Курсовая работа по дисциплине Ионные механизмы потенциалов покоя и действия
 Скачать 62.88 Kb. Скачать 62.88 Kb.
|

|
МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ РОССИЙСКОЙ ФЕДЕРАЦИИ ПЕНЗЕНСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ Кафедра «Общая биология и биохимия» Курсовая работа по дисциплине «Ионные механизмы потенциалов покоя и действия» Выполнил студент: Аслаева С.М. Группа:20ФБм2 Руководитель: доц. Гамзин С.С. Работа защищена с оценкой ____________ Дата защиты _________________ Пенза, 2019 Содержание Введение………………………………………………………………...………..3 Глава 1. Ионные механизмы потенциала действия…………………...……….5 Натриевые и калиевые токи…………………………………………………5 Рост потенциала……………………………………………………………...7 Глава 2. Идеальная клетка………………………………………………………9 2.1 Ионное равновесие………………………………………………………….10 Глава 3. Состояние покоя………………………………………………………14 Заключение……………………………………………………………………...19 Список литературы……………………………………………………………..20 Введение В состоянии покоя на мембране нейронов поддерживается постоянный электрический потенциал, при этом внутриклеточная среда заряжена отрицательно по отношению к внеклеточной. Поскольку внутри клетки больше ионов калия и меньше ионов натрия и хлора, чем снаружи, то каждый из этих ионов стремится проникнуть сквозь мембрану в направлении своего электрохимического градиента, то есть ионы калия стремятся наружу, а ионы натрия и хлора - внутрь клетки. Этому перемещению препятствует мембранный потенциал. Если представить себе упрощенную модель клетки, мембрана которой проницаема лишь для калия и хлора, то в ней возможен такой баланс между концентрационными градиентами и мембранным потенциалом, при котором суммарный ток каждого из этих ионов равен нулю. Тогда мембранный потенциал будет в точности равен равновесному потенциалу для этих двух ионов. Если в такой модели изменить внеклеточную концентрацию калия, то изменится равновесный потенциал для калия, а следовательно и мембранный потенциал. Изменение внеклеточной концентрации хлора, напротив, приведет к соответствующему изменению его внутриклеточного уровня. При этом ни равновесный потенциал для хлора, ни мембранный потенциал не изменятся. Мембрана реальной клетки проницаема также для натрия. В состоянии покоя ионы натрия непрерывно перемещаются внутрь клетки, снижая таким образом отрицательный заряд на мембране. Поскольку при этом калий уже не находится в равновесии, он начинает вытекать из клетки. Если бы не механизмы компенсации, то такое перемещение натрия и калия привело бы к изменению их внутриклеточных концентраций. Этого не происходит благодаря работе натрий-калиевого насоса-обменника, переносящего ионы натрия наружу и ионы калия внутрь клетки в пропорции 3:2. Мембранный потенциал покоя зависит от калиевого и натриевого равновесного потенциала, относительной проницаемости клеточной мембраны для этих ионов, а также от стехиометрии натрий-калиевого насоса. В состоянии покоя скорость переноса натрия и калия насосом в точности компенсирует пассивную утечку этих ионов. Натрий-калиевый обменник вносит также непосредственный вклад в мембранный потенциал (в размере нескольких милливольт), поскольку он перемещает больше положительных зарядов наружу, чем внутрь клетки. В зависимости от механизмов транспорта, равновесный потенциал для хлора может быть как более, так и менее отрицательным по отношению к потенциалу покоя. Наличие значительной хлорной проницаемости играет важную роль в обеспечении электрической стабильности многих клеток, несмотря на то, что распределение ионов хлора не столь важно для формирования потенциала покоя. Возникновение электрических сигналов в нервной клетке происходит в основном за счет изменения проницаемости мембраны для таких ионов, как натрий и калий. Увеличение проницаемости мембраны позволяет ионам перемещаться внутрь или наружу клетки в направлении своих электрохимических градиентов. Изменения проницаемости зависят от активации ионных каналов. Перемещение ионов через каналы в мембране приводит к изменению заряда на ней и, следовательно, мембранного потенциала. Чтобы понять принцип возникновения электрического сигнала, необходимо уяснить сущность ионных градиентов на мембране, а также то, как они влияют на потенциал покоя. 