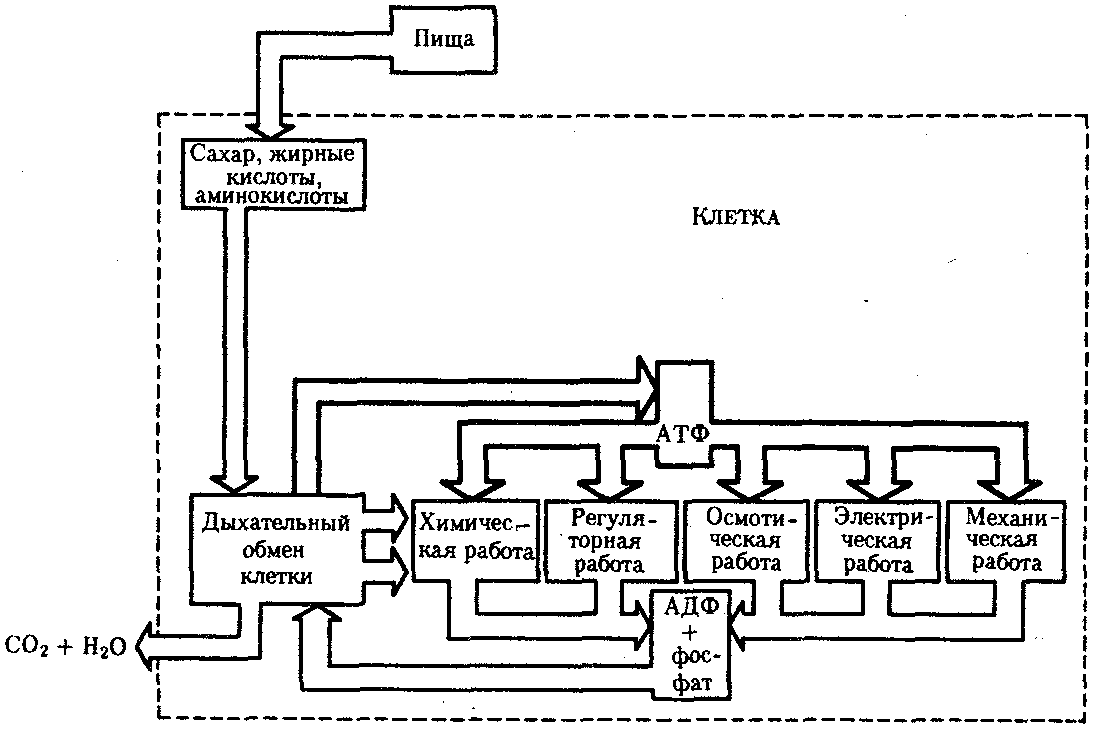 Рис. 2.8. Поток энергии в клетке Среди органелл животной клетки особое место в дыхательном обмене принадлежит митохондриям, выполняющим функцию окислительного фосфорилирования, а также матриксу цитоплазмы, в котором протекает процесс бескислородного расщепления глюкозы — анаэробный гликолиз (рис. 2.8). Из двух механизмов, обеспечивающих жизнедеятельность клетки энергией, анаэробный гликолиз менее эффективен. В связи с неполным (в отсутствие кислорода) окислением, прежде всего глюкозы, в процессе гликолиза для нужд клетки извлекается не более 10% энергии. Недоокисленные продукты гликолиза (пируват) поступают в митохондрий, где в условиях полного окисления, сопряженного с фосфорилированием АДФ до АТФ, отдают для нужд клетки оставшуюся в их химических связях энергию. Из преобразователей энергии химических связей АТФ в работу наиболее изучена механохимическая система поперечно-полосатой мышцы. Она состоит из сократительных белков (актомиозиновый комплекс) и фермента аденозинтрифосфатазы, расщепляющего АТФ с высвобождением энергии. Особенность потока энергии растительной клетки состоит в наличии фотосинтеза — механизма преобразования энергии солнечного света в энергию химических связей органических веществ. Механизмы энергообеспечения клетки отличаются эффективностью. Коэффициенты полезного действия хлоропласта и митохондрий, достигая соответственно 25 и 45—60%, существенно превосходят аналогичный показатель паровой машины (8%) или двигателя внутреннего сгорания (17%). 2.3.5. Внутриклеточный поток веществРеакции дыхательного обмена не только поставляют энергию, но и снабжают клетку строительными блоками для синтеза разнообразных молекул. Ими являются многие продукты расщепления пищевых веществ. Особая роль в этом принадлежит одному из этапов дыхательного обмена — циклу Кребса,осуществля-емому в митохондриях. Через этот цикл проходит путь углеродных атомов (углеродных скелетов) большинства соединений, служащих промежуточными продуктами синтеза химических компонентов клетки. В цикле Кребса происходит выбор пути превращения того или иного соединения, а также переключение обмена клетки с одного пути на другой, например с углеводного на жировой. Таким образом, дыхательный обмен одновременно составляет ведущее звено потока веществ, объединяющего метаболические пути расщепления и образования углеводов, белков, жиров, нуклеиновых кислот (рис. 2.9). 