КУРС ЛЕКЦИЙ ПО ФИЗИОЛОГИИ. Курс лекций по нормальной физиологии. Ю. И. Савченков. Красноярск Издво , 2012, 470 с
 Скачать 8.61 Mb. Скачать 8.61 Mb.
|

23. 2. Транспорт О2 и СО2 кровью. Газообмен между кровью и тканями.Связывание кислорода гемоглобином. Транспорт О2 из альвеол в кровь и транспорт СО2 из крови в альвеолы осуществляется с помощью диффузии. Транспорт газов осуществляется в физически растворенном и химически связанном виде. Физические процессы, т. е. растворение газа, не могут обеспечить запросы организма в О2. Подсчитано, что физически растворенный О2 может поддерживать нормальное потребление О2 в организме (250 мл/мин), если минутный объем кровообращения составит примерно 83 л/мин в покое. Наиболее оптимальным является механизм транспорта О2 в химически связанном виде. В количественном отношении формы транспортируемого газа значительно различаются, так как количество растворенного физически газа невелико. Однако следует отметить, что хотя количество физически растворенных О2 и СО2 невелико, эта часть газов крови играет огромную роль в жизнедеятельности организма. Дело в том, что прежде чем связаться с определенными веществами крови газы должны быть доставлены к ним в физически растворенном состоянии. Содержание физически растворенного газа в жидкости зависит от парциального давления (или напряжения в жидкости) и коэффициента растворимости. В артериальной крови при 37 °С содержание физически растворенного кислорода составляет 0,3 мл на 100 мл крови, а углекислого газа – 2,6 мл на 100 мл крови. Гемоглобин (Нb) способен избирательно связывать О2 и образовывать оксигемоглобин (НbО2) в зоне высокой концентрации О2 в легких и освобождать молекулярный О2 в области пониженного содержания О2 в тканях. При этом свойства гемоглобина не изменяются и он может выполнять свою функцию на протяжении длительного времени. Г 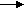 емоглобин переносит О2 от легких к тканям. Эта функция зависит от двух свойств гемоглобина: 1) способности изменяться от восстановленной формы, которая называется дезоксигемоглобином, до окисленной (Нb + О2 НbО2) с высокой скоростью (полупериод 0,01 с и менее) при нормальном РО2 в альвеолярном воздухе; 2) способности отдавать О2 в тканях (НbО2 Нb + О2) в зависимости от метаболических потребностей клеток организма. емоглобин переносит О2 от легких к тканям. Эта функция зависит от двух свойств гемоглобина: 1) способности изменяться от восстановленной формы, которая называется дезоксигемоглобином, до окисленной (Нb + О2 НbО2) с высокой скоростью (полупериод 0,01 с и менее) при нормальном РО2 в альвеолярном воздухе; 2) способности отдавать О2 в тканях (НbО2 Нb + О2) в зависимости от метаболических потребностей клеток организма.Б ольшая часть О2 переноситcя в виде химического соединения с гемоглобином – HbO2. Это видно из того, что в цельной артериальной крови в покое находится 20 мл О2 на 100 мл крови. Поскольку молекула Hb состоит из четырех субъединиц, а каждая из них связывает по одной молекуле О2, то одна молекула кислорода связывает 4 моля О2Следовательно, при молекулярной массе гемоглобина в 64 500 1 г гемоглобина связывает 1,39 мл О2. Фактически эта величина несколько меньше, так как часть молекул гемоглобина находится в неактивном виде и составляет 1,34–1,36 мл. Кривая диссоциации оксигемоглобина. Реакция, отражающая присоединение кислорода к гемоглобину подчиняется закону действующих масс. Это означает, что соотношение между Hb и HbO2 зависит от содержания физически растворенного кислорода. Отношение количества оксигемоглобина к общему