1. Ионные механизмы потенциала действия Количественное описание механизмов, участвующих в генерации потенциала действия, стало возможным благодаря методу измерения мембранных токов в условии фиксации потенциала. Этот метод позволяет определить, какой вклад вносят ионы того или иного типа в мембранный ток, а также вычислить величину и временной ход изменений соответствующих ионных проводимостей. Активация натриевой проводимости носит кратковременный характер, за ней следует инактивация. Увеличение калиевой проводимости продолжается до тех пор, пока не закончится деполяризация. Именно зависимость натриевой и калиевой проводимостей от мембранного потенциала и их попеременная активация качественно определяют как амплитуду, так и временной ход потенциала действия, равно как и другие мембранные характеристики, включая порог и рефрактерный период. Исследование проводимостей одиночных калиевых и натриевых каналов во время потенциала действия проводились в условиях фиксации потенциала участка мембраны. Наблюдаемые при этом принципы работы отдельных каналов соответствуют результатам, полученным ранее в экспериментах с фиксацией потенциала целой клетки: при деполяризации вероятность открытия натриевых и калиевых каналов возрастает. Возрастание вероятности происходит с тем же временным ходом, что и соответствующие токи в условиях фиксации потенциала. Так, натриевые каналы наиболее часто открываются в начале деполяризующего импульса и вероятность таких открытий падает по мере развития натриевой инактивации. В генерации потенциала действия могут принимать участие и другие катионные каналы. В некоторых клетках фаза роста потенциала действия определяется активацией кальциевых каналов, а реполяризация происходит благодаря активации различных типов калиевых каналов. 1.1 Натриевые и калиевые токи Потенциал покоя зависит главным образом от разности концентраций калия (как было предложено Бернштейном в 1902 году), и, в меньшей степени, от концентраций натрия и хлора. Одновременно с созданием Бернштейном теории потенциала покоя, Овертон сделал важное открытие: он показал, что для генерации нервом потенциала действия необходим натрий, и сделал робкое предположение о том, что основой потенциала действия является вход ионов натрия в клетку. Развитие эта гипотеза получила благодаря опытам на аксоне кальмара. В 1939 году Ходжкин и Хаксли показали, что на пике потенциала действия происходит кратковременная смена знака («овершут») потенциала на внутренней стороне мембраны в область положительных значений. Это наблюдение подтвердило участие натрия в потенциале действия, поскольку вход натрия в клетку происходит и при положительных значениях мембранного потенциала вплоть до натриевого равновесного потенциала ENa. Спустя десятилетие Ходжкин и Катц добились уменьшения явления овершута путем снижения внеклеточной концентрации натрия, а следовательно, и ЕNa. Они пришли к выводу, что потенциал действия есть результат значительного, хотя и кратковременного, увеличения натриевой проводимости мембраны. Сегодня известно, что это увеличение происходит за счет открытия огромного числа потенциалзависимых натриевых каналов. Чем объясняется фаза спада потенциала действия? Можно предположить, что спад происходит просто в результате закрытия натриевых каналов. Это, действительно, один из факторов. Однако, если бы это был единственный механизм, то возврат мембранного потенциала к уровню потенциала покоя в большинстве клеток происходил бы гораздо медленнее, чем в действительности. Причина этого в том, что общая проводимость мембраны в покое достаточно мала, и выход накопленных в результате потенциала действия положительных ионов из клетки занял бы несколько миллисекунд, а то и десятки миллисекунд. В действительности, возврат к уровню потенциала покоя происходит гораздо быстрее благодаря значительному увеличению другой мембранной проводимости, а именно, за счет открытия калиевых каналов. С той же быстротой, с которой мембранный потенциал деполяризовался в направлении ENa, он гиперполяризуется на фазе спада потенциала действия в направлении ЕK. Увеличение калиевой проводимости может продолжаться до нескольких миллисекунд, что приводит во многих клетках не только к реполяризации, но и к гиперполяризации мембраны до значений более отрицательных, чем потенциал покоя. 