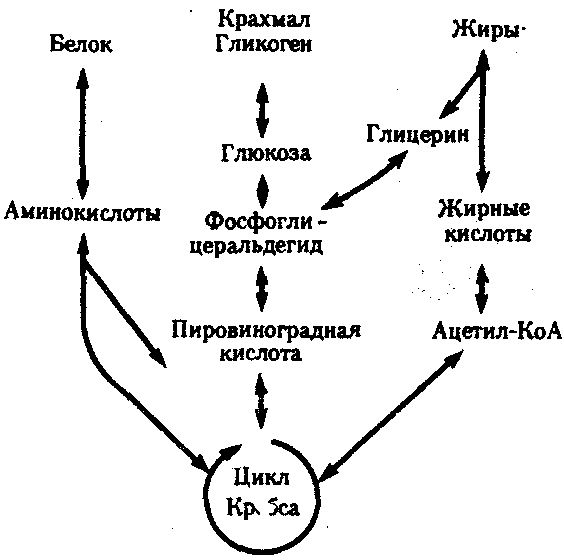 Рис. 2.9. Взаимосвязь внутриклеточного обмена белков, жиров и углеводов через цикл Кребса 2.3.6. Другие внутриклеточные механизмы общего значенияПотоки информации, энергии и веществ осуществляются непрерывно и составляют необходимое условие сохранения клетки как живой системы. Кроме структур и процессов, прямо включенных в названные потоки, в клетке функционируют механизмы, которые также являются жизненно необходимыми. Так, лизосомы, воздействуя ферментами на пиноцитированный или аутофагированный материал, обеспечивают гидролитическое расщепление макромолекул до низкомолекулярных соединений. Они же обусловливают разрушение внутриклеточных структур, утративших свое функциональное значение. Образовавшиеся при этом химические соединения включаются в потоки энергии, веществ и информации. Пероксисомы ликвидируют возникающие в клетке пероксиды, токсичные для живой протоплазмы. Организация внутриклеточных транспортных потоков обусловливается наличием и активностью микротрубочек, микрофибрилл. 2.3.7. Клетка как целостная структура. Коллоидная система протоплазмыВыше мы познакомились с различными по строению и функциям клеточными структурами. Однако, взаимодействуя с окружающей средой и отвечая на регуляторные стимулы, клетка ведет себя как целостная структура. Об этом свидетельствует однотипность реакции разных видов клеток на действие раздражителей, вызывающих переход клетки в возбужденное состояние1. Важная роль в функциональном объединении структурных компонентов и компартментов клетки принадлежит свойствам живой протоплазмы2. В целом ее принято рассматривать как особую многофазную коллоидную систему, или биоколлоид. От банальных коллоидных систем биоколлоид отличается сложностью дисперсной фазы. Основу ее составляют макромолекулы, которые присутствуют либо в составе плотных микроскопически видимых структур (органелл), либо в диспергированном состоянии, близком к растворам или рыхлым сетеобразным структурам типа гелей. Будучи коллоидным раствором в физико-химическом смысле, биоколлоид благодаря наличию липидов и крупных частиц проявляет одновременно свойства соответственно эмульсии и суспензии. На обширных поверхностях макромолекул оседают разнообразные «примеси», что ведет к изменению агрегатного состояния протоплазмы. Между крайними полюсами организации протоплазмы в виде вязких гелей и растворов имеются переходные состояния. При указанных переходах совершается работа, в результате которой осуществляются различные внутриклеточные превращения,—образование мембран, сборка микротрубочек или микрофиламентов из субъединиц, выброс из клетки секрета, изменение геометрии белковых молекул, приводящее к торможению или усилению ферментативной активности. Особенностью биоколлоида является также и то, что в физиологических условиях переходы протоплазмы из одного агрегатного состояния в другое (в силу наличия особого ферментативного механизма) обратимы. Названное свойство биоколлоида обеспечивает клетке способность при наличии энергии многократно совершать работу в ответ на действие стимулов. 