количеству гемоглобина (в %) в крови называется насыщением гемоглобина кислородом Если гемоглобин полностью дезоксигенирован, то насыщение составляет 0%, если гемоглобин полностью насыщен кислородом, то насыщение составляет 100%. Графическая зависимость насыщения гемоглобина кислородом от напряжения О2 называется кривой диссоциации оксигемоглобина. Эта кривая имеет S-образную форму (Рис. 43). Такая форма имеет большой физиологический смысл. В области высокого напряжения кислорода, составляющего в артериальной крови около 95 мм рт. ст. (молодые люди в покое), насыщение составляет 97%. 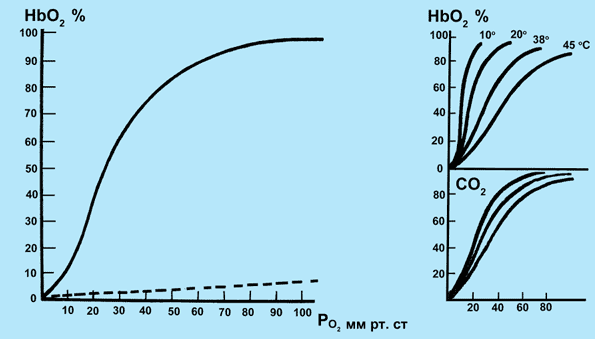 Рис. 42. Кривая диссоциации гемоглобина. Справа вверху - влияние температуры на кривую диссоциации В этой области максимального насыщения степень насыщения мало зависит от напряжения кислорода. Поэтому насыщенность гемоглобина кислородом сохраняется на высоком уровне даже при существенных сдвигах в напряжении кислорода. Крутой наклон кривой диссоциации означает, что в области средних и малых концентраций кислорода, даже небольшие изменения в содержании кислорода приводят к существенной отдаче его. Это облегчает отдачу кислорода тканям. В состоянии покоя в области венозного конца капилляра напряжение О2 примерно равно 40 мм рт. ст., что соответствует 73% насыщению. Если в результате потребления кислорода его напряжение снизится всего на 5 мм рт. ст., то насыщение гемоглобина кислородом уменьшится на 7%. Высвобождающийся О2 может быть сразу же использован для процессов окисления. Метаболические факторы являются основными регуляторами связывания О2 с гемоглобином в капиллярах легких, когда уровень O2, рН и СО2 в крови повышает сродство гемоглобина к О2 по ходу легочных капилляров. В условиях тканей организма эти же факторы метаболизма понижают сродство гемоглобина к О2 и способствуют переходу оксигемоглобина в его восстановленную форму — дезоксигемоглобин. В результате О2 по концентрационному градиенту поступает из крови тканевых капилляров в ткани организма. Артерио-венозная разница по О2. Так как содержание кислорода в артериальной крови составляет 0,2 л на 1 л крови, а в венозной – 0,15 л, то артерио-венозная разница достигает 0,05 л О2 на 1 л крови. Следовательно, в норме при прохождении крови через капилляры используется лишь 25% кислорода. Разумеется, это средняя величина. Она неодинакова в разных органах и тканях и зависит от функционального состояния организма, т.е. интенсивности метаболизма. Факторы, влияющие на кривую диссоциации оксигемоглобина. Кривая диссоциации обусловлена, главным образом, химическими свойствами гемоглобина. В то же время существует ряд факторов, влияющих на наклон этой кривой, но не изменяющих при этом ее S-образный характер. К таким факторам относят температуру, рН, напряжение СО2 и некоторые другие. При понижении температуры наклон кривой возрастает, при повышении температуры наклон кривой уменьшается. У теплокровных этот эффект проявляется только при гипотермии и при лихорадочной реакции. При закислении среды сродство гемоглобина к О2 уменьшается, так как кривая уплощается. Этот эффект носит название эффекта Бора. Величина кислотности крови тесно связана с содержанием СО2. Поэтому понятно, что при повышении напряжения СО2 рН снижается и это вызывает уплощение кривой, т.е. снижение сродства гемоглобина к кислороду. Эффект Бора имеет биологический смысл, так как способствует отдаче кислорода там, где выше интенсивность метаболизма, например в работающих мышцах. Под кислородной емкостью крови понимают количество О2, которое связывается кровью до полного насыщения гемоглобина. При содержании гемоглобина в крови 8,7 ммоль/л кислородная емкость крови составляет 0,19 мл О2 в 1 мл крови (температура 0oC и барометрическое давление 760 мм рт.