1.2 Рост потенциала Если учесть, что на фазе роста потенциала действия в клетку входит большое количество натрия и большое количество калия ее покидает на фазе спада, то становится очевидным, что концентрации этих ионов в цитоплазме должны измениться. Величину этих изменений можно определить либо экспериментально, либо путем расчетов. Вычисления соотношения между мембранным потенциалом и трансмембранных концентраций. При потенциале -67 мВ на внутренней поверхности мембраны находится приблизительно 4 • 1011 отрицательных зарядов на см2. На пике потенциала действия (+40 мВ) вместо этого отрицательного заряда внутри клетки накапливается около 2,4 • 1011 положительных зарядов, что происходит в результате входа в клетку 6,4 • 1011 ионов натрия на см2. Это соответствует приблизительно 10–12 моль/см2. Экспериментальные измерения входа радиоактивного натрия и выхода радиоактивного калия дали значения между 3 • 10–12 и 4 • 10–12 моль/см2. Различие между теоретически предсказанным и экспериментальным значениями объясняются главным образом тем, что при расчетах не принималось во внимание то, что вход натрия и выход калия частично перекрываются во времени. Таким образом, количество входящего натрия превышает значение, необходимое для деполяризации мембраны до пикового уровня потенциала действия, поскольку выход калия начинается до момента достижения пика. Каково влияние входа натрия на его внутриклеточную концентрацию? Сегмент аксона кальмара длиной 1 см и диаметром 1 мм имеет площадь поверхности около 0,31 см2, поэтому при скорости входа натрия 3,5 • 10–12 приведет к накоплению приблизительно 10–12 M натрия. Объем этого отрезка аксона составляет 7,85 • 10–12 л, в нем содержится (при плотности 50 ммоль/л) 4 • 10–7 M натрия, поэтому изменение концентрации натрия в результате потенциала действия очень мало и составляет 0,0000025. Выход калия производит сравнимый эффект и изменяет внутриклеточную концентрацию калия в 0,000003 раз. Потенциал действия в более тонких отростках нейрона может привести к более значительным изменениям внутриклеточных концентраций натрия и калия, чем в гигантском аксоне кальмара. Так, нервное окончание диаметром 1 мкм и длиной 100 мкм имеет площадь поверхности 3 • 10–6 см2 и объем 8• 10–14 л. Во время потенциала действия вход натрия 3 • 10–6 моль/см2 производит увеличение количества натрия внутри терминалы на 10–17 М. При внутриклеточной концентрации 20 мМ в терминалы содержится приблизительно 1,5•10–15 М, так что увеличение концентрации натрия в результате одного потенциала действия составляет 0,7 %. Пачка из 50 импульсов теоретически приведет к увеличению уровня натрия на 35 % и сходному снижению уровня калия. Вход натрия увеличивает активность натрий-калиевого обменника так, что концентрации быстро возвращаются к изначальному уровню. Главным свойством натриевой и калиевой проводимостей, определяющим характер токов во время потенциала действия, является их потенциалзависимость: вероятность открытия ионных каналов увеличивается с деполяризацией мембранного потенциала. Деполяризация увеличивает натриевую проводимость, а также, с некоторой задержкой, калиевую. Воздействие деполяризации на натриевую проводимость носит регенеративный характер: сначала небольшая деполяризация увеличивает количество открытых каналов; ионы натрия, входящие в клетку по направлению своего электрохимического градиента, производят дальнейшую деполяризацию мембраны, открытие большего количества каналов, что влечет за собой вход еще большего числа ионов, и так далее. Такой самоусиливающийся процесс характеризуется положительной обратной связью. Потенциал--зависимость калиевых каналов, наоборот, характеризуется наличием отрицательной обратной связи. При деполяризации количество открытых калиевых каналов возрастает, и ионы калия входят в клетку в направлении электрохимического градиента. Однако, вход калия не усиливает деполяризацию, а ведет к реполяризации и возвращению калиевой проводимости в состояние покоя. То, что до сих пор обсуждалось, было предложено Ходжкином, Хаксли и Катцем и