2.4. ЗАКОНОМЕРНОСТИ СУЩЕСТВОВАНИЯ КЛЕТКИ ВО ВРЕМЕНИ2.4.1. Жизненный цикл клеткиЗакономерные изменения структурно-функциональных характеристик клетки во времени составляют содержание жизненного цикла клетки (клеточного цикла). Клеточный цикл — это период существования клетки от момента ее образования путем деления материнской клетки до собственного деления или смерти. Важным компонентом клеточного цикла является митотический (пролиферативный) цикл —комплекс взаимосвязанных и согласованных во времени событий, происходящих в процессе подготовки клетки к делению и на протяжении самого деления. Кроме того, в жизненный цикл включается период выполнения клеткой многоклеточного организма специфических функций, а также периоды покоя. В периоды покоя ближайшая судьба клетки не определена: она может либо начать подготовку к митозу, либо приступить к специализации в определенном функциональном направлении (рис. 2.10). Продолжительность митотического цикла для большинства клеток составляет от 10 до 50 ч. Длительность цикла регулируется путем изменения продолжительности всех его периодов. У млекопитающих время митоза составляет 1—1,5 ч, 02-периода интерфазы —2—5 ч, S-периода интерфазы — 6—10 ч. Биологическое значение митотического цикла состоит в том, что он обеспечивает преемственность хромосом в ряду клеточных поколений, образование клеток, равноценных по объему и содержанию наследственной информации. Таким образом, цикл является всеобщим механизмом воспроизведения клеточной организации эукариотического типа в индивидуальном развитии. Главные события митотического цикла заключаются в редупликации (самоудвоении) наследственного материала материнской клетки и в равномерном распределении этого материала между дочерними клетками. Указанным событиям сопутствуют закономерные изменения химической и морфологической организации хромосом — ядерных структур, в которых сосредоточено более 90% генетического материала эукари-отической клетки (основная часть внеядерной ДНК животной клетки находится в митохондриях). Хромосомы во взаимодействии с внехромосомными механизмами обеспечивают: а) хранение генетической информации, б) использование этой информации для создания и поддержания клеточной организации, в) регуляцию считывания наследственной информации, г) удвоение (самокопирование) генетического материала, д) передачу его от материнской клетки дочерним. Химическая организация и строение хромосом описаны в разд. 3.5.2. 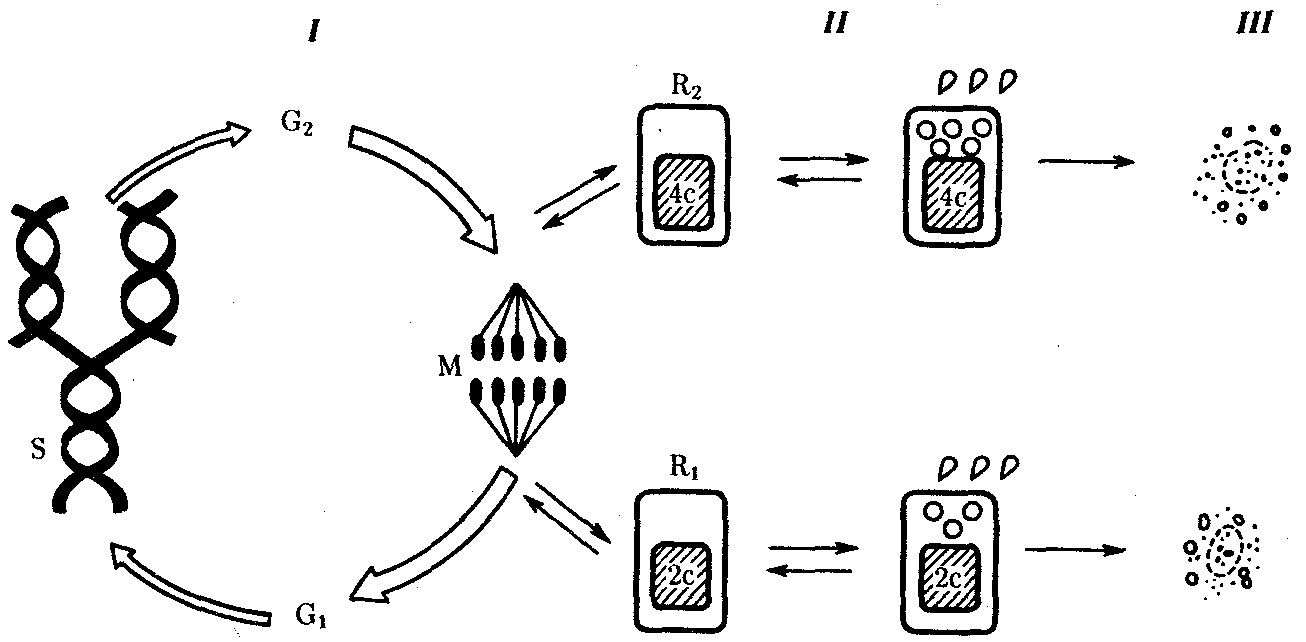 Рис. 2.10. Жизненный цикл клетки многоклеточного организма. I — митотический цикл; II — переход клетки в дифференцированное состояние; III— гибель клетки: G1 — пресинтетический период, G2 — постсинтетический (предмитотический) период, М —митоз, S — синтетический период, R1 и R2 — периоды покоя клеточного цикла; 2с —количество ДНК в диплоидном наборе хромосом, 4с —удвоенное количество ДНК 2.4.2. Изменения клетки в митотическом циклеПо двум главным событиям митотического цикла в нем выделяют репродуктивную и разделительную фазы, соответствующие интерфазе и митозу классической цитологии (рис. 2.11). 