генетика. контрольная работа. Контрольная работа по дисциплине Генетика растений и животных
 Скачать 316.55 Kb. Скачать 316.55 Kb.
|

|
Контрольная работа по дисциплине «Генетика растений и животных» Вариант №18 Новосибирск 2021 СодержаниеВопрос №12 3 Правило единообразия гибридов первого поколения (приведите схемы скрещивания). Гомозиготность и гетерозиготность 3 Вопрос №30 5 Полиплоидия, причины возникновения и значение для селекции 5 Вопрос №45 8 Факторы, изменяющие генетическую структуру популяции 8 Вопрос №70 14 Вопрос №94 15 СЛОВАРЬ ТЕРМИНОВ 16 СПИСОК ЛИТЕРАТУРЫ 17 Вопрос №12Правило единообразия гибридов первого поколения (приведите схемы скрещивания). Гомозиготность и гетерозиготностьЗакон единообразия гибридов первого поколения (первый закон Менделя) утверждает, что у гибридов первого поколения (F1) от скрещивания форм, различающихся только по одному альтернативному признаку (например, жёлтые или зелёные семена), проявляется признак только одного из родителей. Такое преобладание одного из признаков родителей у гибридов Мендель назвал доминированием, а соответствующий признак – доминантным. Альтернативные признаки, не проявляющиеся у гибридов F1, он назвал рецессивными. Позднее было установлено, что полное доминирование одних признаков над другими представляет собой широко распространённое явление не только у растений, но и у животных, грибов, микроорганизмов (рис. 1.). В некоторых случаях при скрещивании имеет место промежуточный фенотип (неполное доминирование). Так, у львиного зева (Antirrhinum) цветки гибридных растений первого поколения от скрещивания родителей с малиновыми и белыми цветками всегда розовые. Бывает также, что в потомстве F1 проявляются признаки обоих родителей (кодоминирование). Например, если один из родителей имеет группу крови А, а другой – В, то в крови их детей присутствуют антигены, характерные для обеих групп крови. Г. Менделем впервые был установлен факт, свидетельствующий о том, что растения, сходные по внешнему виду, могут резко отличаться по наследственным свойствам. Особи, не дающие расщепления в следующем поколении, получили название гомозиготных. Особи, в потомстве у которых обнаруживается расщепление признаков, назвали гетерозиготными [4]. Гомозигота – особь, содержащая в соматических клетках одинаковые аллели данного гена (например, АА / аа). Переход гена в гомозиготное состояние приводит к проявлению в структуре и функции организма (фенотипе) рецессивных аллелей, эффект которых при гетерозиготности подавляется доминантными аллелями. Тестом на гомозиготность служит отсутствие расщепления при определённых видах скрещивания. Гомозиготный организм образует по данному гену только один вид гамет [1]. 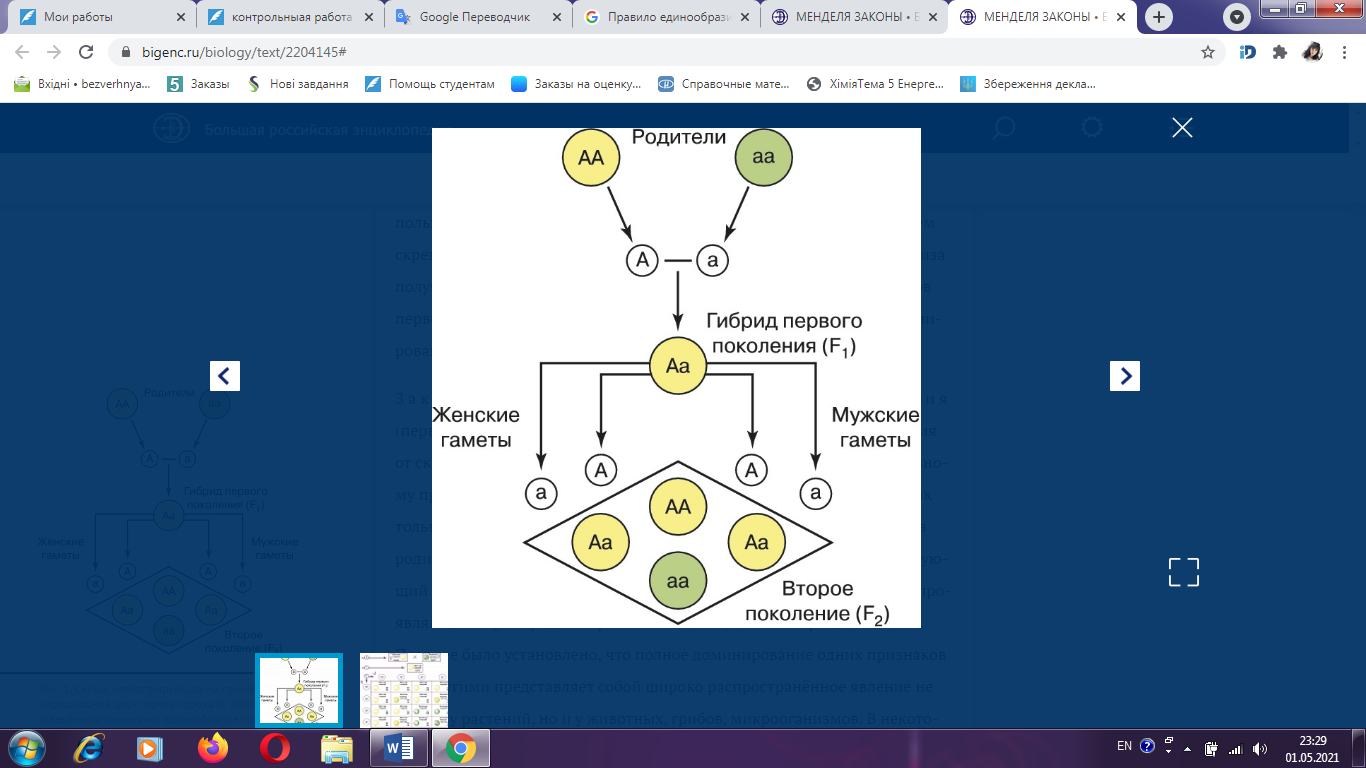 Рис. 1. Схема, иллюстрирующая на примере скрещивания двух сортов гороха (с жёлтыми и зелёными семенами) единообразие F1 (первый закон Менделя) и расщепление признаков у потомства F2с преобладанием доминантного фенотипа над рецессивным в соотношении 3:1. Гетерозигота – особь, содержащая в соматических клетках разные аллели данного гена (например, Аа). При гетерозиготности эффект вредных и летальных рецессивных аллелей подавляется присутствием соответствующего доминантного аллеля и проявляется только при переходе этого гена в гомозиготное состояние. Поэтому гетерозиготность широко распространена в природных популяциях и является, по-видимому, одной из причин гетерозиса. Маскирующее действие доминантных аллелей при гетерозиготности – причина сохранения и распространения в популяции вредных рецессивных аллелей. Их выявление (например, путём испытания производителей по потомству) осуществляется при любой племенной и