анатомия собак. Анатомия и физиология животных (1). Контрольная работа по дисциплине Анатомии и физиологии животных студент 1 курса спо затолокин А. М. (шифр аб21037С)
 Скачать 188.07 Kb. Скачать 188.07 Kb.
|

|
МИНИСТЕРСТВО СЕЛЬСКОГО ХОЗЯЙСТВА РОССИЙСКОЙ ФЕДЕРАЦИИ ФГБОУ ВПО РОССИЙСКИЙ ГОСУДАРСТВЕННЫЙ АГРАРНЫЙ ЗАОЧНЫЙ УНИРЕСИТЕТ ФАКУЛЬТЕТ АГРО-И БИОТЕХНОЛОГИЙ НАПРАВЛЕНИЕ ПОДГОТОВКИ: 35.02.15 « КИНОЛОГИЯ» Кафедра зоотехнии, производства и переработки продукции и животноводства Контрольная работа по дисциплине: Анатомии и физиологии животных Выполнил: студент 1 курса СПО Затолокин А.М. (шифр АБ-21/037С) Балашиха 2021г. Содержание Типы дробления зиготы. Особенности дробления и ранних стадий развития млекопитающих. Роль трофобласта……………………………3 Особенности строения стенки трахеи. Чем обусловлены эти особенности?..........................................................................................7 Каковы сущность и значение пищеварения? Опишите особенности ротового и желудочного пищеварения у жвачных животных………….9 Какие функции выполняет кора головного мозга? В чем заключается аналитическая и синтетическая деятельность центральной нервной системы у млекопитающих?.................................................................13 Используемая литература………..……………………………………………...15 Типы дробления зиготы. Особенности дробления и ранних стадий развития млекопитающих. Роль трофобласта. Оплодотворение играет чрезвычайно важную роль в развитии, но оно — лишь только первая его ступень. Зигота с ее новым генетическим потенциалом приступает к созданию многоклеточного организма. У всех животных это начинается с процесса, носящего название дробление — серии митотических делений, в результате которых огромный объем цитоплазмы яйца разделяется на многочисленные клетки меньшего размера. Такие клетки, образующиеся в период дробления, называются бластомерами. Объем зародыша во время дробления не увеличивается. В большинстве других случаев клеточной пролиферации в период между митозами происходит рост клеток; клетка увеличивается в объеме почти вдвое и затем делится. Такой рост приводит к четкому увеличению общего объема клеток при сохранении относительно постоянного отношения объема ядра к объему цитоплазмы. Однако в период дробления зиготы объем цитоплазмы не возрастает: огромная масса цитоплазмы зиготы разделяется на все более мелкие клетки. Такое деление цитоплазмы яйца, не сопровождающееся ростом, осуществляется путем выпадения интерфазного периода роста между делениями. Дробление — строго координированный процесс, находящийся под генетическим контролем. Видовые особенности процесса дробления у разных животных определяются двумя основными параметрами: количеством и распределением желтка в цитоплазме и цитоплазматическими факторами, которые влияют на ориентацию митотического веретена. Наибольшие сложности возникают при изучении дробления у млекопитающих. Яйца млекопитающих относятся к числу самых мелких в животном царстве, что делает затруднительной экспериментальную работу с ними. Диаметр зиготы человека составляет, например, всего лишь 100 мкм, и ее объем меньше одной тысячной объема яйца шпорцевой лягушки. Кроме того, по количеству зигот, которые могут быть получены от одной особи, млекопитающие несравнимы с морскими ежами или лягушками. Обычно у самки млекопитающего одновременно овулирует менее 10 яиц. Кроме того, воспроизведение вне организма матери условий, необходимых для нормального дробления эмбрионов создает дополнительные методические сложности. Однако, к сегодняшнему дню процесс дробления у млекопитающих описан достаточно подробно. Ооцит млекопитающих, высвобождаясь из яичника, попадает в яйцевод. Оплодотворение происходит в ампуле яйцевода — его отделе, расположенном вблизи яичника. В это время мейоз завершается, и приблизительно через сутки начинается первое деление дробления. 