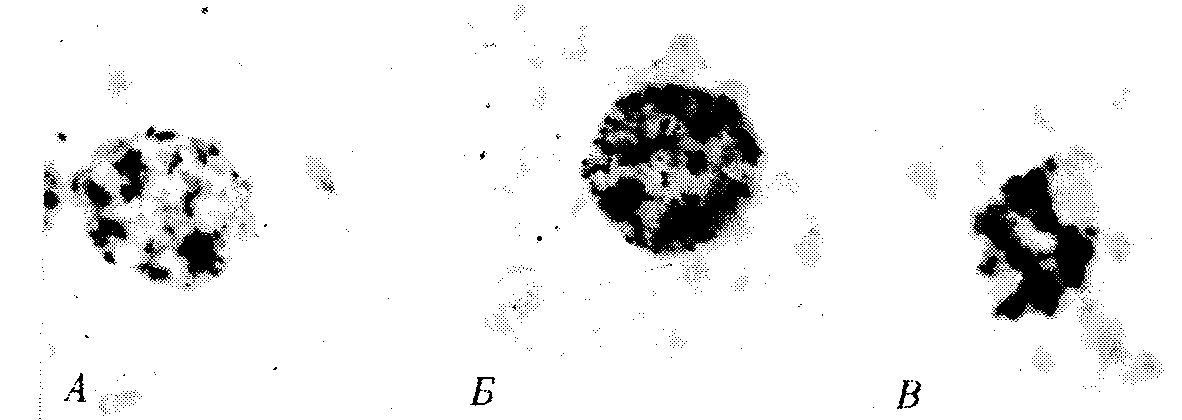 Рис. 2.11. Интерфазная и митотическая формы структурной организации хромосом. А — хроматин в интерфазном ядре; Б — включение в ядро радиоактивного предшественника ДНК в синтетическом периоде митотического цикла; В— клетка в анафазе митоза В начальный отрезок интерфазы (постмитотический, пресинтетический, или Gi-период) восстанавливаются черты организации интерфазной клетки, завершается формирование ядрышка, начавшееся еще в телофазе. Из цитоплазмы в ядро поступает значительное (до 90%) количество белка. В цитоплазме параллельно реорганизации ультраструктуры интенсифицируется синтез белка. Это способствует росту массы клетки. Если дочерней клетке предстоит вступить в следующий митотический цикл, синтезы приобретают направленный характер: образуются химические предшественники ДНК, ферменты, катализирующие реакцию редупликации ДНК, синтезируется белок, начинающий эту реакцию. Таким образом осуществляются процессы подготовки следующего периода интерфазы — синтетического. В синтетическом или S-периодв удваивается количество наследственного материала клетки. За малыми исключениями редупликация1 ДНК осуществляется полуконсервативным способом (рис. 2.12). Он заключается в расхождении биспирали ДНК на две цепи с последующим синтезом возле каждой из них комплементарной цепочки. В результате возникают две идентичные биспирали (детали см. 3.4.2.1). Молекулы ДНК, комплементарные материнским, образуются отдельными фрагментами по длине хромосомы, причем неодномоментно (асинхронно) в разных участках одной хромосомы, а также в разных хромосомах. Затем участки (единицы репликации — репликоны) новообразованной ДНК «сшиваются» в одну макромолекулу. В клетке человека содержится более 50 000 репликонов. Длина каждого из них около 30 мкм. Число их меняется в онтогенезе. Смысл редупликации ДНК репликонами становится понятным из следующих сопоставлений. Скорость синтеза ДНК составляет 0,5 мкм/мин. В этом случае редупликация нити ДНК одной хромосомы человека длиной около 7 см должна была бы занять около трех месяцев. Продолжительность же синтетического периода в клетках человека составляет 7—12 ч. 