биология. Курсовая работа Основные гидробиомы Исполнитель студентка группы гэ22 Д. В. Котюжинская
 Скачать 2.8 Mb. Скачать 2.8 Mb.
|

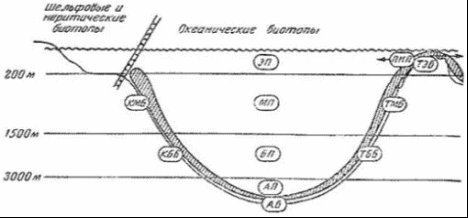 Рисунок 4 - Океанические биотопы [7] В расположенной ниже, до глубин порядка 2500 м, батипелагиали фауна рыб и кальмаров беднеет не только в количественном, но и в качественном отношении. Из океанических пелагических рыб для этой зоны наиболее характерны такие глубоководные рыбы, как мешкоротые и большеротые. Это сравнительно крупные рыбы, длиной от 60 до 180 см, и состоящие буквально из огромной, широко раскрывающейся пасти с крохотным черепом в виде придатка верхней челюсти и длинного угревидного тела. Кости нижней челюсти поддерживают гигантскую, растягивающуюся глотку. Эти рыбы приспособились схватывать и заглатывать жертву, превышающую по размерам самого хищника. В условиях бедности жизни на больших глубинах преимущество такого образа жизни становится несомненным. Близки к мешкоротым и небольшие рыбки - моногнаты, также очень характерные для батипелагиали, у которых верхняя челюсть редуцировалась, а огромная нижняя прикрепляется непосредственно к черепу. Несмотря на хищный образ жизни, как те, так и другие рыбы относятся скорее к жизненной форме планктона. Настоящими нектонными рыбами можно считать живущих на батиальных глубинах пелагических долгохвостов. Большинство же долгохвостов представлено в этой зоне придонно-нектическими видами. Вообще долгохвосты, или макрорусы, - наиболее многочисленная группа придонных глубоководных рыб - высокого разнообразия достигают в Тихом океане. Долгохвосты, близкие к тресковым рыбам, отличаются удлиненным, сходящим у хвоста на нет телом. Самый конец хвоста иногда заканчивается длинной тонкой нитью. Глаза у них очень крупные, способные, видимо, различать даже на значительном удалении слабый свет светящихся глубоководных рыб и планктонных ракообразных. Это, а также сложная сейсмосенсорная система позволяют долгохвостам обнаруживать добычу и успешно охотиться за ней. Возможно, что и обоняние у этих рыб развито достаточно хорошо. Применение специальных ловушек с приманкой и автоматически работающими фотокамерами показывает, насколько быстро у таких ловушек собираются десятки крупных макрурид. Кроме долгохвостов у дна держатся многие глубоководные угри, галозавры и спинношипы. Нектонная фауна кальмаров в батипелагиали складывается почти из тех же родов, что и в мезопелагиали. Большая группа батипелагических кальмаров включает виды, для которых характерен нектобентосный (придонный) образ жизни. Все эти кальмары представляют собой стайных хищников, они обладают средними (20 - 30 см) и даже крупными (до 1 м) размерами [8].  Рисунок 5 - Растительность батипелагиали [8] На рисунке 5 изображена растительность, которая распространена в батипелагиали. 1.3 Абиссопелагиаль Под термином «абиссопелагиаль» понимают водную толщу океана в пределах абиссали, которая простирается от нижней границы батиали (3 - 4 км) до глубины 6 - 7 км, где преобладают планктонные организмы крупнее 100 мм - макропланктон. На, уже упомянутом ранее, рисунке 3 хорошо видны границы абиссопелагиаля, который обозначается сочетанием букв АП. Среди рыб настоящие нектонные формы представлены лишь пелагическими макруридами. Ихтиофауна абиссопелагиали открытого океана очень бедна и слагается в основном из глубоководных удильщиков. В отличие от мелководных удильщиков, обычно ожидающих добычу лежа на дне, глубоководные виды постоянно обитают в толще воды. Они обладают чаще всего уродливым мешкообразным телом с огромной головой и широкой пастью, над которой свешивается «удочка» - видоизмененный первый луч спинного плавника - со светящимся органом на ее конце в виде приманки. Привлеченная светом жертва, часто не менее крупная, чем удильщик, захватывается загнутыми назад острыми длинными