селекционной работе, а также при составлении медико-генетических прогнозов [7]. Вопрос №30Полиплоидия, причины возникновения и значение для селекцииЧрезвычайно ценным источником изменчивости для селекции растений служит полиплоидия. Увеличение числа наборов хромосом у культурных растений сыграло выдающуюся роль в эволюции видов и селекции. Народная селекция, не зная самого явления полиплоидии, давно использовала ее как источник изменчивости в создании ряда таких ценнейших культур, как пшеница, овес, хлопчатник, картофель, а также в цветоводстве. Изучение полиплоидии позволило овладеть этим источником изменчивости растений. Число искусственно созданных полиплоидов с каждым годом прогрессивно растет. В настоящее время только у ржи получено несколько десятков тетраплоидов. Применение колхицина ускорило получение полиплоидов. При этом успех зависит от способа обработки тканей, вида растения, стадии воздействия [3]. Соматические клетки растений и животных, как правило, содержат двойное (диплоидное) число хромосом (2n). В отличие от соматических, половые клетки имеют уменьшенное (гаплоидное) число хромосом (n). В гаплоидных клетках каждая хромосома единична, не имеет парной себе гомологичной. Гаплоидное число хромосом в клетках организмов одного вида называется основным, или базовым, а совокупность генов, заключённую в таком гаплоидном наборе, – геномом. Гаплоидное число хромосом в половых клетках возникает вследствие редукции (уменьшения) вдвое числа хромосом в мейозе, а диплоидное число восстанавливается при оплодотворении (довольно часто у растений в диплоидной клетке бывают так называемые В-хромосомы, добавочные к какой-либо из хромосом; роль их мало изучена, хотя у кукурузы, например, всегда имеются подобные). Число хромосом у различных видов растений весьма разнообразно. Так, один из видов папоротника (Ophioglosum reticulata) имеет в диплоидном наборе 1260 хромосом, а у самого филогенетически развитого семейства сложноцветных вида Haplopappus gracilis – всего 2 хромосомы в гаплоидном наборе. Если изменяется число наборов хромосом (3n, 4n, 5n и т.д.), то говорят о полиплоидии. Организмы с соответственным кратным увеличением наборов хромосом (плоидности) в клетках называются триплоидами, тетраплоидами, пентаплоидами и т.д. или в целом – полиплоидами [2]. Кратное увеличение числа хромосом в клетках может возникать под действием высокой или низкой температуры, ионизирующих излучений, химических веществ, а также в результате изменения физиологического состояния клетки. Механизм действия этих факторов сводится к нарушению расхождения хромосом в митозе или мейозе и образованию клеток с кратно увеличенным числом хромосом по сравнению с исходной клеткой. Из химических агентов, вызывающих нарушение правильного расхождения хромосом, наиболее эффективен колхицин, препятствующий образованию нитей веретена деления клетки. Возникновение полиплоидных клеток, по мнению М.Е. Лобашева, может происходить по ряду причин. При нарушении митоза клеток соматической ткани в процессе развития организма формируется митотическая полиплоидия. При нарушении мейоза, приводящего к неправильному расхождению хромосом и образованию диплоидных нередуцированных гамет, слияние которых в процессе оплодотворения дает начало организму с умноженным набором хромосом, наблюдается мейотическая полиплоидия. R.A. McFeely считает, что триплоидия может возникать посредством слияния сперматозоида с яйцеклеткой и полярным тельцем либо при оплодотворении яйцеклетки двумя сперматозоидами. Естественная полиплоидия значительно чаще встречается в пустынях, высокогорьях, полярных областях (затруднено нормальное деление клеток). Предполагается, что на