ст., или 101,3 кПа). Величину кислородной емкости крови определяет количество гемоглобина, 1 г которого связывает 1,36—1,34 мл О2. Кровь человека содержит около 700—800 г гемоглобина и может связать таким образом почти 1 л О2. Физически растворенного в 1 мл плазмы крови О2 очень мало (около 0,003 мл), что не может обеспечить кислородный запрос тканей. Растворимость О2 в плазме крови равна 0,225 мл/л/кПа-1. С другой стороны, известно, что при напряжении О2 в артериальной крови капилляров, равном 100 мм рт.ст. (13,3 кПа), на мембранах клеток, находящихся между капиллярами, эта величина не превышает 20 мм рт.ст. (2,7 кПа), а в митохондриях равна в среднем 0,5 мм рт.ст. (0,06 кПа). Обмен О2 между кровью капилляров и клетками тканей также осуществляется путем диффузии. Концентрационный градиент О2 между артериальной кровью (100 мм рт.ст., или 13,3 кПа) и тканями (около 40 мм рт.ст., или 5,3 кПа) равен в среднем 60 мм рт.ст. (8,0 кПа). Изменение градиента может быть обусловлено как содержанием О2 в артериальной крови, так и коэффициентом утилизации О2, который составляет в среднем для организма 30— 40%. Коэффициентом утилизации кислорода называется количество О2, отданного при прохождении крови через тканевые капилляры, отнесенное к кислородной емкости крови. Перенос углекислого газа. Поступление СО2 в легких из крови в альвеолы обеспечивается из следующих источников: 1) из СО2, растворенного в плазме крови (5—10%); 2) из гидрокарбонатов (80—90%); 3) из карбаминовых соединений эритроцитов (5—15%), которые способны диссоциировать. Для СО2 коэффициент растворимости в мембранах аэрогематического барьера больше, чем для О2, и составляет в среднем 0,231 ммоль/л-1/кПа-1 поэтому СО2 диффундирует быстрее, чем O2. Это положение является верным только для диффузии молекулярного СО2. Большая часть СО2 транспортируется в организме в связанном состоянии в виде гидрокарбонатов и карбаминовых соединений, что увеличивает время обмена СО2, затрачиваемое на диссоциацию этих соединений. Эндотелий капилляров проницаем только для молекулярного СО2 как полярной молекулы (О — С — О). Из крови в альвеолы диффундирует физически растворенный в плазме крови молекулярный СО2. Кроме того, в альвеолы легких диффундирует СО2, который высвобождается из карбаминовых соединений эритроцитов благодаря реакции окисления гемоглобина в капиллярах легкого, а также из гидрокарбонатов плазмы крови в результате их быстрой диссоциации с помощью фермента карбоангидразы, содержащейся в эритроцитах. Углекислый газ, также как и кислород транспортируется в форме физически растворенного и химически связанного газа. Физически растворено 10% газа от общего количества, 10% образует карбаминовую связь с гемоглобином, 35% транспортируется в виде бикарбонатов в эритроците, 45% – в виде бикарбонатов в плазме. Д 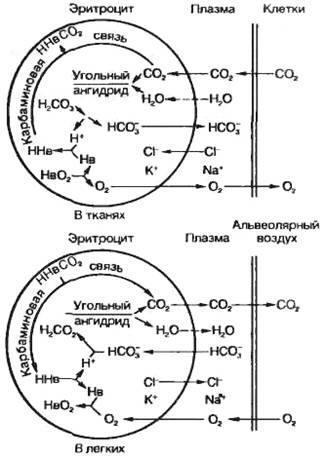 иффузия СО2 из тканей в кровь. Обмен СО2 между клетками тканей с кровью тканевых капилляров осуществляется с помощью следующих реакций: 1) обмена С1- и НСО3- через мембрану эритроцита; 2) образования угольной кислоты из гидрокарбонатов; 3) диссоциации угольной кислоты и гидрокарбонатов (рис. 43). иффузия СО2 из тканей в кровь. Обмен СО2 между клетками тканей с кровью тканевых капилляров осуществляется с помощью следующих реакций: 1) обмена С1- и НСО3- через мембрану эритроцита; 2) образования угольной кислоты из гидрокарбонатов; 3) диссоциации угольной кислоты и гидрокарбонатов (рис. 43).