детально разработано Ходжкином и Хаксли, которые поставили элегантные опыты на гигантском аксоне кальмара. Они экспериментально показали, что при потенциале действия происходят изменения калиевой и натриевой проводимостей, и что последовательность этих изменений в точности определяет временной ход и величину потенциала действия. На основании каких опытов был сделан этот вывод? На первый взгляд, измерение натриевой (gNa) и калиевой (gк) проводимостей кажется простой задачей. Нужно только измерить величину тока (I), протекающего через мембрану при том или ином потенциале (Vm), поскольку: Однако, прежде необходимо решить две проблемы. Во-первых, ток, протекающий через мембрану, приведет к изменению мембранного потенциала; это, в свою очередь, вызовет изменение проводимостей. Решение было найдено путем разработки метода, позволяющего быстро устанавливать заданный мембранный потенциал и поддерживать его на постоянном уровне, при этом измеряя величину и временной ход мембранного тока. Поскольку потенциал фиксирован в течение всего времени эксперимента, ток будет аккуратно отражать изменения проводимости мембраны. Вторая проблема состоит в том, чтобы разделить различные компоненты мембранного тока для того, чтобы измерить их индивидуальные характеристики. Для преодоления этого затруднения применялись различные методы, в том числе замена натрия на ион, не проникающий через мембрану, а позднее использовались селективные токсины и яды. 2. Идеальная клетка Начнем с рассмотрения модели идеальной клетки. В этой клетке содержатся ионы калия, натрия и хлора, а также крупные анионы. Клетка помещена в раствор, содержащий ионы натрия и калия. Другие ионы, присутствующие в реальных клетках, не играют важной роли в формировании потенциала покоя и потому не рассматриваются в данной модели. Концентрации ионов внутри и снаружи клетки приблизительно соответствуют средним величинам, полученным на клетках лягушки. В клетках птиц и млекопитающих эти значения несколько выше, а у таких морских беспозвоночных, как кальмар - значительно выше, чем у лягушки (см. табл. 1.1). В настоящей модели рассматривается клеточная мембрана, проницаемая для калия и хлора, но не для натрия и внутриклеточных анионов. Для того чтобы клетка пребывала в стабильном состоянии, необходимо выполнение трех условий: 1.1 Распределение ионов в идеальной клетке. Мембрана проницаема для К и Cl, но непроницаема для Na и внутриклеточных анионов (А). Концентрационный градиент ионов калия способствует выходу этих ионов из клетки (черная стрелка); градиент потенциала стремится перенести ионы калия внутрь клетки (серая стрелка). В состоянии покоя эти две силы уравновешивают друг друга. Концентрационный и электрический градиенты для хлора имеют противоположные направления. Ионные концентрации выражены в миллимолях (ммоль). 1. Как внутриклеточная, так и внеклеточная среды должны быть электрически нейтральными. Так, например, раствор не может состоять из одних лишь ионов хлора. Отрицательный заряд этих ионов должен компенсироваться такими положительными ионами, как натрий или калий (иначе взаимное отталкивание одинаково заряженных частиц приведет к разрушению раствора). 2. Клетка должна находиться в осмотическом равновесии. В противном случае вода будет проникать в клетку (или вытекать из нее) до тех пор, пока равновесие не будет достигнуто. Осмотическое равновесие наступает при условии равенства суммарной концентрации растворенных частиц внутри и вне клетки. 3. Суммарный заряд каждого отдельно взятого иона, переносимый через мембраны клетки, должен равняться нулю. 2.1 Ионное равновесие Как создаются и поддерживаются ионные градиенты и соответствующий электрический потенциал? На рис.1.1 показано, что ионы находятся в положении обратной пропорциональности: ионы калия более концентрированы внутри клетки, а ионы хлора снаружи. Представим себе, что мембрана клетки проницаема только для ионов калия. Возникает вопрос, почему эти ионы не диффундируют из клетки наружу до тех пор, пока концентрации внутри и снаружи клетки не сравняются. Причина этого заключается в том, что если ионы калия покидают клетку, то снаружи накапливается положительный заряд, а внутри клетки образуется избыток