зубами. У глубоководных удильщиков наблюдается половой диморфизм, доходящий в своей крайности до фазы паразитизма самцов. Самка во много раз (в 10 - 13 раз в длину и в 5000 раз по весу) крупнее самца. У самцов удочка не образуется, а зубы превращаются в своеобразные щипчики для схватывания мелкой добычи, а в 4 семействах - для прикрепления к самке. Вскоре после внедрения зубов-щипчиков в кожу губы и язык самцов срастаются с телом самки, глаза и кишечник атрофируются, а питание осуществляется за счет крови самки. Личинки глубоководных удильщиков развиваются в планктоне поверхностных вод в тропической и субтропической зонах (от 40° с. ш. до 35° ю. ш.), однако взрослые рыбы, обитающие на глубине порядка 3000 м и глубже и при сравнительно одинаковой температуре воды в любой части океана (2 - 4 °С), встречаются в Тихом, да и в других океанах повсеместно. Постоянно держатся в абиссопелагиали и некоторые виды одночелюстных рыб. У подножия материкового склона в приконтинентальной полосе встречаются рыбы других семейств. В отличие от чисто океанических, окрашенных обычно в черный цвет и имеющих чешую, прибрежные нектобентосные, абиссальные рыбы характеризуются светлой кожей, лишенной чешуи, а иногда и глаз. Таковы, в частности, ипноповые рыбы, обладающие прогонистым телом и хорошо развитыми плавниками. На вытянутой уплощенной голове ипнопид под тонкими костными пластинками лежат так называемые головные органы, способные воспринимать свет. Настоящие глаза у них отсутствуют. У рыб близкого семейства - плавники тоже хорошо развиты, а отдельные лучи брюшных и хвостовых плавников достигают значительной длины. На этих лучах, как на треножнике, рыбы одного из родов батиптерид - могут подолгу стоять на дне. Всего в ихтиофауне прибрежной абиссопелагиали известно семейств рыб океанического происхождения. К ним можно приубавить еще три семейства глубоководных рыб шельфового происхождения [8]. 1.4 Ультраабиссопелагиаль Ультраабиссопелагиаль - толща воды над ультраабиссалью, простирающаяся от 6 - 7 км до максимальных глубин океана [9]. . Пелагиаль озёр Наука об озерах и прудах (лимнология) позволила нам лучше понять частичные кругообороты и их интеграцию. Озера и более мелкие пресные водоемы представляют собой хорошо отграниченные, легко поддающиеся описанию водные экосистемы. В них имеются как аэробные, так и анаэробные зоны. Такие зоны можно обнаружить и в большинстве почв; но если в почве они сосредоточены поблизости друг от друга в очень тесном пространстве и потому их трудно изучать, в озерах такие зоны весьма обширны и легко поддаются исследованию. Однако есть основания полагать, что результаты лимнологических исследований в принципе можно перенести и на почву с ее микрогетерогенностью. На биологические процессы в озерах и прудах большое влияние оказывают физические свойства воды. Вода имеет наибольшую плотность при 4 °С. С увеличением глубины меняется температура воды; может наблюдаться более или менее устойчивая слоистость (стратификация) в зависимости от времени года (рисунок 6). 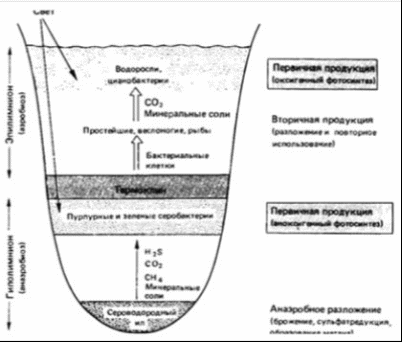 Рисунок 6 - Схематический вертикальный разрез эвтрофного озера [13] Стратификация характерна для озер двух типов. К одному типу относятся пресно водные озера умеренных широт. Весной холодная вода озера прогревается солнцем; поверхностный слой воды становится теплым, его плотность уменьшается. Этот слой называют эпилимнионом. Он лежит поверх более холодного слоя воды - гиполимниона. Слои разделены переходной зоной, называемой термоклином или металимнионом; эта граница между слоями иногда бывает очень резкой. В глубоких озерах такое разделение может сохраняться в течение всего лета. В результате процессов аэробного распада растворенный в воде кислород, начиная с придонного слоя, расходуется, и в гиполимнионе создаются анаэробные условия. Эпилимнион