Рис. 43. Участие эритроцитов в обмене О2 и СО2 в тканях и в легких Химические реакции связывания СО2 сложнее, чем связывание О2. Это вызвано тем, что механизмы, ответственные за транспорт СО2, должны одновременно обеспечивать поддержание кислотно-основного равновесия и тем самым гомеостаза организма в целом. Углекислый газ, поступивший по градиенту напряжения из тканей в капилляры, в небольшом количестве остается в форме физического растворенного газа, остальная часть химически связывается. Прежде всего, гидратируется СО2: СО2 + Н2О → Н2СО3. В плазме эта реакция протекает медленно, а в эритроцитах в 10 000 раз быстрее благодаря присутствии в них фермента карбоангидразы. Затем угольная кислота, будучи слабой и нестойкой диссоциирует; Н2СО3 → НСО3– + Н+. Ионы НСО3– выходят в плазму, а вместо них в эритроцит поступают анионы хлора – так сохраняется электрический заряд клетки. По мере поступления СО2 образуются в эритроцитах и Н+. Однако это не приводит к сдвигу рН, так как оксигемоглобин более слабая кислота, чем угольная и может связать большее количество ионов Н+. В ходе газообмена СО2 между тканями и кровью содержание НСОз- в эритроците повышается и они начинают диффундировать в кровь. Для поддержания электронейтральности в эритроциты начнут поступать из плазмы дополнительно ионы С1-. Наибольшее количество бикарбонатов плазмы крови образуется при участии карбоангидразы эритроцитов. Углекислый газ связывается также непосредственно аминогруппами гемоглобина, образуя карбаминогемоглобин. При поступлении крови в капилляры легких эти реакции протекают в обратном направлении и СО2 диффундирует в альвеолы. Карбаминовый комплекс СО2 с гемоглобином образуется в результате реакции СО2 с радикалом NH2 глобина. Эта реакция протекает без участия какого-либо фермента, т. е. она не нуждается в катализе. Реакция СО2 с Нb приводит, во-первых, к высвобождению Н+; во-вторых, в ходе образования карбаминовых комплексов снижается сродство Нb к О2. Эффект сходен с действием низкого рН. Как известно, в тканях низкое рН потенцирует высвобождение О2 из оксигемоглобина при высокой концентрации СО2 (эффект Бора). С другой стороны, связывание О2 гемоглобином снижает сродство его аминогрупп к СО2 (эффект Холдена). Зависимость содержания СО2 в крови от его напряжения, выраженная графически, называется кривой связывания СО2. Между кривыми для связывания СО2 и кривой диссоциации оксигемоглобина существует принципиальная разница. Она состоит в том, что для СО2- кривая не имеет плато насыщения. Процесс выведения СО2 из крови в альвеолы легкого менее лимитирован, чем оксигенация крови. Это обусловлено тем, что молекулярный СО2 легче проникает через биологические мембраны, чем О2. По этой причине он легко проникает из тканей в кровь. К тому же карбоангидраза способствует образованию гидрокарбоната. Яды, которые ограничивают транспорт О2 (такие как СО, метгемоглобинобразующие субстанции — нитриты, метиленовый синий, ферроцианиды и др.) не действуют на транспорт СО2. Блокаторы карбоангидразы, например диакарб, которые используются нередко в клинической практике или для профилактики горной или высотной болезни, полностью никогда не нарушают образование молекулярного СО2. Наконец, ткани обладают большой буферной емкостью, но не защищены от дефицита О2. По этой причине нарушение транспорта О2 наступает в организме гораздо чаще и быстрее, чем нарушения газообмена СО2. Тем не менее при некоторых заболеваниях высокое содержание СО2 и ацидоз могут быть причиной смерти. |