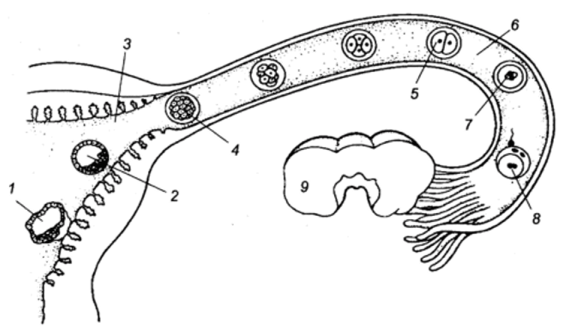 Рис. 1 Локализация ранних стадий развития в половых путях самки (Тисктапп Вир1езз в/ а1., 1972; по Гилберт С., 1993): 1 — ранняя стадия имплантации; 2 — бластоциста; 3 — матка; 4 — морула; 5 — двухклеточная стадия; 6 — яйцевод; 7 — первое деление дробления; 8 — оплодотворение; 9 — яичник. Деления дробления у млекопитающих относятся к числу самых медленных, встречающихся в животном царстве. Каждое из них продолжается от 12 до 24 часов. Тем временем дробящййся зародыш передвигается по яйцеводу по направлению к матке. Первые деления дробления происходят во время перемещения яйца по яйцеводу (рис. 1). Первая особенность дробления млекопитающих заключается в относительно медленном темпе делений. Второе важное отличие состоит в своеобразном расположении бластомеров относительно друг друга. Первое деление представляет собой нормальное меридиональное деление, т. е. плоскость деления проходит через полюса зиготы. Однако при втором делении один из двух бластомеров делится также меридионально, а второй — экваториально. Такой тип дробления называется чередующимся. Третьим важным отличием дробления у млекопитающих является выраженная асинхронность раннего дробления. Бластомеры у млекопитающих не делятся все одновременно, поэтому у зародышей не происходит равномерного нарастания числа бластомеров от 2-клеточной к 4- и 8-клеточной стадиям; зародыши часто содержат нечетное число клеток. Поскольку яйцеклетки млекопитающих относятся к изо- лецитальному типу, т. е. содержат минимальное количество равномерно распределенного в цитоплазме желтка, для млекопитающих характерно так называемое полное, или голоб- ластическое дробление. Это означает, что борозды дробления проходят через все яйцо. Таким образом, дробление млекопитающих является полным чередующимся асинхронным. Еще одно важное отличие дробления у млекопитающих от всех других типов дробления заключается в явлении компактизации. Бластомеры млекопитающих на 8-клеточной стадии располагаются рыхло, между ними остаются большие пространства. Однако после третьего деления дробления поведение бластомеров резко изменяется. Они внезапно сближаются, площадь контакта между ними максимально увеличивается, и они образуют компактный клеточный шар. Эта тесная упаковка стабилизируется плотными контактами, которые образуются между клетками, расположенными на поверхности шара, и изолируют лежащие внутри клетки. Между клетками внутри шара возникают щелевые контакты, которые позволяют малым молекулам и ионам переходить из клетки в клетку. Сегодня имеется много данных о том, что компактизация связана с явлениями, происходящими на клеточных поверхностях соседних бластомеров. Во-первых, до компактизации у каждого из восьми бластомеров наблюдаются далеко идущие изменения плазматической мембраны, известные под названием поляризации. Во-вторых, в процессе компактизации участвуют специфические белки клеточной поверхности. Одним таким белком является увоморулин — гликопротеин с молекулярной массой 120 000 дальтон. Антитела к молекуле увоморулина вызывают декомпактизацию морулы и подавляют прикрепление клеток друг к другу. В-третьих, плазматическая мембрана при компактизации может также изменяться вследствие реорганизации цитоскелета. На соседних клеточных поверхностях в результате образования актиновых микрофиламентов возникают микроворсинки, прикрепляющие клетки друг к другу. Именно на ворсинках функционирует увоморулин как медиатор межклеточной адгезии. Уплощение поверхностей соседних бластомеров в месте их контакта может вызываться укорочением микроворсинок путем деполимеризации актина. 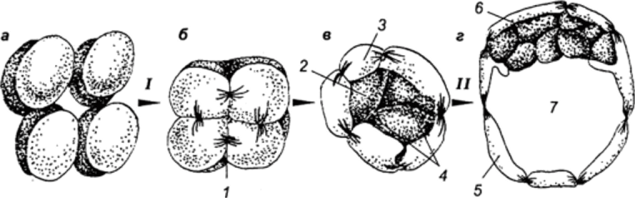 Рис. 2 Основные стадии доимплантационного развития млекопитающих (по Гилберт С., 1993): Клетки компактизованного зародыша делятся и образуют 16-клеточную морулу (рис. 19). Такая морула состоит из небольшого числа внутренних клеток, окруженных более многочисленными наружными клетками. Большая часть потомков наружных клеток становится клетками трофобласта (или трофэктодермы). Эта группа клеток не образует эмбриональных структур, а превращается в хорион, участвующий в образовании плаценты. Сам зародыш формируется потомками внутренних клеток 16-клеточного зародыша. Эти клетки образуют внутреннюю клеточную массу (ВКМ), которая и дает начало зародышу. Клетки ВКМ отличаются от клеток трофобласта не только по своему виду, но и по спектру белков, которые они синтезируют на этой ранней стадии. К стадии 64 клеток внутренняя клеточная