соприкасается с кислородом воздуха, перемешивается с помощью ветра, и поэтому, как правило, условия здесь остаются аэробными. Это приводит к образованию градиентов окислительно-восстановительного потенциала и химических параметров в области термоклина, в связи с чем термоклин называют также хемоклином. Осенью происходит охлаждение эпилимниона. Если температура эпилимниона становится ниже температуры гиполимниона, происходит перемешивание обоих слоев, чему способствуют осенние бури. В случае полного перемешивания глубинные слои воды поднимаются вверх и вновь обогащаются кислородом. Благодаря этому ежегодно восстанавливается равномерное распределение питательных веществ, которыми богаты глубинные воды. Озера с полным перемешиванием воды называют голомиктическими. Зимой может происходить обратное расслоение. Температура воды на глубине составляет 4 °С, а над ней находится слой более холодной воды, имеющей меньшую плотность, а иногда и слой льда. Когда весной температура в поверхностном слое поднимается выше 4 °С, стратификация снова нарушается. В озерах, как и в морях, различают планктон, нектон и бентос. Однако в озерах и других пресных водоемах зообентоса меньше, чем в морях и океанах, а видовой его состав однообразен. Главным образом это простейшие, губки, ресничные и малощетинковые черви, пиявки, моллюски, личинки насекомых и др. Фитобентос пресных вод представлен бактериями, диатомовыми и зелеными водорослями. Прибрежные растения располагаются от берега вглубь четко выраженными поясами. Первый пояс - полупогруженные растения (камыши, рогоз, осоки и тростники); второй пояс - погруженные растения с плавающими листьями (водокрас, кубышки, кувшинки, ряски). В третьем поясе преобладают растения - рдесты, элодея и др. По образу жизни водные растения подразделяют на две основные экологические группы: гидрофиты - растения, погруженные в воду только нижней частью и обычно укореняющиеся в грунте, и гидатофиты - растения, которые полностью погружены в воду, а иногда и плавающие на поверхности или имеющие плавающие листья. В жизни водных организмов большую роль играют вертикальное перемещение воды, плотность, температурный, световой, солевой, газовый (содержание кислорода и углекислого газа) режимы, концентрация водородных ионов (рН). Температурный режим отличается в воде, во-первых, меньшим притоком тепла, во-вторых, большей стабильностью, чем на суше. Часть тепловой энергии, поступающей на поверхность воды, отражается, часть расходуется на испарение. На рисунке 7 изображена растительность, произрастающая на пелагиале озер. Виды растений, укореняющихся на дне, такие как рогоз (1), ситник (2), стрелолист (3), кувшинка (4), рдесты (5,6), хара (7) и свободно плавающие водоросли, таки как нитчатые зеленые (8,9), зеленые (10 - 13), диатомеи (14 - 17), сине - зеленые (18 - 20). 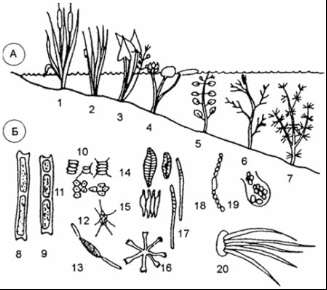 Растения, укореняющиеся на дне (А): 1 - рогоз; 2 - ситник; 3 - стрелолист; 4 - кувшинка; 5, 6 - рдесты; 7 - хара. Свободно плавающие водоросли (Б): 8, 9 - нитчатые зеленые; 10 - 13 - зеленые; 14 - 17 - диатомеи; 18 - 20 - синезеленые Рисунок 7 - Растительность, произрастающая на пелагиали озёр [15] Испарение воды с поверхности водоемов, при котором затрачивается около 2263×8 Дж/г, препятствует перегреванию нижних слоев, а образование льда, при котором выделяется теплота плавления (333,48 Дж/г), замедляет их охлаждение. Изменение температуры в текущих водах следует за ее изменениями в окружающем воздухе, отличаясь меньшей амплитудой. В озерах и прудах умеренных широт термический режим определяется хорошо известным физическим явлением - вода обладает максимальной плотностью при 4 °С. Вода в них четко делится на три слоя: верхний - эпилимнион, температура которого испытывает резкие сезонные колебания; переходный, слой температурного скачка, металимнион, где отмечается резкий перепад температур; глубоководный (придонный) - гиполимнион, доходящий до самого дна, где температура в течение года изменяется незначительно [15]. В замерзающих озерах средних широт в разные сезоны года в распределении температуры по глубине наблюдаются характерные особенности, связанные с сезонными колебаниями теплообмена в озере и перемешиванием. Годовой термический цикл в озерах умеренного пояса, по предложению Б.