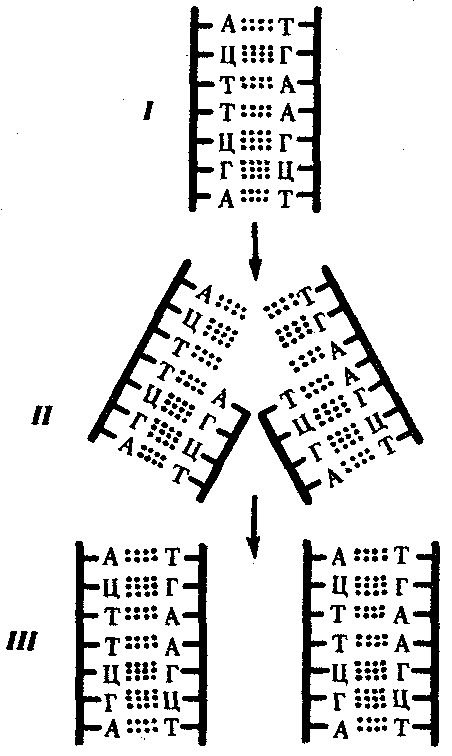 Рис. 2.12. Полуконсервативный редупликации ДНК. I — материнская биспираль ДНК; II—достраивание комплементарных полинуклеотидных цепей; III — две дочерние биспирали ДНК: А, Т, Г, Ц - символы азотистых оснований нуклеотидов Участки хромосом, в которых начинается синтез, называют точками инициации. Возможно, ими являются места прикрепления интерфазных хромосом к внутренней мембране ядерной оболочки. Можно думать, что ДНК отдельных фракций, о которых речь пойдет ниже (см. разд. 2.4.2), редуплицируется в строго определенной фазе S-периода. Так, большая часть генов рРНК удваивает ДНК в начале периода. Редупликация запускается поступающим в ядро из цитоплазмы сигналом, природа которого не выяснена. Синтезу ДНК в репликоне предшествует синтез РНК. В клетке, прошедшей S-период интерфазы, хромосомы содержат удвоенное количество генетического материала. Наряду с ДНК в синтетическом периоде интенсивно образуются РНК и белок, а количество гистонов строго удваивается. Примерно 1% ДНК животной клетки находится в митоховдриях. Незначительная часть митохондриальной ДНК редуплицируется в синтетическом, тогда как основная —в постсинтетическом периоде интерфазы. Вместе с тем известно, что продолжительность жизни митохондрий печеночных клеток, например, составляет 10 сут. Учитывая, что в обычных условиях гепатоциты делятся редко, следует допустить, что редупликация ДНК митохондрий может происходить независимо от стадий митотического цикла. Отрезок времени от окончания синтетического периода до начала митоза занимает постсинтетический (предмитотический), или G2-neриод интерфазы. Он характеризуется интенсивным синтезом РНК и особенно белка. Завершается удвоение массы цитоплазмы по сравнению с началом интерфазы. Это необходимо для вступления клетки в митоз. Часть образуемых белков (тубулины) используется в дальнейшем для построения микротрубочек веретена деления. Синтетический и постсинтетический периоды связаны с митозом непосредственно. Это позволяет выделить их в особый период интерфазы — препрофазу. В митозе можно выделить четыре фазы. Главные события по отдельным фазам представлены ниже и на рис. 2.13. 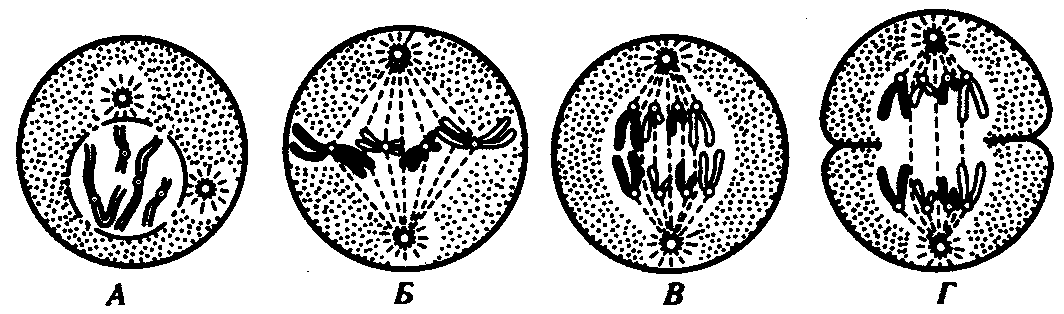 Рис. 2.13. Митоз в животной клетке. А — профаза; Б — метафаза; В — анафаза; Г — телофаза
Наряду с преобразованием строения и упорядоченными перемещениями хромосом обязательным для разделительной фазы цикла является построение митотического аппарата. Он состоит из системы микротрубочек (ахроматиновое веретено, или веретено деления светооптической микроскопии) и структур, поляризующих митоз1, т.е. обозначающих два полюса в клетке, к которым разойдутся дочерние хромосомы. В клетках млекопитающих и человека роль поляризующих структур выполняют центриоли. Митотический аппарат обеспечивает направленное перемещение дочерних хромосом в анафазе. Для этого необходим контакт микротрубочек со специализированными участками хромосом — центромерами (кинетохорами). При разрушении центромер в эксперименте расхождения хромосом к полюсам клетки не происходит. Природа сил, обеспечивающих