масса и клетки трофобласта превращаются в полностью сформированные клеточные слои, ни один из которых не поставляет клеток другой группе. Многочисленные эксперименты показали, что судьба клетки зависит от ее пространственного положения в составе морулы. Если любой бластомер 4-клеточного зародыша мыши поместить на наружную поверхность рыхлой массы бластомеров другого зародыша, то из пересаженной клетки разовьется ткань трофобласта. Если большинство клеток бластоцисты дает начало тро- фобласту, то сколько же клеток фактически образует зародыш? Один из путей для получения ответа на этот вопрос заключается в создании аллофенных мышей. Аллофенные мыши являются результатом развития химерных зародышей, полученных путем слияния двух зародышей на ранних стадиях дробления (обычно 4- или 8-клеточных). У двух генетически различающихся зародышей удаляют прозрачную оболочку, и зародышей приводят в контакт для их слияния и образования единой бластоцисты. Полученные бластоцисты имплантируют в матку самки-реципиента. Рождающиеся мышата содержат клетки, которые произошли от обоих зародышей. Это хорошо видно, если используемые линии мышей различаются по окраске шерсти. Когда агрегируются бластомеры белых и черных мышей, то обычно развивается мышь с черными и белыми полосами. На основании изучения рождающихся аллофенных мышат был сделан вывод, что абсолютное число бластомеров, образующих зародыш, может быть равно 3 или ненамного превышать эту величину. Первоначально морула не имеет внутренней полости. Однако в процессе так называемой кавитации клетки трофобласта секретируют в морулу жидкость, что приводит к образованию полости бластоцисты. Внутренняя клеточная масса располагается на одной стороне полого шара, образованного клетками трофобласта. Такая структура называется бластоцистой, и ее образование является еще одной отличительной особенностью дробления млекопитающих. Пока зародыш перемещается по яйцеводу к матке, бластоциста увеличивается в объеме. В плазматических мембранах клеток трофэктодермы имеется натриевый насос (Ма+/К+-АТФаза), который переносит ионы натрия в центральную полость. Такое накопление ионов натрия вызывает поступление в полость воды осмотическим путем, в результате чего размеры полости бластоцисты увеличиваются. В период перемещения зародыша прозрачная оболочка предотвращает прилипание бластоцисты к стенкам яйцевода. Если у человека происходит такое прилипание, наблюдается «трубная беременность». Это крайне опасное явление, поскольку имплантация зародыша в яйцевод может вызвать угрожающее жизни кровотечение. Однако, когда зародыш достигает матки, он должен освободиться от прозрачной оболочки, чтобы прикрепиться к стенке матки. Бластоциста мыши освобождается от оболочки путем лизиса в ней небольшого отверстия, через которое она выдавливается при увеличении своего объема. Гистологические данные свидетельствуют о том, что одна из клеток тро- фобластической стенки образует вырост, который приходит в контакт с прозрачной оболочкой. На плазматической мембране этого выроста локализована трипсиноподобная протеаза стрипсин, которая лизирует отверстие в фибриллярном матриксе оболочки. Освободившись от оболочки, бластоциста может вступить в непосредственный контакт с маткой. Здесь трофобласт будет секретировать другую трипсиноподобную протеазу, активатор плазминогена. Этот расщепляющий белок фермент необходим для разрушения ткани матки, чтобы бластоциста могла погрузиться в ее стенку. 2. Особенности строения стенки трахеи. Чем обусловлены эти особенности? Трахея — trachea служит для проведения воздуха в легкие и обратно. Это трубка с постоянно зияющим просветом, что обеспечивается имеющимися в ее стенке не замкнутыми сверху кольцами из гиалинового хряща. Изнутри трахея выстлана слизистой оболочкой, снаружи покрыта в области шеи адвентицией, а в грудной полости —серозной оболочкой. Трахея простирается от гортани до основания сердца, где делится на два бронха. Место деления на-зывается бифуркацией трахеи. Длина трахеи зависит от длины шеи, в связи с чем количество ее хрящевых колец колеблется от 32 у свиньи до 60 у лошади.