Д. Зайкова, разделяется на четыре периода: весеннего и летнего нагревания и осеннего и зимнего охлаждения. Продолжительность периодов и моменты их наступления зависят не только от времени года и географического положения озера. В значительной степени они так же обусловлены объемом массы воды в озере и морфологическими особенностями его котловины. Период весеннего нагревания начинается с момента устойчивого преобладания притока тепла в озеро в течении суток над его потерями. Это происходит обычно перед вскрытием озер при усилении солнечной радиации, проникающей через лед. В эту раннюю фазу весеннего нагревания наблюдается, как и зимой обратная стратификация. При подледном нагревании верхних слоев воды на десятые доли градуса выше нуля начинается частичная циркуляция, постепенно переходящая в полную после вскрытия озера и таяния льда. Наступает состояние гомотермии. Весь слой воды принимает температуру равную температуре придонных слоев. В дальнейшем прогревание происходит при гомотермии. Заканчивается период весеннего нагревания к моменту достижения температуры наибольшей плотности во всем озере. В этот период большая роль в передачи тепла с поверхности в глубину принадлежит динамическому перемешиванию. Длительность периода весеннего нагревания в небольших озерах невелика несколько дней после вскрытия. В глубоких озерах, например Телецком, 270 - метровая толща прогревается до июля; в среднем к 15 июля заканчивается период весеннего нагревания в Ладожском озере. В период летнего нагревания начинается с момента возникновения прямой стратификации во всем озере. По мере нагревания озера в условиях прямой стратификации разность температур и плотностей воды между поверхностными и глубинными слоями, особенно в безветренную погоду, резко возрастает. Конвекция, возникающая при ночном охлаждении, выравнивает температуру лишь в сравнительно тонком поверхностном слое. В результате в верхнем прогретом слое воды устанавливается более или менее одинаковые температуры. В нижних глубинных слоях сохраняются холодные «весенние» воды с плавным изменение температуры. Между теплым и холодным слоями возникает промежуточный, сравнительно тонкий слой с резким падение температуры с глубиной, иногда до 8 - 10 °С на 1 метр. Слой этот известен как слой температурного скачка, или металимниона. Слой воды расположенный выше металимниона называется эпилимнион, а ниже - гиполимнион. Подобное термическое расслоение на 3 хорошо выраженные термические зоны в период летнего нагревания характерно для многих озер. Слой температурного скачка в различных озерах располагается на разных глубинах. В озерах, незащищенных от действия ветра, слой температурного скачка расположен глубже, в неглубоких, хорошо прогреваемых озерах он либо вовсе отсутствует, либо слабо выражен. Слой температурного скачка исчезает при сильных ветрах, вызывающих смешение вод верхнего слоя с водами гиполимниона, а в штилевую погоду может появится вновь. Иногда при неустойчивой погоде может возникнуть второй и третий слой температурного скачка. К концу лета слой скачка погружается и осенью исчезает, уступая место плавному падению температуры с глубиной. Период осеннего охлаждения начинается со времени устойчивого преобладания в течении суток теплоотдачи водной поверхностью озера над поступлением тепла к ней. Обычно в озерах умеренных широт это время приходится на