биология. Курсовая работа Основные гидробиомы Исполнитель студентка группы гэ22 Д. В. Котюжинская
 Скачать 2.8 Mb. Скачать 2.8 Mb.
|

|
Как было выше сказано, что совокупность различных групп организмов и среды их обитания в определенных широтной и вертикальной зонах называют биомом. Так, и в Мировом океане, рассматривая его как отдельный биом, выделяются зоны и подзоны [2]. На рисунке 1 видны широтные зоны Мирового океана, которые обозначаются следующим образом: ВА - высокоарктическая и НА - низко-арктическая подзоны арктической зоны; ВБ - высокобореальная и НБ - низкобореальная подзоны бореальной зоны; ССТ - северная субтропическая зона: СЦ - североцентральная, Э - экваториальная, ЮЦ - южно-центральная подзоны тропической зоны; ЮСТ - южная субтропическая зона; Н - нотальная зона; НАн - низко-антарктическая. ВАн - высокоантарктическая подзоны антарктической зоны. Переходные полосы вертикально заштрихованы. Прерывистые линии - границы подзон арктической и антарктической зон. Переходная полоса между южной субтропической и тотальной зонами косо заштрихована. 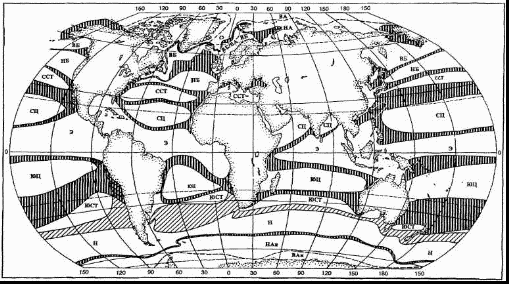 Рисунок 1 - Широтные зоны пелагиали Мирового океана [3] Пелагиаль делится на следующие зоны: эпипелагиаль, или слой воды покрывает материковую отмель (глубина 0-200 м), батипелагиаль, или толща воды над материковым склоном, и абисопелагиаль - толща воды над океаническим ложем. Эпипелагиаль - наиболее продуктивная зона морских экосистем. В ее верхней части наиболее интенсивно протекают биологические процессы (фотосинтез автотрофных организмов - фитопланктона) и создается первичная органическая продукция используется животными организмами, которые живут в батипелагиали, абисопелагиали и бентали. На глубине 2,5 - 3 км, у подземных термальных вод, органические вещества могут образовываться также в результате хемосинтеза, осуществляемой хемотрофных бактериями. Крупномасштабные циркуляционные процессы в экосистеме океана формируют своеобразный химический, гидрологический и гидробиологический режим его отдельных зон. Каждая из них характеризуется определенными условиями, к которым приспосабливаются морские организмы. Циркуляция водных масс определяет распределение взвешенных и растворенных веществ абиотической и биотического происхождения как по акватории, так и по глубине морей и океанов. Под водной массой понимают не всю воду моря или океана, а лишь определенный, небольшой ее объем, связанный с конкретным районом Мирового океана. Если же учесть, что в океане вследствие температурной и солевой разницы формируются зоны с неоднородной плотностью воды и резко ограничивается турбулентный обмен между ними, то станет понятной возможность существования среди просторов океана отдельных водных масс. В рамках больших водных масс могут формироваться водные массы не только первого, но и второго, третьего и следующего порядков, объем которых меньше, а свойства не так резко отличаются. Как правило, первичные водные массы привязаны к определенным климатическим районов океана, а вторичные образуются на границе двух соседних первичных, или в районах проникновения в океан водных масс из близлежащих морей. Циркуляционные воды, при этом формируются, быстрее обновляются, их свойства не такие постоянные, как свойства первичных водных масс. Общая циркуляция водных масс в Мировом океане происходит вследствие ветровой напряжения на поверхности воды, неравномерного распределения атмосферного давления и температурного режима отдельных его зон. В шельфовой зоне морей и океанов вследствие небольших глубин и соседства суши формируются наиболее благоприятные условия для развития жизни (высокой биологической продуктивности) [4]. Формула гидробиома выглядит следующим образом: термические условия + динамика водных масс (интенсивность водообмена, скорость и характер течений) + соотношение окислительных и восстановительных условий + субстрат + ключевые жизненные формы биоты (экоморфы, ценоэкоморфы) + трофическая структура (источники энергии, ключевые трофические связи) + хорология (границы, ярусность, стратификация, мозаичность) или Т+Д+ОВ+С+ЖФ+Тр+Х. Исходя из этой формулы могут быть представлены следующие основные характеристики гидробиомов [2]. Основные характеристики отображены в таблице 1. Таблица 1 - Основные характеристики гидробиомов [2]