ранних этапах эволюции жизни полиплоидизации подвергались все организмы. Биологический смысл умножения количества генетического материала заключается в том, что происходит увеличение объема наследственной информации в клетке или создаются предпосылки для данного явления, посредством чего возрастают адаптивные способности организма. Явление полиплоидии отмечается у эукариотических микроорганизмов (грибов, водорослей), у растений, среди простейших. Полиплоидия всего организма у животных очень редка, хотя она часто встречается в некоторых дифференцированных тканях, например в печени млекопитающих, а также кишечнике, слюнных железах, мальпигиевых сосудах ряда насекомых. Генетическая несбалансированность у полиплоидов-животных, по мнению ряда авторов, возникает в силу того, что разные гены различно реагируют на дозирование: одни обладают разной степени кумулятивным эффектом, а другие – нет [5]. Вопрос №45Факторы, изменяющие генетическую структуру популяцииВ популяциях сельскохозяйственных животных постоянно изменяются частоты генов, что можно наблюдать при анализе смежных поколений. Такие изменения составляют суть генетической эволюции. Основные факторы эволюции: мутации, естественный и искусственный отбор, миграции, дрейф генов. Одна из основных причин генетической изменчивости в популяции – мутации. Спонтанные мутации каждого гена происходят с низкой частотой, однако общая частота мутаций всех генов, которые содержат популяции, очень велика. Мутации, возникающие в половых клетках родительского поколения, приводят к изменению генетической структуры у потомства. В популяции постоянной численности в отсутствие отбора большинство возникших мутаций быстро утрачивается, однако некоторые из них могут сохраниться в ряде поколений. Исчезновению мутантных генов из популяции противостоит действие мутационного процесса, в результате которого образуются повторные мутации. Генетическая структура популяций формируется и изменяется под действием естественного и искусственного отбора. Действие естественного отбора состоит в том, что преимущественное размножение имеют особи с высокой жизнеспособностью, скороспелостью, плодовитостью и т.д., т.е. более приспособленные к условиям окружающей среды. При искусственном отборе определяющее значение имеют признаки продуктивности. В.И. Власов отмечает, что естественный отбор идет на всех этапах онтогенеза популяции – от образования гамет до взрослого организма. При этом он существенно влияет на темпы искусственного отбора вследствие противоположного действия при селекции на высокий уровень развития продуктивных признаков, несвойственный видовым биологическим границам. Исходя из этого, при отборе животных необходимо учитывать не только продуктивные признаки, но и признаки приспособленности к условиям окружающей среды. По С.М. Гершензону, критерием интенсивности естественного отбора служит разность приспособленности сравниваемых групп, называемая коэффициентом отбора и выражаемая в долях единицы. Например, если вероятность оставления потомства особями с генотипом аа на 10% меньше, чем особями с генотипом АА или Аа, то приспособленность этих групп для особей АА и Аа равна 1, для особей аа – 0,9. С точки зрения ветеринарной генетики имеет значение эффективность отбора против вредных мутаций, прежде всего рецессивного типа. Анализ показывает, что высокие частоты рецессивного мутантного гена путем отбора могут быть быстро снижены до низких значений. Чтобы снизить частоту летального гена, например с 0,3 до 0,2, достаточно двух поколений. Частота гомозигот (аа) по мутантному гену зависит от частоты гетерозиготных животных в популяции. Выявление этих гетерозигот и элиминация их соответственно будут снижать частоту генетических аномалий, обусловленных мутантным геном, что особенно важно при высокой частоте мутаций [6]. Генетическая структура популяции может изменяться в силу случайных генетико-автоматических процессов (по Н.