август. Первоначально охлаждение происходит в условиях прямой стратификации. Разность температур и плотностей между зонами эпилимниона и гиполимниона постепенно уменьшается. Этому способствует не только охлаждение поверхностных слоев, но и усиливающееся конвективное и поверхностное перемешивание, выравнивающее температуру по вертикали. Устанавливается осенняя гомотермия. Она может возникнуть при разных температурах в зависимости от температуры воды в зоне гиполимниона, морфометрических особенностей котловины озера и силы ветра. В мелководных открытых действию ветра водоемах гомотермия устанавливается раньше и при более высоких температурах, чем в глубоких. Дальнейшее охлаждение происходит при гомотермии. Окончание периода осеннего охлаждения считают время установления температуры наибольшей плотности во всей толще озера. Происходи это в разные сроки в зависимости от глубины озера, количества накопленного за лета тепла, например в озерах Ладожском и Севан к концу декабря. Период зимнего охлаждения начинается с момента установления обратной термической стратификации. В начале этого периода, до замерзания, в больших по площади, но мелководных озерах охлаждение всей водной массы происходит очень интенсивно, чему способствует ветровое перемешивание. Запасы тепла в таких озерах быстро истощаются и водоемы замерзают при очень низкой температуре всей воды в озере. Малые озера, защищенные от ветра, замерзают при больших запасах тепла в придонных слоях и температура у дна близка к 4 0С. При ледоставе, как указывалось, и потери тепла из водоема осуществляются через снего-ледяной покров. Если эти потери будут меньше, чем тепловой поток, направленный от дна в водную толщу, охлаждения водоема не происходит. В подобных случаях наблюдается зимние нагревание воды в озере. В незамерзающих полностью или поздно замерзающих озерах охлаждение воды происходит до конца зимнего периода. В период наибольшего охлаждения, в марте, температура придонных слоев в озерах Севан и Телецком, например, понижается до 2,5 °С. Зимой при обратной стратификации возможно образование слоя температурного скачка, выражен он менее отчетливо, чем летом. Располагается вблизи поверхности [16]. 2.1 Эпилимнион Эпилимнионом называют верхние слои воды в озере. Они теплее, чем более глубокие, в них проникает свет, и поэтому там может происходить фотосинтез и проживают растения. Перемешивание этих слоев происходит под воздействием ветра или внутренних течений. Эпилимнион образуется в озерах и водохранилищах в фазу летнего нагревания вследствие накопления тепла поглощаемой водой солнечной радиации и периодического (ночного и при похолоданиях) конвективного и динамического перемешивания верхнего слоя водоема. Толщина его в течение лета и первой фазы осенного охлаждения водоема возрастает по мере погружения подстилающего его сезонного слоя скачка от 1 - 2 м до 30 - 40 м в крупнейших озерах России (Ладожское, Онежское, Телецкое) и даже до 150 м в Байкале. Вода Э. в сравнении с другими слоями водоема наиболее прогрета и насыщена кислородом, содержит наибольшую биомассу фито-, зоо- и бактериопланктона и характеризуется наиболее интенсивной трансформацией энергии, биогенных и органических веществ. В антициклональную солнечную и штилевую погоду Э. может быть разделен на три подслоя. В верхнем из них особенно велик диапазон внутрисуточных колебаний всех характеристик физических, химических и биологических свойств воды. Под ним располагается вторичный (временный) слой скачка, который обычно размывается ночной конвекцией, выравнивающей к утру свойства воды по всей глубине. Наиболее интенсивен процесс вертикального перемешивания эпилимниона в периоды существования в нем упорядоченной конвекции [11]. Эпилимнион почти всегда насыщен растворенным кислородом, образующимся здесь в процессе фотосинтеза, а также захваченным