заряда отрицательного. Возникший таким образом электрический потенциал снижает скорость перемещения ионов калия, а при достижении определенного уровня приводит к полному его прекращению. Это равновесный потенциал для калия (Ек). При потенциале Ек электрический градиент полностью уравновешивает градиент химический, в результате чего движение ионов прекращается. Отдельные ионы калия по прежнему втекают в клетку и вытекают из нее, но суммарный ток равен нулю. Ионы калия находятся в равновесии. Условия нахождения ионов калия в равновесии те же самые, что описаны и при рассмотрении нулевого суммарного тока через одиночный канал в небольшом участке (patch) мембраны. В этом случае концентрационный градиент уравновешивался потенциалом, приложенным к patch-электроду. Важное отличие описываемой здесь ситуации в том, что перемещение ионов само по себе производит электрический потенциал, уравновешивающий и останавливающий это перемещение. Другими словами, равновесие в данной модели достигается автоматически и является неизбежным. Напомним, что в главе 2 равновесный потенциал для калия получался из уравнения Нернста: где [К] 1 и [К] 0 - внутриклеточная и внеклеточная концентрации калия, соответственно. Для клетки, изображенной на рис.1.1, получим ЕK = - 85 мВ. Допустим теперь, что в мембране, кроме калиевых, присутствуют еще и хлорные каналы. Поскольку для анионов z = - 1, получим равновесный потенциал для хлора: или, пользуясь свойствами логарифма, Для модели идеальной клетки получим соотношение концентраций хлора, которое тоже равно 1: 30, и хлорный равновесный потенциал, равный - 85 мВ. Как и в случае с калием, мембранный потенциал величиной в - 85 мВ в точности уравновешивает стремление ионов хлора двигаться в направлении их концентрационного градиента, т.е. внутрь клетки. Обобщая, можно заключить, что мембранный потенциал препятствует перемещению как хлора внутрь клетки, так и калия из клетки наружу. Равновесные потенциалы для двух ионов равны благодаря тому, что соотношения их внеклеточных и внутриклеточных концентраций одинаковы (1: 30). Поскольку калий и хлор единственные ионы в нашей модели, способные проникать через мембрану клетки, и при - 85 мВ оба иона находятся в равновесии, то клетка может находиться сколь угодно долго в состоянии покоя, при котором суммарное перемещение ионов в клетку и из клетки будет равно нулю. Перемещение ионов калия наружу и ионов хлора внутрь клетки приводит к накоплению отрицательного заряда в клетке и положительного - во внеклеточном пространстве. На первый взгляд, такая ситуация противоречит принципу электрической нейтральности, однако это не так. Ионы калия, покидая клетку, накапливаются в непосредственной близости от ее мембраны, в то время как их отрицательно заряженные спутники остаются внутри клетки вблизи от внутренней поверхности той же мембраны. И те и другие фактически оказываются вне основной части раствора, как вне-, так и внутриклеточного. Также и ионы хлора, входя в цитоплазму, остаются вблизи от мембраны, а их спутники, оставленные перед проходом через мембрану, скапливаются поблизости от нее. Образуются два слоя ионов - катионов снаружи и анионов внутри клетки, которые удерживаются у мембраны благодаря взаимному притяжению. Таким образом, мембрана играет роль электрической емкости, разделяющей и запасающей заряд. Вышесказанное не означает, что ионы калия и хлора прикованы к поверхностям мембраны. Отдельные ионы свободно обмениваются с ионами внутри - или внеклеточного раствора. Тем не менее, заряд, накопленный на мембране, остается неизменным, а растворы - нейтральными. Интересным представляется вопрос, какую долю от общего количества ионов в клетке составляют ионы, накапливающиеся на мембране. Доля их весьма незначительна. Если предположить, что диаметр клетки составляет 25 мкм, то при концентрации 120 ммоль общее количество катионов (а следовательно, и анионов) получится 4 1012. При мембранном потенциале - 85 мВ величина заряда, разделенного мембраной, составляет приблизительно 5 1011 одновалентных ионов на см2. При площади поверхности клетки 8 10-5 см2 получается, что на внутренней поверхности мембраны накапливается около 4 107 отрицательных ионов, или одна стотысячная