П. Дубинину) или дрейфа генов (по С. Райту). Наблюдения показывают, что наиболее интенсивно дрейф генов протекает в малых популяциях. Например, известны случаи высокой концентрации редких мутаций в малочисленных изолированных популяциях крупного рогатого скота и других видов животных, связанные, очевидно, с генетико-автоматическими процессами [3]. Распространение мутаций в разных популяциях животных может произойти в результате миграций. На практике это означает завоз животных, особенно производителей или их спермы, из других зон страны или стран. Известно много случаев, когда импортные производители оказывались носителями мутаций и распространяли генетические аномалии при использовании в воспроизводстве местных популяций. Так, экспорт голштинов из США в ФРГ, по данным Рика, способствовал распространению пупочных грыж у немецкого черно-пестрого скота. В нашей стране наблюдалась ситуация, когда из одного племзавода ремонтных быков с транслокацией 1/29 хромосомы продавали без ограничений в разные области. Это могло бы привести к распространению нежелательной мутации в других популяциях. По рекомендации ученых были приняты меры по элиминации носителей транслокаций. Спаривание животных, находящихся в родственных отношениях, называют инбридингом. Родственное спаривание, или инбридинг, – метод подбора, используемый в племенном животноводстве для закрепления ценных наследственных признаков того или иного животного в последующих поколениях. У родственных между собой животных наблюдается сходство по определенным парам аллелей, которые они получили от общего предка. Это сходство тем больше, чем ближе степень родства. Каждое животное в генотипе имеет аллельные гены, как в гомозиготном, так и в гетерозиготном состоянии. В гетерозиготе обычно находятся вредные мутантные рецессивные гены. При инбридинге возрастает вероятность слияния тождественных гамет, несущих мутантные гены в гетерозиготном состоянии, и перехода их в гомозиготное состояние. Эта вероятность пропорциональна степени родства спариваемых животных. Повышение гомозиготности у потомства при разных степенях родственных отношений родителей определяют по несколько измененной формуле С. Райта: 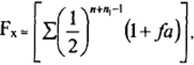 где Fх – коэффициент инбридинга потомка; n – ряд родословной с материнской; n1 – ряд родословной с отцовской стороны, где находится общий предок; fа – коэффициент инбридинга общего предка. Таким образом, в результате применения инбридинга происходит изменение генных частот, возрастает вероятность выщепления рецессивных гомозигот, что является причиной инбредной депрессии, выражающейся в снижении жизнеспособности, плодовитости животных, рождении аномальных особей. Инбредная депрессия по показателям, характеризующим продуктивность и жизнеспособность животных, не является фатальным спутником родственного спаривания. Имеется множество примеров того, когда при инбридинге разных степеней, в том числе и близких, отрицательных последствий не наблюдали. Н.