Основными характеристиками среды в пелагическом океаническом биоме являются свойства водных масс океана: замедленный в целом водообмен (относительно водообмена поверхностных вод суши), существование фронтов и разномасштабных циркуляций, разделение на относительно малую по объему фотическую, изменчивую по температуре и большую по объему дисфотическую и достаточно стабильную по температуре холодную зоны. Однако, эти характеристики могут быть использованы для деления биома на меньшие категории, и не влияют на целостность всего биома (мегабиома). Характеристика жизненных форм океанического пелагического биома может быть образно охарактеризована так: его населяют организмы, которые «не знают» и «не подозревают» о существовании границ вода - берег, вода - дно. Переходя эту границу, например, переходя из стадии пелагической личинки к седентарной стадии взрослой особи, организмы попадают в другие экосистемы и в другой биом и представляют собой уже совершенно иные жизненные формы и экоморфы. Пелагический мегабиом океана может быть разделен на два макробиома - по характеру освещенности и, соответственно, с различными трофо-энергетическими характеристиками с автотрофно-гетеротрофными (фотическая зона) и гетеротирофными (афотическая) сообществами. Прибрежный или циркумконтинентальный биом океана ограничен береговой линией и шельфовой зоной. Здесь тесное взаимодействие водной толщи, её поверхности, дна, граничной области море-суша создают чрезвычайно большое биотопическое разнообразие. Это является предпосылкой существования здесь многих жизненных форм гидробионтов - седентарных и подвижных, с разнообразными типами питания. Значительная часть экосистем этого биома автотрофно-гетеротрофные, высоко продуктивны. В прибрежном мегабиоме хорошо выражена широтная зональность, что позволяет выделять биогеографические прибрежные шельфовые области с различными условиями обитания и различающимся населением. Тесное взаимодействие с биомами суши проявляется в эвтуарных зонах. Особенности генезиса, состава жизненных форм, метаболических процессов позволяют выделить биогермовый биом, хотя пространственно он частично совпадает с прибрежным. Биогермовым биомом представляет собой комплекс экосистем, в которых живыми организмами создаются структуры, сопоставимые с геоморфологическими, которые являются топической основой существования экосистем . Хотя в настоящее время этот биом представлен в основном коралловыми экосистемами, начиная с докембрийских времен не менее полутора десятков групп организмов (от цианобактерий до мшанок) участвовали в формировании биогермовых экосистем. Батиально-абиссальный биом океана представляет собой совокупность донных экосистем за пределами шельфа. Топически, видимо, целесообразно выделить макробиомы - срединно-океанических хребтов, донных осадков океанического ложа, глубоководных желобов ультраабиссали. Основной особенностью этих экосистем является их полная энергетическая зависимость от процессов первичного продуцирования в фотической зоне океана. Именно поэтому, как отмечал Г.В. Богоров минимальным изопланктам в поверхностной зоне соответствуют минимальные изобенты. Основными жизненными формами гидробионтов в этом биоме являются седентарные и подвижные с опорой на донный субстрат формы. Деление как пелагического, так и донного океанического биомов по экологическим критериям совпадает в основных чертах с фаунистическим. В пелагическом биоме можно выделить мелководную (продуцирующую, 0 - 200 м), переходную - 200 - 1000 м - и глубоководную, потребляющую области. Гидротермально-сиповый биом можно назвать также биомом восстановительных условий. Основным абиотическим фактором выступает не столько высокая температура гидротермальных флюидов, сколько локальное преобладание восстановительных условий над окислительными. Особенностью этого биома как совокупности своеобразных экосистем является их островное расположение. Трофическая структура этих сообществ базируется на функционировании хемоавтотрофных организмов и значительную роль играют симбиотрофические связи. Все экосистемы поверхностных вод суши при наиболее общем подходе могут быть разделены на две большие группы: лимнобиом - совокупность экосистем водоемов с замедленным водообменом и реобиом или совокупность экосистем текучих вод и. На уровне отдельных экосистем бывает сложно обособить экосистемы водоемов и водотоков. Лимнобиом в масштабах континентов имеет мозаичную, островную природу. Водоемы с замедленным водообменом более или менее ограничены друг от друга, объединяются водотоками. Основной градиент условий проходит в них от литорали в глубину к профундальной части. В литоральной области преобладают седентарные формы, в том числе высшие водные растения. Водотокам свойственна структура последовательных, связанных