расхождение, неизвестна. Предполагают, что функциональную основу митотического аппарата составляет механохимическая система, сходная с действующей в поперечно-полосатой мышце. В нее входят сократимые белки и фермент, катализирующий расщепление АТФ для обеспечения процесса энергией. Нарушения той или иной фазы митоза приводят к патологическим изменениям клеток. Отклонение от нормального течения процесса спирализации может привести к набуханию и слипанию хромосом. Иногда наблюдается отрыв участка хромосомы, который, если он лишен центромеры, не участвует в анафазном перемещении к полюсам и теряется. Отставать при движении могут отдельные хроматиды, что приводит к образованию дочерних ядер с несбалансированными хромосомными наборами. Повреждения со стороны веретена деления приводят к задержке митоза в метафазе, рассеиванию хромосом. При изменении количества центриолей возникают многополюсные или асимметричные митозы. Нарушение цитотомии приводит к появлению дву- и многоядерных клеток. Данные генетики и цитологии указывают на сохранение структурной индивидуальности хромосом в клеточном цикле. Есть свидетельства в пользу упорядоченного размещения хромосом в объеме интерфазного ядра. Особенности взаиморасположения хромосом могут иметь большое функциональное значение. Так, пространственная близость в клетках человека хромосом 13, 14, 15, 21 и 22-й пар, содержащих гены рРНК, объясняется, по-видимому, их участием в формировании ядрышка и образовании рибосом, а 11-й и 16-й хромосом —в образовании молекулы гемоглобина. Есть указания на то, что в клетках разных типов расположение хромосом не одинаково. На основе митотического цикла возник ряд механизмов, с помощью которых в том или ином органе количество генетического материала и, следовательно, интенсивность обмена могут быть увеличены при сохранении постоянства числа клеток. Удвоение ДНК клетки не всегда сопровождается ее разделением на две. Поскольку механизм такого удвоения совпадает с предмитотической редупликацией ДНК и оно сопровождается кратным увеличением количества хромосом, это явление получило название эндомитоза. С генетической точки зрения, эндомитоз — геномная соматическая мутация, о чем будет сказано ниже. Другое явление, сходное по результату и названное политенией, заключается в кратном увеличении содержания ДНК в хромосомах при сохранении их диплоидного количества. Эндомитоз и политения приводят к образованию полиплоидных клеток, отличающихся кратным увеличением объема наследственного материала. В таких клетках в отличие от диплоидных гены повторены более чем два раза. Пропорционально увеличению числа генов растет масса клетки, что повышает ее функциональные возможности. В организме млекопитающих полиплоидизация с возрастом свойственна печеночным клеткам. |