из пограничного слоя атмосферы при циркуляции воды. В то же время все прочие элементы, необходимые для фотосинтеза и роста, извлекаются из воды водорослями, и химизм вод эпилимниона подвергается соответствующим изменениям. Одновременно эпилимнион производит много органического детрита, состоящего из отмерших фрагментов водорослей, опускающегося в гиполимнион. Там растворенный кислород затрачивается на дыхание и разложение, и многие неорганические вещества возвращаются в воду. Таким образом, в стратифицированном озере первоначально однородная водная масса подразделяется на два четко различающихся слоя: верхний, более теплый, с дефицитом доступных питательных элементов, и нижний, более холодный, с более высокой концентрацией питательных элементов. На рисунке 8 изображены температурные слои в озере [18]. 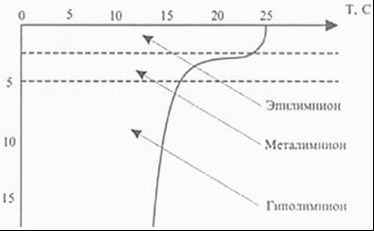 Рисунок 8 - Температурные слои в озере [13] В условиях умеренного климата это разделение имеет место и зимой и летом, хотя зимой оно менее выражено, поскольку подо льдом из-за меньшего доступа света значительно снижается уровень первичной продукции вод. В нестратифицированных озерах сезонные изменения происходят во всей водной толще. Во многих озерах, богатых питательными элементами, фотосинтез протекает настолько интенсивно, что растворенный кислород оказывается полностью израсходованным непосредственно у поверхности донных отложений. На поверхности раздела донных осадков и воды содержащие кислород нерастворимые соединения железа теряют кислород и становятся растворимыми, в результате чего большое количество железа, марганца, фосфора и азота поступает в воду. Этот процесс называется внутренней эвтрофикацией, так как в некоторых озерах в результате ветрового перемешивания или влияния внутренних сейш высвободившиеся из осадков питательные элементы попадают в верхний слой воды, повышая таким образом трофический уровень озера [13]. 2.2 Металимнион Промежуточный слой воды в озерах и водохранилищах, расположенный под эпилимнионом и над гиполимнионом. По глубине совпадает со слоем скачка температуры и плотности воды. В нем, кроме того, обычно наблюдаются большие вертикальные градиенты содержания кислорода, рН, биогенных и органических веществ. Толщина составляет несколько метров летом и увеличивается осенью до несколько десятков метров в самых глубоких озерах России. В эвтрофных водоемах при значительном вертикальном градиенте плотности воды в пикноклине над ним (в верхнем подслое металимниона) задерживается осаждающийся из эпилимниона обильный детрит. В результате интенсивного его разложения бактериопланктоном в этом подслое может наблюдаться наряду с повышенной мутностью и окисляемостью воды, пониженное ее насыщение кислородом [14]. 2.3 Гиполимнион Характерный для глубоких озер холодный, бедный кислородом слой воды, лежащий ниже зоны быстрого перепада температуры - термопланы. Если ниже глубины эффективного проникновения света (компенсаторного горизонта), запасы кислорода в гиполимнионе истощаются. В гиполимнионе нет зеленых растений, т. к. в него не проникает кислород атмосферы. В то же время, благодаря положительным температурам, зимой служит своеобразным убежищем для организмов, обитающих в верхних слоях водоемов [15]. Заключение Биом и, в частности, гидробиом представляет собой категорию хорологическую. Совокупности сходных по своему строению экосистем занимают вполне определенное пространство. Биом выглядит как “ареал” сходных экосистем. Определенное сходство состава жизненных форм индицирует сходство комплекса условий существования организмов. Существует определенная структура биомов как хорологических единиц биосферы. Биом также - категория экологическая и функциональная. Если идти от частного к общему, биом представляет собой систему сходных экосистем, надэкосистемное образование. При этом биомы