по их протяженности экосистем, обобщенный взгляд на которую представлен в концепции речного континуума . Бывает практически невозможно выделение и разграничение отдельных малых экосистем, поэтому предлагается введение понятия реобиом: "надо говорить о существовании особой надэкосистемной формы организации природных комплексов - континууме речных экосистем". Очевидно, что такой подход может быть распространен не только на экосистему конкретной реки, но на всю речную сеть данного бассейна. В то же время, по основным признакам - общему характеру условий среды, ведущих жизненным форм гидробионтов, организации речного континуума все экосистемы водотоков в отличие от других гидроэкоситем могут быть объединены в единый мегареобиом гидросферы. Таким образом микро- и мезореобиомы представляют собой топо-хорологические единицы, а макрореобиом - как типологические, с позиций концептульного укрупнения понятий, так и хорологические - с позиций структуры биогидросферы [2]. Зона обитания водных животных, не связанных с дном водоема. Разделяется на зоны по вертикали и горизонтали (рисунок 2). гидробиом пелагиаль океан 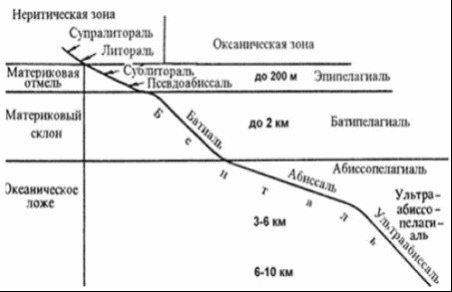 Рисунок 2 - Экологические зоны Мирового океана [2] Разделение по вертикали (на слои, лежащие друг над другом): эпипелагиаль - верхний слой, покрывающий материковую отмель, воды до глубины 200 м, нижняя граница сублиторали; батипелагиаль - глубже лежащий слой до нижней границы батиали, толща воды над материковым склоном; абиссопелагиаль - располагается от нижней границы батиали до глубин 6 - 7 км; ультраабиссопелагиаль. 1.1 Эпипелагиаль Эпипелагиаль - верхний слой воды моря до глубины 200 м (нижняя граница сублиторали) [5]. Для эпипелагиали особенно характерны рыбы крупного размера, относящиеся главным образом к нектону, способные к быстрому продолжительному плаванию и ведущие активный образ жизни, - именно здесь обитают рыбы-рекордисты по длине и массе тела (китовая акула - 15,2 м и 14 т), скорости плавания (меч-рыба - 130 км/ч) и его дальности (синий тунец - 5800 миль, т. е. 10 тыс. км). Планктонных рыб в этом биотопе немного, но и среди них имеются настоящие гиганты (луна-рыба «диаметром» 3 м и массой 1,5 т). Почти все обитатели эпипелагиали имеют широкие пищевые спектры, быстро растут, рано достигают половой зрелости, имеют сравнительно непродолжительную жизнь и большую индивидуальную плодовитость при отсутствии заботы о потомстве. По внешнему облику, образу жизни и экологическим характеристикам они резко отличаются от рыб глубоководной пелагиали [6]. На рисунке 3 изображены основные группировки рыб, такие как голоэпипелагическая (ГЭ), мероэпипелагическая (МЭ), ксеноэпипелагическая (КЭ).  Рисунок 3 - Основные группировки рыб эпипелагиали [6] Рыбы эпипелагиали, как правило, отличные, быстрые пловцы с обтекаемой формой тела. Для них характерна синяя или фиолетово-синяя окраска спины и серебристо-белое брюхо. Такая окраска имеет защитное значение - тело рыбы плохо различимо как при взгляде сверху, на темно-синюю поверхность океана, так и при взгляде снизу, на ярко освещенную солнцем его поверхность. Однако есть и исключения. В эпипелагиали Тихого океана живут малоподвижные рыбы, обитающие среди скоплений плавающих водорослей и других предметов, занесенных течениями в океан, т. е. всего того, что называется «плавником». Эти рыбы обладают причудливой формой тела, приспособленной к существованию среди переплетения водорослей, и преимущественно имеют зеленую или коричневую окраску с темными полосами и пятнами. Таковы морская мышь и рыба-игла. В эпипелагиали открытого океана держатся также взрослые стадии рыб, но для размножения подходящие к берегам материков и иногда даже входящие в реки. Таковы прежде всего лососи - кета, горбуша, камчатская семга, голец, куиджа, нерка и некоторые другие виды. Область морского распространения этих рыб в Тихом океане охватывает ту часть его акватории, которая лежит к северу от течения Куросио и Северо-Тихоокеанского течения, т. е. примерно от 38° с. ш. Период пребывания мальков в пресной воде рек длится несколько лет, и только мальки кеты, горбуши и от части нерки и чавычи выплывают в открытый океан в первое же лето после выклева из икры. Именно поэтому кета и горбуша отличаются наиболее «морскими» чертами биологии. Эти виды раньше других освоили пищевые ресурсы моря и более других размножились и распространились в Тихом океане. Область нереста кеты и горбуши охватывает все побережье Азии, от о-ва Хонсю (Япония) и южного Приморья до Берингова пролива, и