часть общего числа ионов во внутриклеточном растворе. Следовательно, перемещение ионов калия и хлора, достаточное для создания мембранного потенциала, никак не влияет на концентрации ионов в клетке. На мембранный потенциал нейронов, а также многих других клеток, влияют изменения внеклеточной концентрации калия, но не хлора. Обратимся к модели идеальной клетки. Сделаем допущение (для рассмотрения данной ситуации), что объем внеклеточной жидкости бесконечно велик, и что перемещение ионов не влечет за собой значительного изменения концентрации ионов вне клетки. На рис.1.2А показаны изменения внутриклеточного ионного состава и мембранного потенциала, вызванные повышением внеклеточного уровня калия с 3 до 6 ммоль. Для сохранения первоначальной осмолярности, одновременно с добавлением 3 ммоль калия из раствора удалили 3 ммоль натрия, в результате чего общая концентрация растворенных ионов осталась на уровне 240 ммоль. Увеличение внеклеточной концентрации калия приводит к снижению его трансмембранного градиента, движущего ионы из клетки наружу. При этом на начальном этапе мембранный потенциал не меняется. В результате происходит перенос суммарного положительного заряда внутрь клетки. Вследствие накопления положительного заряда на внутренней поверхности мембраны она деполяризуется. Ионы натрия благодаря деполяризации выходят из состояния равновесия и начинают двигаться внутрь клетки. Перемещение ионов калия и хлора продолжается до тех пор, пока не будет достигнуто новое равновесное состояние, основанное на новом соотношении концентраций и новом уровне мембранного потенциала, в данном примере - 68 мВ. Вход калия и хлора сопровождается током в клетку некоторого количества воды, приводящим к небольшому увеличению объема клетки. В процессе достижения нового состояния равновесия концентрация калия возрастает с 90 до 91 ммоль, хлора - с 4 до 7,9 ммоль, а объем клетки увеличивается на 3,5%. На первый взгляд кажется, что количество вошедшего в клетку хлора превышает количество калия, но представим себе, каковы были бы эти концентрации, если бы объем клетки не увеличился: концентрации обоих ионов были бы выше на 3,5%. Следовательно, вместо 7,9 ммоль концентрация хлора была бы 8,2 ммоль, а концентрация калия была бы на уровне 94,2 ммоль, то есть увеличение обеих концентраций составило бы 4,2%. Таким образом, сначала калий и хлор входят в клетку в приблизительно равных количествах (за исключением незначительного количества, необходимого для изменения заряда на мембране), а уже затем вода втекает в клетку, снижая концентрации обоих ионов до окончательного уровня. Влияние изменений внеклеточных концентраций ионов на внутриклеточные концентрации и на мембранный потенциал. (А) Внеклеточный уровень калия удвоен, а концентрация натрия снижена с целью сохранения осмолярности на прежнем уровне. (В) Половина ионов внеклеточного хлора заменена на анионы, не проникающие через мембрану (А). Ионные концентрации выражены в миллимолях (ммоль), объем внеклеточного пространства рассматривается как бесконечно большой в сравнении с объемом клетки, поэтому вход и выход ионов через мембрану не изменяет их внеклеточных концентраций. Сходные рассуждения применимы и в случае увеличения внеклеточной концентрации хлора, с той лишь разницей, что при этом мембранный потенциал не меняется. В показан результат 50-процентного снижения концентрации хлора во внеклеточном растворе путем замены 60 ммоль хлора на другой анион, не проникающий через мембрану. Ионы хлора вытекают из клетки, тем самым деполяризуя мембрану до нового хлорного равновесного уровня (-68 мВ). Калий, выведенный из равновесия, также начинает вытекать из клетки. Как и в предыдущем примере, оба иона покидают клетку в равных количествах, увлекая за собой воду. Снижение внутриклеточного уровня калия невелико в процентном отношении, поскольку начальная его концентрация в клетке высока. Напротив, снижение внутриклеточного уровня хлора весьма существенно, и оно влечет за собой значительное изменение равновесного потенциала для хлора. По мере выхода хлора из клетки значение равновесного хлорного потенциала вновь приближается к первоначальному. Процесс продолжается до тех пор, пока равновесные потенциалы для хлора и калия не сравняются и мембранный потенциал не восстановит свое прежнее значение. 