В области шеи трахея прилежит своей дорсальной поверхностью к длинной мышце головы и шеи, а также пищеводу; ее вентральная поверхность прикрыта грудино-головной, грудино-щитовидной и грудино-подьязычной мышцами. В грудной полости трахея лежит дорсально от сердца между листками средостения. 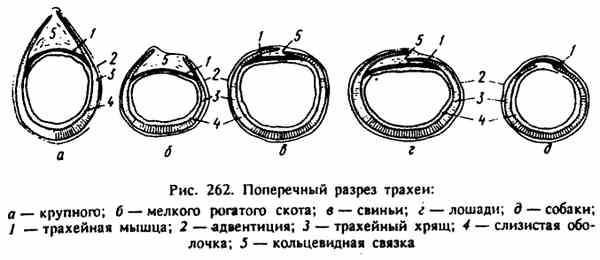 Трахейные хрящи соединяются друг с другом кольцевидными (трахейными) связками. Свободные концы хрящей входят в состав дорсальной перепончатой поверхности трахеи. Эта поверхность образована поперечными кольцевидными связками, соединяющими концы трахейных хрящей, и трахейными мышцами, расположенными под слизистой оболочкой. Сокращения этих мышц могут значительно суживать просвет трахеи. Слизистая оболочка трахеи покрыта мерцательным эпителием, содержит серозные, слизистые и смешанные железы. Подслизистый слой развит между хрящами и в дорсальной стенке трахеи. У крупного рогатого скота трахея сжата с боков, дорсальный край ее заострен. Диаметр трахеи 4—4,5 см; хрящей 45—50 шт. Бифуркация расположена на уровне 5-го ребра. Краниальнее бифуркации от трахеи отделяется трахейный бронх для правой краниальной доли легкого. У с в и н ь и трахея цилиндрическая, хрящей 32—36. Бифуркация трахеи находится на уровне 4—5-го ребра. Краниальнее бифуркации отделяется, как и у жвачных, трахейный бронх для правой краниальной доли легкого. Концы хрящей заходят один на другой. У лошади трахея слегка сплюснута сверху вниз, ее диаметр 4— 7 см; хрящевых колец 48—60. Бифуркация трахеи — на уровне 5— 6-го ребра. Концы хрящей накладываются друг на друга. У собаки трахея цилиндрическая, хрящей 42—46. Бифуркация расположена на уровне 4-го ребра. Концы хрящей друг на друга не накладываются. Трахеальная поперечная мышца расположена дорсально от хрящей. 3. Каковы сущность и значение пищеварения? Опишите особенности ротового и желудочного пищеварения у жвачных животных. Строение и деятельность пищеварительного аппарата взрослых жвачных обеспечивает переваривание большого количества грубого корма и превращение его питательных веществ в биологически ценные продукты питания человека. Однако в начале постэмбриональной жизни наблюдается кишечный тип пищеварения, обеспечивающий эффективное использование только легкопереваримых кормов, преимущественно молока. Молоко, которое является основной пищей телят в ранний молочный период, в их ротовой полости смешивается со слюной, что способствует образованию рыхлого молочного сгустка. При быстром же выпаивании молока оно не успевает хорошо перемешаться со слюной. Сгусток в результате образуется плотный, что нарушает пищеварение. Более того, из плотного сгустка затрудняется усвоение содержащихся в молоке питательных веществ. Это имеет место преимущественно при выпойке молока из ведра, когда телята заглатывают его большими порциями. При этом часть молока из пищевого желоба может прорваться в рубец, что нежелательно, так как оно там не переваривается и даже может загнивать, вызывая расстройство процесса пищеварения. Неблагоприятные для пищеварения условия образуются и тогда, когда молочная пища в сычуге не створаживается, что имеет место при выпаивании телятам, к примеру, соевого молока. В этом случае белок не задерживается в сычуге, проходит транзитом в кишечник, и это плохо, поскольку отсутствует фаза желудочного пищеварения. Под воздействием ферментов кишечных соков, поджелудочной железы и желчи белки, жиры и углеводы молока расщепляются до аминокислот, жирных кислот, простых сахаров и в этом виде всасываются. Всасывается в тонком кишечнике до 95-98% питательных веществ молока. У телят в начале постэмбриональной жизни очень низкая активность ферментов, расщепляющих растительные корма, а также крахмал и сахар. Так, в 10-дневном возрасте в их желудочно-кишечном тракте концентраты растительного происхождения перевариваются только лишь на 16%. Но раннее приучение животных к растительным кормам стимулирует функцию пищеварительных желез. Поэтому с 5-10-дневного возраста телят уже приучают к потреблению концентратов. С 10-15-дневного возраста им вводят в рацион корнеплоды в измельченном виде (в виде кашицы). Хороший эффект получают при скармливании вареного картофеля. Скармливанием телятам-молочникам комбикормов–стартеров и высококачественных