отсюда простирается на юг, вдоль американского побережья до залива Монтерей. Сходный нерестовый ареал свойствен нерке, кижучу и чавыче, однако эти рыбы, в отличие от кеты и горбуши, в большей степени тяготеют к американскому побережью. Напротив, семга обладает привязанностью только к азиатскому побережью Тихого океана. Взрослые лососевые рыбы к наступлению икрометания объединяются в огромные стаи и совершают длительные переходы к устьям тех рек, из которых они мальками выплыли в открытый океан. И опять-таки лишь у кеты и горбуши существуют нерестовые миграции из океана не в ту самую реку, которую они покинули мальками, а в другую, но лежащую в пределах одного и того же района побережья. Обратный путь к рекам достигает иногда многих тысяч километров. Примечательно, что во время перехода в реках к местам нереста рыбы перестают питаться и существуют лишь за счет накопленного жира. Биологический смысл такого поведения вполне понятен - река не смогла бы прокормить огромные стаи рыб. После икрометания тихоокеанские лососи погибают. По данным, северная часть Тихого океана бедна настоящими эпипелагическими рыбами открытых вод. По сути дела сюда лишь для нагула проникают те формы, которые характерны для умеренных тихоокеанских вод. Здесь можно встретить сельдевую акулу, кинжалозуба, алепизавра, морского леща. Эти рыбы размножаются в южнобореальных водах Тихого океана. Для нагула сюда заходят питающаяся планктоном и совершенно безопасная для человека гигантская акула и свирепый хищник - голубая акула. Последняя потенциально опасна для человека. Однако, будучи голоэпипелагической рыбой, она крайне редко подходит к берегам, где встреча с купающимися или терпящими бедствие людьми наиболее вероятна. В открытом океане если такие встречи и случаются, то океан хранит тайну гибели случайно упавших за борт людей и жертв кораблекрушений. Из костистых рыб в южнобореальных водах Тихого океана держатся сайра - наиболее характерный компонент эпипелагического промысла на Дальнем Востоке, длинноперый тунец, полосатый тунец, морской колючий лещ, кубохвосты и некоторые другие рыбы. Заходит в эту зону и меч-рыба. Кроме меч-рыбы и тропических тунцов все они размножаются южнее, в северной периферической части тропического пояса Тихого океана. Здесь же размножаются скумбриещуки и полосатый марлин. В периферической части тропической зоны имеется и свой собственный набор эпипелагических рыб, не встречающихся в остальной тропической части океана. Непременным его компонентом служат карликовая акула и летучие рыбы, относящиеся к отряду сарганообразных рыб. Сарганообразные рыбы обитают только в поверхностных водах океана, и у большинства из них в процессе эволюции выработалась способность, спасаясь от хищников, совершать прыжки над водой. Но у летучих рыб и летучих полурылов этот процесс пошел еще дальше и простой прыжок превратился в планирующий полет, менее совершенный у полурылов и достигающий значительной дальности у летучих рыб. Самые мелкие китообразные - дельфиновые. Их размеры колеблются у разных видов от 1 до 10 м, но большинство дельфинов имеет 2 - 3 м в длину. Дельфины обладают стройным телосложением и очень подвижны. Они любят играть возле проходящих судов и, «оседлав» волну у носа корабля, способны без особого труда развивать скорость более 60 км/час. Почти все дельфины хорошо уживаются в океанариях, очень умны и легко дрессируются (возможно, что это справедливо и в отношении других китообразных, просто еще не созданы океанарии для самых крупных из них). Дельфины владеют сложной звуковой сигнализацией в способны издавать и улавливать звуки в очень широком диапазоне, а афалины могут подражать и человеческому голосу. Как и все зубатые киты, они владеют и эхолокацией, при помощи которой ориентируются в воде, обнаруживают и успешно добывают пищу - рыбу и кальмаров. Больше того, основные представители голоэпипелагической ихтиофауны тропической зоны встречаются как в Тихом, так и в Атлантическом и Индийском океанах, что объясняется высокой подвижностью нектонных животных и их способностью совершать длительные миграции и подолгу находиться для нагула в водах с более низкой температурой, чем в тропиках. 1.2 Батипелагиаль Экологическая зона открытой части моря (водная толща), соответствующая материковому склону называется батипелагиаль. Зона расположена на глубине от 200 м до 4000 м. В биоценозах батипелагиаля преобладают популяции низших ракообразных (копепод), медуз, амфипод. По функциональным характеристикам экосистемы | ||||||||||||||||||||||||||||||||||||