3. Состояние покоя О состоянии покоя в возбудимых тканях говорят в том случае, когда на ткань не действует раздражитель из внешней или внутренней среды. При этом наблюдается относительно постоянный уровень метаболизма, нет видимого функционального отправления ткани. Мембранный потенциал (или потенциал покоя) – это разность потенциалов между наружной и внутренней поверхностью мембраны в состоянии относительного физиологического покоя. Потенциал покоя возникает в результате двух причин: 1) неодинакового распределения ионов по обе стороны мембраны. Внутри клетки находится больше всего ионов К, снаружи его мало. Ионов Na и ионов Cl больше снаружи, чем внутри. Такое распределение ионов называется ионной асимметрией; 2) избирательной проницаемости мембраны для ионов. В состоянии покоя мембрана неодинаково проницаема для различных ионов. Клеточная мембрана проницаема для ионов K, малопроницаема для ионов Na и непроницаема для органических веществ. За счет этих двух факторов создаются условия для движения ионов. Это движение осуществляется без затрат энергии путем пассивного транспорта – диффузией в результате разности концентрации ионов. Ионы K выходят из клетки и увеличивают положительный заряд на наружной поверхности мембраны, ионы Cl пассивно переходят внутрь клетки, что приводит к увеличению положительного заряда на наружной поверхности клетки. Ионы Na накапливаются на наружной поверхности мембраны и увеличивают ее положительный заряд. Органические соединения остаются внутри клетки. В результате такого движения наружная поверхность мембраны заряжается положительно, а внутренняя – отрицательно. Внутренняя поверхность мембраны может не быть абсолютно отрицательно заряженной, но она всегда заряжена отрицательно по отношению к внешней. Такое состояние клеточной мембраны называется состоянием поляризации. Движение ионов продолжается до тех пор, пока не уравновесится разность потенциалов на мембране, т. е. не наступит электрохимическое равновесие. Момент равновесия зависит от двух сил: 1) силы диффузии; 2) силы электростатического взаимодействия. Значение электрохимического равновесия: 1) поддержание ионной асимметрии; 2) поддержание величины мембранного потенциала на постоянном уровне. В возникновении мембранного потенциала участвуют сила диффузии (разность концентрации ионов) и сила электростатического взаимодействия, поэтому мембранный потенциал называется концентрационно-электрохимическим. Для поддержания ионной асимметрии электрохимического равновесия недостаточно. В клетке имеется другой механизм – натрий-калиевый насос. Натрий-калиевый насос – механизм обеспечения активного транспорта ионов. В клеточной мембране имеется система переносчиков, каждый из которых связывает три иона Na, которые находятся внутри клетки, и выводит их наружу. С наружной стороны переносчик связывается с двумя ионами K, находящимися вне клетки, и переносит их в цитоплазму. Энергия берется при расщеплении АТФ. Работа натрий-калиевого насоса обеспечивает: 1) высокую концентрацию ионов К внутри клетки, т. е. постоянную величину потенциала покоя; 2) низкую концентрацию ионов Na внутри клетки, т. е. сохраняет нормальную осмолярность и объем клетки, создает базу для генерации потенциала действия; 3) стабильный концетрационный градиент ионов Na, способствуя транспорту аминокислот и сахаров. При действии порогового или сверхпорогового раздражителя изменяется проницаемость клеточной мембраны для ионов в различной степени. Для ионов Na она повышается в 400–500 раз, и градиент нарастает быстро, для ионов К – в 10–15 раз, и градиент развивается медленно. В результате движение ионов Na происходит внутрь клетки, ионы К двигаются из клетки, что приводит к перезарядке клеточной мембраны. Наружная поверхность мембраны несет отрицательный заряд, внутренняя – положительный. Возбуждение нервной клетки под действием химического сигнала (реже электрического импульса) приводит к возникновению потенциала действия. Это означает, что потенциал покоя -60 мВ скачком изменяется на +30 мВ и спустя 1 мс принимает исходное значение. Процесс начинается с открывания