П. Дубинин в этой связи отмечает, что «линия ухудшается, пока в ней идут процессы последовательного накопления вредных рецессивных генов, переходящих в гомозиготное состояние. Когда же наступает более или менее выраженное завершение этого процесса, линии по своим свойствам становятся относительно константными и могут в таком устойчивом состоянии сохраняться длительно. Изменить генотип таких линий могут лишь новые накапливающиеся в них мутации». В ходе длительной эволюции животных наряду с полезными мутациями, подхватываемыми отбором, в популяциях или породах накопился определенный спектр генных и хромосомных мутаций. Каждое поколение популяции наследует этот груз мутаций, и в каждом из них возникают новые мутации, часть которых передается последующим поколениям. Очевидно, что большая часть вредных мутаций отметается естественным отбором или элиминируется в процессе селекции. Это, прежде всего, доминантные генные мутации, фенотипически проявляющиеся в гетерозиготном состоянии, и количественные изменения наборов хромосом. Рецессивно действующие генные мутации в гетерозиготном состоянии и структурные перестройки хромосом формируют генетический груз популяции. Таким образом, под генетическим грузом популяции понимают совокупность вредных генных и хромосомных мутаций. Различают мутационный и сегрегационный генетический груз. Первый формируется вследствие новых мутаций, второй – в результате расщепления и перекомбинирования аллелей при скрещивании гетерозиготных носителей «старых» мутаций. Частота летальных, полулегальных и субвитальных мутантных генов, передающихся из поколения в поколение в форме мутационного генетического груза, из-за трудности идентификации носителей не поддается точному учету. Мортон и Кроу предложили форму расчета уровня генетического груза в количестве летальных эквивалентов. Один летальный эквивалент соответствует одному летальному гену, обусловливающему смертность с 100%-ной вероятностью, двум летальным генам при 50%-ной вероятности смерти и т.д. Величина генетического груза по формуле Мортона: log eS = А + BF, где S – часть потомства, оставшаяся в живых; А – смертность, измеряемая летальным эквивалентом в популяции при условии случайных спариваний (F=0), плюс смертность, обусловленная внешними факторами; В – ожидаемое увеличение смертности, когда популяция становится полностью гомозиготной (F= 1); F – коэффициент инбридинга. Уровень генетического груза можно определять на основании фенотипического проявления мутаций (уродства, врожденные аномалии обмена и т.д.), анализа типа их наследования, частоты в популяции. Н.П. Дубинин предлагает определять генетический груз популяции путем сравнения частот мертворожденных в родственных и неродственных подборах родительских пар. При этом следует иметь в виду, что при высокой частоте гетерозигот по рецессивным летальным и полулегальным мутантным генам рождение животных с аномалиями необязательно должно быть связано с инбридингом близких и умеренных степеней. Хромосомные мутации являются составной частью генетического груза. Учет их ведется прямым цитологическим методом. По результатам многочисленных исследований основной компонентой груза аберраций хромосом у крупного рогатого скота являются робертсоновские транслокации, а у свиней – реципрокные. Наиболее распространенной мутацией у крупного рогатого скота оказалась транслокация 1/29 хромосомы. Размах изменчивости частоты этой аберрации в популяциях палево-пестрого скота составлял от 5 до 26%. Таким образом, концепция генетического груза в свете современных достижений цитогенетики должна быть расширена. Сейчас, когда известен широкий спектр аберраций хромосом и установлено строгое наследование отдельных из них (транслокации и инверсии), представляется целесообразным учитывать их наряду с вредными мутациями генов как составляющую часть генетического груза [6]. Вопрос №70У кур оперенные ноги (О) доминируют над голыми (о), а гороховидный гребень (Р) – над простым (р). Петух с оперенными ногами и гороховидным гребнем, скрещенный с голоногой курицей, имеющий также гороховидный гребень, дал потомство с оперёнными