сочных и грубых кормов удается стимулировать переваримость и усвоение питательных веществ. У телят, рано приученных к потреблению грубых кормов и концентратов, уже в 1-1,5-месячном возрасте переваримость клетчатки может достигать 50%, а в возрасте 2-3 месяца – 70% от максимальной. Обычно переход с кишечного типа пищеварения на желудочно-кишечный у телят завершается к 2-4-месячному возрасту. К этому времени рубец и другие преджелудки телят заселяются микрофлорой, свойственной взрослым животным, которая играет главенствующую роль в переваривании растительных кормов. При этом, на переваримости кормов сказывается степень заселения преджелудков микрофлорой. В свою очередь скорость и степень заселения преджелудков микрофлорой стимулируется способом выращивания телят. При содержании их в индивидуальных клетках эти процессы замедляются. Так, у телят, размещенных в индивидуальных клетках, плотность микрофлоры меньше. Поэтому количество переваренных органических веществ в сложном желудке таких животных составило всего лишь 13,5%, в то время как у содержащихся на подсосе – 34-35%. В возрасте 2-3 месяцев процессы рубцового пищеварения развиваются в такой степени, что теленок может потреблять и переваривать значительные количества растительных кормов. К полной замене молочных кормов растительными телята должны поедать не менее 0,8-1,0 кг сухого вещества растительных кормов. В процессе переваривания потребляемого корма у жвачных важную роль играет микрофлора преджелудков. Общее количество микрофлоры в рубце взрослых животных составляет 1010 степени в 1 г содержимого. В рубце постоянно присутствуют живые микроорганизмы, мертвые и разрушающиеся. Самые важные – это целлюлозолитические. Их содержится 109 на 1 г содержимого рубца. При скармливании зерновых, крахмалистых и сахаристых кормов увеличивается содержание амилолитических бактерий. Один вид микроорганизмов влияет на развитие других, угнетая или стимулируя их рост. Так, увеличение количества лактобактерий сдерживает рост амилолитических, стрептококков и наоборот. Скорость размножения микроорганизмов высокая и составляет, как минимум, 4-5 поколений в день. В 1 г содержимого рубца находится около 1 млн. инфузорий. Они разрыхляют, измельчают корм, используют для своего питания клетчатку. Переваривают белки, крахмал, сахара, клетчатку, накапливают в своем теле полисахариды. Общая масса микрофлоры рубца составляет 6-7 кг или 10% его содержимого. В 1 мл рубцового содержимого находится от 6 до 40 млрд. бактерий. Их всего может быть около 150 видов. Кроме того, в рубце содержится до 23 видов грибов (дрожжей и плесеней) и примерно 60 видов простейших (инфузорий). Они подвергают корм механической обработке. Бактерии и дрожжи расщепляют почти все растворимые углеводы корма. Сахар и крахмал на 80-85%, клетчатку – на 40-45%. При этом образуются жирные кислоты, представленные уксусной (65-70%), пропионовой (15-20%) и масляной (8-12%) кислотами. Общая масса образуемых в преджелудках взрослых животных ЛЖК составляет 4-4,5 кг в сутки. До 80% образованной уксусной кислоты усваивается тканями молочной железы и используется для синтеза жира молока. Пропионовая кислота является предшественником глюкозы, откладываемой в форме гликогена мышц и печени. Оптимальное соотношение уксусной и пропионовой кислот – 3:1. Из масляной кислоты в печени и других тканях синтезируется жир молока. Однако при интенсивном и избыточном образовании масляной кислоты она участвует в синтезе высокотоксичных продуктов обмена – кетоновых тел. Образование уксусной кислоты стимулируется скармливанием грубых кормов. В рубце переваривается до 70% сухого вещества рациона. Рн содержимого рубца составляет 6,5-7,4 ед. и сдвигается в кислую сторону в период наиболее интенсивного сбраживания корма. Нейтрализация кислот в рубце осуществляется с помощью слюны, суточное образование которой доходит до 200 л. На функциональной деятельности желудочно-кишечного тракта сказывается тип, уровень кормления и физическое состояние кормов. Так, степень измельчения корма сказывается на переваримости клетчатки. При скармливании тонко измельченных грубых кормов переваримость клетчатки составляла 20-26%, в то время как дача не