Nа+-канала. Ионы Na+устремляются в клетку (по градиенту концентрации), что вызывает локальное обращение знака мембранного потенциала . При этом Na+-каналы тотчас закрываются, т. е. поток ионов Na+ в клетку длится очень короткое время. В связи с изменением мембранного потенциала открываются (на несколько мс) потенциал-управляемые К+-каналы и ионы К+ устремляются в обратном направлении, из клетки. В результате мембранный потенциалпринимает первоначальное значение, и даже превышает на короткое время потенциал покоя. После этогонервная клетка вновь становится возбудимой. За один импульс через мембрану проходит небольшая часть ионов Na+ и К+, и концентрационные градиенты обоихионов сохраняются (в клетке выше уровень К+, а вне клетки выше уровень Na+). Поэтому по мере получения клеткойновых импульсов процесс локального обращения знака мембранного потенциала может повторяться многократно. Распространение потенциала действия по поверхности нервной клетки основано на том, что локальное обращениемембранного потенциала стимулирует открывание соседних потенциал-управляемых ионных каналов, в результате чего возбуждение распространяется в виде деполяризационной волны на всю клетку. Восходящая ветвь графика: 1. потенциал покоя – исходное обычное поляризованное электроотрицательное состояние мембраны (–70 мВ); 2. нарастающий локальный потенциал – пропорциональная раздражителю деполяризация; 3. критический уровень деполяризации (–50 мВ) – резкое ускорение деполяризации(за счет самораскрытия натриевых каналов), с этой точки начинается спайк – высокоамплитудная часть потенциала действия; 4. самоусиливающаяся круто нарастающая деполяризация; 5. переход нулевой отметки (0 мВ) – смена полярности мембраны; 6. «овершут» – положительная поляризация (инверсия, или реверсия, заряда мембраны); 7. пик (+30 мВ) – вершина процесса изменения полярности мембраны, вершина потенциала действия. Нисходящая ветвь графика: 8. реполяризация– восстановление прежней электроотрицательности мембраны; 9. переход нулевой отметки (0 мВ) – обратная смена полярности мембраны на прежнюю, отрицательную; 10. переход критического уровня деполяризации (–50 мВ) – прекращение фазы относительной рефрактерности (невозбудимости) и возврат возбудимости; 11. следовые процессы (следовая деполяризация или следовая гиперполяризация); 12. восстановление потенциала покоя – норма (–70 мВ). 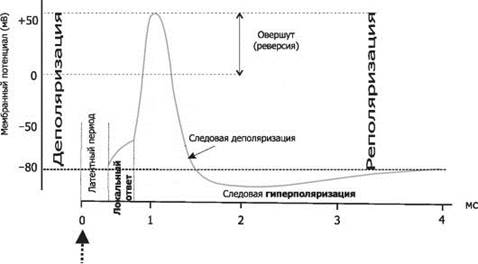 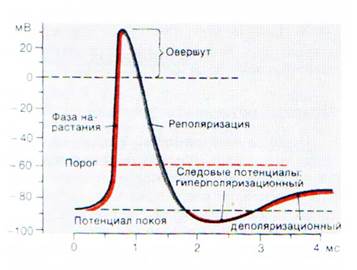 Заключение Потенциал действия является результатом быстрого и значительного увеличения натриевой проводимости мембраны. Вход большого количества ионов хлора и аккумуляция положительного заряда на внутренней поверхности мембраны клетки сдвигает мембранный потенциал в направлении ENa. Реполяризация мембраны происходит в результате последующего увеличения калиевой проводимости и уменьшения внутриклеточного положительного заряда из-за выхода ионов калия из клетки. Мембранный потенциал при этом вновь приближается к Ек. Такое объяснение механизма генерации потенциала действия ведет к пониманию принципов распространения импульса. Список литературы 1.Батуев А.С. Высшая нервная деятельность. – М.: Высшая школа, 1991. 2.Бреже М. Электрическая активность нервной системы. – М., 1979. 3.Борукаев Р.К. Принципы настройки в деятельности ЦНС. – М.: Наука, 1991. 4.Дмитриев А.С. Физиология высшей нервной деятельности. – М.: Высшая школа, 1974. 5. Иваницкий А.М. Мозговые механизмы оценки сигналов. – М., 1976. 6. Кацнельсон 3. С. Клеточная теория в ее историческом развитии. - Л., 1963. 7.Медведев С. Что знает наука о мозге // Наука и жизнь. – 1998. – № 8. – С. 58-64. 8. Методы клинической нейрофизиологии изучение физиологии головного мозга человека // Под ред. В. Б. Гречина. – Л., 1977. 9.Мозг // Под ред. В. П. Симонова. – М.: Мир, 1982. 10. Ройтбак А.И. Биоэлектрические явления в коре больших полушарий. – Тбилиси, 1955. 11.Ротенберг Г. Мозг. Стратегия полушарий // Наука и жизнь. – 1984. - № 6. – С. 54-57. 12.Симонов П.В. Лекции о работе головного мозга. – М.: институт психологии РАН, 1998. 13.Скулачев В.П. Энергетика биологических мембран. – М.: Наука, 1980. 14. Шагас Ч. Вызванные потенциалы мозга в норме и патологии. – М., 1975. |