ногами. При этом большинство потомков имело гороховидный гребень, но встречались куры и с простым гребнем. Каковы генотипы родителей? Какое потомство дала бы эта курица при скрещивании с одним из своих сыновей, имеющим оперенные ноги и простой гребень? Дано: петух курица О – оперенные ноги; Р: ООРр × ооРр о – голые ноги; G: ОР, Ор; оР, ор Р – гороховидный гребень; р – простой гребень. F1: ОоРР, ОоРр, ОоРр, Оорр В итоге получилось, что: 75% потомства F1 имеет оперенные ноги и гороховидный гребень, а 25% потомства имеет оперенные ноги и простой гребень. Ответ: генотип курицы – ооРр, петуха – ООРр. Дано: петух курица О – оперенные ноги; Р: Оорр × ооРр о – голые ноги; G: Ор, ор; оР, ор Р – гороховидный гребень; р – простой гребень. F2: ОоРр, Оорр, ооРр, оорр Ответ: генотип курицы – ооРр, петуха – Оорр. В итоге получилось, что: у 25% потомства F2 оперенные ноги и гороховидный гребень, у 25% – оперенные ноги и простой гребень, у 25% – голые ноги и гороховидный гребень, у 25% – голые ноги и простой гребень. Вопрос №94Скорость оперения у цыплят является признаком, сцепленным с полом. Доминантный аллель (К) обуславливает медленное формирование перьев, а рецессивный аллель (k) – быстро формирующееся оперение. Скорость оперения цыплят можно узнать сразу же после выклева, поэтому этот признак используют для раннего разделения по полу. Какие генотипы должны иметь петух и курица, чтобы их потомство можно было разделить по полу? Дано: К – медленно формирующееся оперение; k – быстро формирующееся оперение. Решение: Р: ♀ХКY × ♂ХКXk G: ХК, Y; ХК, Xk F1: ХКХК – петух с медленно формирующимся оперением; ХКY – курица с медленно формирующимся оперением; ХКXk – петух с быстро формирующимся оперением; XkY – курица с быстро формирующимся оперением. Ответ: генотип петуха – ХКXk, генотип курицы – ХКY. СЛОВАРЬ ТЕРМИНОВАллельные гены – разные формы одного гена, возникшие в результате мутации и располагающиеся в одинаковых участках (локусах) гомологичных хромосом. Гамета – зрелая женская или мужская половая клетка (яйцеклетка или сперматозоид) с гаплоидным набором хромосом. Генотип – совокупность генов данного организма. Гетерозигота – особь, содержащая в соматических клетках разные аллели данного гена (например, Аа). Гомозигота – особь, содержащая в соматических клетках одинаковые аллели данного гена (например, АА / аа). Доминирование – проявление действия одного из аллелей у гетерозиготного организма. Доминирование неполное – внутриаллельное взаимодействие, при котором доминантный ген не полностью подавляет действие рецессивного гена (промежуточное наследование). Дрейф генов – случайные колебания частот генов в малых популяциях (генетико-автоматические процессы). Кодоминирование – аллельное взаимодействия генов, при котором у гетерозиготного организма фенотипически проявляются оба аллеля. Полиплоидия – увеличение числа хромосом, кратное гаплоидному набору [7]. СПИСОК ЛИТЕРАТУРЫ1. Гайсинович А.Е. Зарождение и развитие генетики. – М.: Наука, 1988. – 424 с. 2. Дубинин Н.П. Общая генетика. – М.: «Наука», 1986. – 560 с. 3. Иванов В.И. Генетика / И.В. Иванов, Н.В. Барышникова, Дж.С. Билева. – М.: Академкнига, 2007. – 638 с. 4. Кочнева М.Л. Цитогенетика: Учебное пособие, 3-е издание доп. – Новосибирск: Изд-во НГАУ, 2017. – 115 с. 5. Петухов В Л. Генетика / В.Л. Петухов, О.С. Короткевич, С.Ж. Стамбеков и др. – Новосибирск: СемГПИ, 2007. – 628 с. 6. Тарантул В.З. Толковый биотехнологический словарь. Русско-английский. – М.: Языки славянских культур, 2009. – 936 с. 7. Nissani M. Psychological, Historical, and Ethical Reflections on the Mendelian Paradox // Perspectives in Biology and Medicine. – 1994. – Т. 37, № 2. – 547 р. |