измельченных или крупно измельченных грубых кормов сопровождалась повышением переваримости клетчатки до 60-65%. Скармливание тонко измельченных кормов и кормов, богатых легкопереваримыми углеводами, приводит к сокращению времени жвачки, снижению выделения слюны, закисанию содержимого рубца, понижению содержания в нем аммиака, увеличению уровня ЛЖК в рубце. При таком кормлении в составе ЛЖК повышается содержание пропионовой кислоты до 30-32%, масляной – до 10-11%, а уксусной – уменьшается до 50-52%. Такой тип рациона приемлем для откармливаемых животных, поскольку из пропионовой кислоты в конечном итоге образуется жир, который откладывается в теле животного и в конечном итоге повышается откормочная и мясная продуктивность животных. В том случае, когда требуется формировать племенных животных с высокой молочностью, то целесообразно использовать рационы с преобладанием грубых кормов, то есть обогащать рацион клетчаткой. При скармливании грубых кормов образуется больше уксусной кислоты и ее содержание в составе ЛЖК повышается до 70%, а количество пропионовой - уменьшается до 18-20%. Содержание масляной кислоты остается практически без изменений, то есть составляет 8-10%. При выращивании молодняка на рационах с преобладанием объемистых кормов, у них уже к 2-летнему возрасту формируется рубец и сетка емкостью на 37%, а длина кишечника на 17% больше, чем у телят на рационах с концентратами. Вследствие этого повышается на 17% переваримость клетчатки и на 15,1% - уровень желудочного пищеварения. Важно поддерживать оптимальное соотношение легко- и труднорасщепляемого протеина в рационе жвачных. При высоком содержании легкопереваримого протеина он быстро гидролизуется микрофлорой рубца с образовпанием аммиака. Аммиак поступает в печень, где из него образуется мочевина. Определенное ее количество возвращается со слюной в рубец и используется для питания микрофлоры рубца. Часть же мочевины выводится из организма животных с мочой, то есть расходуется нерационально. Поэтому важно сдерживать, то есть снижать расщепляемость протеинов в рубце животных. Этого добиваются, защищая протеин корма от разрушения в рубце. В рубце происходит переваривание липидов корма, в процессе которого ненасыщенные жирные кислоты превращаются в насыщенные. В результате увеличивается общее содержание сырого жира в пищеварительном тракте и его становится больше, чем принято с кормом. Практически в рубце полностью сбраживаются сахара, крахмал и до 80% клетчатки. Микрофлора преджелудков синтезирует белки и витамины группы В и К, используя клетчатку, целлюлозу корма, а также аммиак, образуемый при распаде протеинов корма. Бактерии могут использовать также и аммиак синтетических соединений. Поэтому крупному рогатому скоту можно скармливать до 150 г карбамида в сутки, овцам – до 18 г. Но при этом следует обеспечивать в рационе оптимальное соотношение легкопереваримых углеводов и протеина. При их дефиците аммиак, образуемый при распаде синтетических соединений, не усваивается микрофлорой, а поскольку распад азотсодержащих соединений происходит быстро, то высокое содержание аммиака может привести к отравлениям животных. Сами микроорганизмы, содержащиеся в преджелудках, по мере их отмирания с кормовой массой попадают в сычуг и тонкий отдел кишечника, где перевариваются и за счет образующихся элементов питания обеспечивается до 30% потребности организма в белке. За счет микроорганизмов жвачные получают до 1,5 кг переваримого белка в сутки. Видовой состав микрофлоры преджелудков зависит от вида корма. При смене состава рациона ее количество и виды меняются, что может неблагоприятно сказываться на процессе пищеварения. Поэтому для жвачных важен постепенный переход от одного рациона к другому. Клетчатка для животных является не только источником энергии. Она участвует также в нормализации моторики преджелудков. При ее дефиците понижается переваримость клетчатки, корм быстрее переходит из преджелудков в кишечник. Введение в рацион жвачных простых углеводов также снижает переваримость клетчатки. В этом случае микроорганизмы предпочитают простые углеводы и не используют клетчатку. В переходный период в преджелудках всасывается до 20% питательных веществ корма. По мере роста животных переваривание в преджелудках увеличивается до 50%, а усвоение клетчатки повышается в три раза. Основным местом всасывания органических веществ является слизистая сложного желудка, где всасывается более 50% питательных веществ. В преджелудках почти полностью всасываются ЛЖК. В тонком кишечнике всасывается до 30% и остальное количество питательных веществ всасывается в толстом кишечнике. 