неравноценны по роли биотического фактора. В этом аспекте может быть предложен, например, такой ряд по увеличению роли биоты: пустынный биом - пелагический биом океана - степной биом - прибрежный - гилея - биогермовый биом. С другой стороны (от общего - к частному), биосфера состоит из крупных функциональных блоков - биомов. Кроме того, гидробиом - категория гидробиологическая. Если рассматривать гидробиологию как науку о закономерностях существования определенным образом структурированного живого вещества в гидросфере, то гидробиом выступает как наибольшая структурно-функциональная единица деления гидросферы с её населением. В океане вместе с входящими в него морями различают прежде всего две экологические области: пелагиаль - толща воды и бенталь - дно. Население пелагиали (водной толщи) называется пелагос. Совокупность организмов, парящих в толще воды и неспособных к передвижению против течения, называется планктон. Различают фитопланктон (совокупность фотосинтезирующих планктонных организмов) и зоопланктон (совокупность планктонных организмов, неспособных к фотосинтезу). Организмы, способные к активному перемещению против течения, называются нектон. По вертикали пелагиаль делится на зоны (перечислены только основные): нейсталь - поверхностный слой воды, граничащий с атмосферой; население нейстали называется нейстон; организмы, часть тела которых находится в воде, а часть над ее поверхностью, называются плейстон; эпипелагиаль - соответствует глубине сублиторали; батипелагиаль соответствует глубине батиали; абиссопелагиаль соответствует глубине абиссали. Совокупность организмов, способных вести и пелагический, и бентосный образ жизни, называется пелагобентос. Совокупность организмов, обитающих на различных предметах и живых телах, находящихся в толще воды, называется перифитон. К благоприятным факторам эпипелагиали открытого океана относятся: достаточно высокая аэрация; высокая освещенность. Лимитирующим фактором является низкое содержание биогенов за счет их миграции в придонные воды. Однако концентрация биогенов может возрастать за счет апвеллинга - выноса глубинных вод на поверхность, например, в приполярных зонах. Основными продуцентами эпипелагиали являются планктонные диатомовые водоросли и перидинеи (способные к миксотрофному питанию) - около 1000 видов. Разнообразие планктона в открытом океане выше, чем на шельфе, поскольку многие виды стеногалинны и не переносят опреснения прибрежных вод. Благоприятным фактором абиссали и абиссопелагиали является стабильность условий обитания. К лимитирующим факторам относятся: отсутствие света и невозможность фотосинтеза; высокое давление. При снижении освещенности органы зрения у животных гипертрофируются, но при полном отсутствии света происходит полная редукция органов зрения. Для обитателей глубин характерна люминесценция с участием симбиотических светящихся бактерий. Из-за нехватки света отсутствуют фотосинтезирующие продуценты. Следовательно, глубоководные экосистемы являются зависимыми от экосистем эпипелагиали, и их собственная продуктивность стремится к нулю. При наличии неорганических окислителей (например, вблизи гидротермальных сульфатных источников) продуцентами являются десульфирующие и другие хемосинтезирующие бактерии. Они участвуют в образовании симбиотических систем с различными беспозвоночными. Список использованных источников 1 Голубев, Г.Н. Геоэкология: Учебное пособие для студентов высших учебных заведений / Г.Н. Голубев. - М.: Издательство ГЕОС, 1999. - 419 с. Давыдов, Л.К. Общая гидрология: Учебное пособие / Л.К. Давыдов, А.А. Дмитриева, Н.Г. Конкина. - М.: издание второе 1973. - 370 с. Лебедева, М.И. Экология: Учебное пособие / М.И. Лебедева - Тамбов: Издательство Тамб. гос. техн. ун-та, 2002. - 328 с; 4 Свободная энциклопедия «Википедия» [Электронный ресурс] / Режим доступа: |