4. Какие функции выполняет кора головного мозга? В чем заключается аналитическая и синтетическая деятельность центральной нервной системы у млекопитающих? Деятельность коры больших полушарий обеспечивает постоянный анализ и синтез раздражений, падающих на организм из внешней среды и возникающих в нем самом. Будучи внешне противоположными, как внешне противоположны лежащие в их основе явления торможения и возбуждения, анализ и синтез неразрывно друг с другом связаны и друг без друга невозможны. Нервный синтез — это объединение друг с другом различных раздражителей в их действии на организм, их связывание механизмом рефлекса с теми или иными функциями организма. Все явления замыкания нервной связи между различными агентами, раздражающими рецепторы, и ответной деятельностью организма являются проявлением синтетической деятельности нервной системы. Постоянное замыкание, обеспечивающее наличие безусловных, врожденных рефлексов, ведущих к возникновению всегда одной и той же реакции на раздражение рецепторного поля каждого такого рефлекса, относится к простым проявлениям синтетической деятельности. Переменное замыкание, обеспечивающее выработку временных связей, благодаря которым деятельность организма определяется условиями действия каждого раздражителя, его сигнальным значением, является высшей формой синтеза, корковым синтезом, деятельностью коры головного мозга. Под анализаторами принято понимать совокупность всех нервных образований, отвечающую за восприятие какого-либо вида информации и включающую в себя периферический отдел — рецепторы. В центральную часть каждого анализатора, образуя его низшие отделы, входят: образования спинного и продолговатого мозга, центры, расположенные между продолговатым мозгом и корой, сама кора головного мозга. Вся аналитическая деятельность основана на процессе торможения, так как лишь этот процесс обеспечивает ограничение иррадиации возбуждения по центральной нервной системе. Выделяемые аналитической деятельностью раздражения являются тем более дробными, чем ограниченнее распространение по центрам нервного возбуждения, возникающего в них при стимуляции каждого рецепторного образования. Деятельность низших центров обеспечивает лишь такую форму анализа, в результате которой раздражение каждой группы рецепторов вызывает различный, но для рецепторного поля каждого рефлекса всегда относительно постоянный рефлекторный ответ. Данный анализ не только груб, но и статичен. Распределение торможения и возбуждения зависит прежде всего от того, с каких рецепторов и с какой частотой и силой импульсы поступают в низшие центры, а не от тех условий, в которых организму ранее наносились раздражения. Кора головного мозга обеспечивает осуществление высшего анализа, основанного на условном торможении, на торможении, которое формируется в коре мозга в зависимости от условий действия раздражителей, в зависимости от их подкрепления или неподкрепления. Благодаря процессу внутреннего, условного торможения из массы раздражений, постоянно и непрерывно доходящих до коры, во временные связи с теми или иными функциями вступают только те агенты и комплексы агентов, действие которых подкрепляется безусловным раздражителем. Используемая литература. Анатомия домашних животных. Учебник. 8-е издание Климов А.Ф., Акаевский А.И. 2011г 1040 с. Издательство Лань Биология. Пособие для поступающих в вузы. Ярыгин В.Н. Изд. 11 2009г- 439стр. Биология промысловых зверей России: учебное пособие для вузов. В.И. Машкин – Киров: Вятская ГСЧФ 2007 Основы физиологии и этологии животных. В.Ф. Лысов, В.И. Максимов Учебник и учебное пособие для студентов. КолосС 2004- 248 с. Писменская В.Н., Боев В.И. Практикум по анатомии и гистологии сельскохозяйственных животных. М.: КолосС, 2010. С. 201 |