Генетика. Лекции 45 Деление клетки и воспроизведение
 Скачать 356.44 Kb. Скачать 356.44 Kb.
|

|
Лекции 4-5 Деление клетки и воспроизведение Цель лекции: ознакомить учащихся с основными этапами клеточного цикла, изучить непрямое деление клетки, амитоз, эндомитоз, мейоз, их особенности и биологическое значение, познакомить учащихся с основными этапами гаметогенеза. План лекции: Клеточный цикл. Непрямое деление клетки. Амитоз. Эндомитоз Мейоз и его значение 4. Краткий обзор этапов гаметогенеза.1.Клеточный цикл Клетки многоклеточного организма чрезвычайно разнообразны по выполняемым функциям. В соответствии со специализацией клетки имеют разную продолжительность жизни. Так нервные клетки после завершения эмбриогенеза перестают делиться и функционируют на протяжении всей жизни организма. Клетки же других тканей (костного мозга, эпидермиса, эпителия тонкого кишечника) в процессе выполнения своей функции быстро погибают и замещаются новыми в результате клеточного деления. Деление клеток лежит в основе развития, роста и размножения организмов. Деление клеток также обеспечивает самообновление тканей на протяжении жизни организма и восстановление их целостности после повреждения. Существует два способа деления соматических клеток: амитоз и митоз. Преимущественно распространено непрямое деление клеток (митоз). Размножение с помощью митоза называют бесполым размножением, вегетативным размножением или клонированием. Жизненный цикл клетки(клеточный цикл) – это существование клетки от деления до следующего деления или смерти. Продолжительность клеточного цикла в размножающихся клетках составляет 10-50 ч и зависит от типа клеток, их возраста, гормонального баланса организма, температуры и других факторов. Детали клеточного цикла варьируют среди разных организмов. У одноклеточных организмов жизненный цикл совпадает с жизнью особи. В непрерывно размножающихся тканевых клетках клеточный цикл совпадает с митотическим циклом. Митотический цикл - совокупность последовательных и взаимосвязанных процессов в период подготовки клетки к делению и период деления (рис 1). В соответствие с приведенным выше определением митотический цикл подразделяют на интерфазу и митоз (греч. “митос” - нить). Интерфаза - период между двумя делениями клетки - подразделяется на фазы G1, S и G2 (ниже указана их продолжительность, типичная для растительных и животных клеток.). По продолжительности интерфаза составляет большую часть митотического цикла клетки. Наиболее вариабельны по времени G1 и G2-периоды. G1 (от англ. grow – расти, увеличиваться). Продолжительность фазы составляет 4–8 ч. Это фаза начинается сразу после образования клетки. В этой фазе в клетке усиленно синтезируются РНК и белки, повышается активность ферментов, участвующих в синтезе ДНК. Если клетка в дальнейшем не делится, то переходит в фазу G0 – период покоя. С учетом периода покоя клеточный цикл может длиться недели или даже месяцы (клетки печени). S (от англ. synthesis - синтез). Длительность фазы составляет 6–9 ч. Масса клетки продолжает увеличиваться, и происходит удвоение хромосомной ДНК. Две спирали старой молекулы ДНК расходятся, и каждая становится матрицей для синтеза новых цепей ДНК. В результате каждая из двух дочерних молекул обязательно включает одну старую спираль и одну новую. Тем не менее хромосомы остаются одинарными по структуре, хотя и удвоенными по массе, так как две копии каждой хромосомы (хроматиды) все еще соединены друг с другом по всей длине. После завершения фазы S митотического цикла клетка не сразу начинает делиться. G2.В этой фазе в клетке завершается процесс подготовки к митозу: накапливается АТФ, синтезируются белки ахроматинового веретена, удваиваются центриоли. Масса клетки продолжает увеличиваться до тех пор, пока она приблизительно вдвое не превысит начальную, а затем наступает митоз. 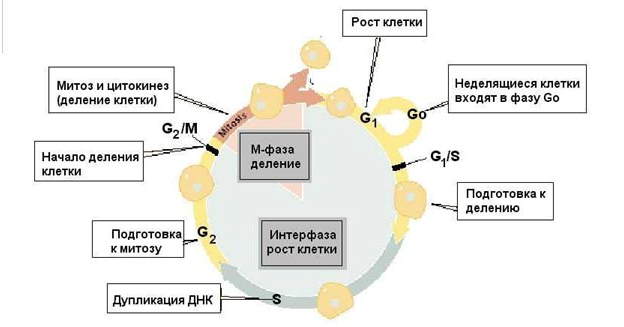 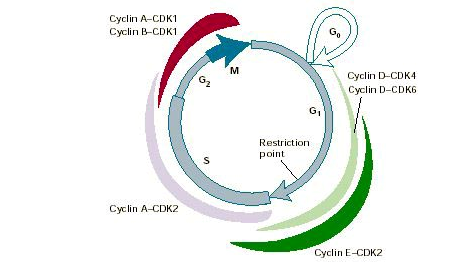 Жизнь клетки и переход от одной фазы клеточного цикла к другой регулируется изменением концентраций белков циклинов, как это показано на рисунке. Толщина цветных секторов соответствует концентрации циклинов. При подготовке к делению происходит репликация ДНК, на каждой хромосоме синтезируется ее копия. Пока эти хромосомы после удвоения не расходятся, каждая хромосома в этой паре называется хроматидой. После репликации ДНК конденсируется, хромосомы приобретают более компактную укладку, и в таком состоянии их можно увидеть в световом микроскопе. Между делениями эти хромосомы не столь конденсированы и в большей степени расплетены. Понятно, что в конденсированном состоянии им трудно функционировать. Хромосома имеет вид в виде буквы Х только во время одной из стадий митоза. Раньше считалось, что между делениями клетки хромосомная ДНК (хроматин) находится в полностью расплетенном состоянии, но сейчас выясняется, что структура хромосом достаточно сложная и степень деконденсации хроматина между делениями не очень велика. 2.Непрямое деление клетки. Амитоз. Эндомитоз Процесс деления, при котором исходно диплоидная клетка дает две дочерние, также диплоидные, клетки, называется митозом. Митоз условно разделяют на четыре фазы: профазу, метафазу, анафазу и телофазу. Профаза. Две центриоли начинают расходиться к противоположным полюсам ядра. Ядерная мембрана разрушается; одновременно специальные белки объединяются, формируя микротрубочки в виде нитей. Центриоли, расположенные теперь на противоположных полюсах клетки, оказывают организующее воздействие на микротрубочки, которые в результате выстраиваются радиально, образуя структуру, напоминающую по внешнему виду цветок астры («звезда»). Другие нити из микротрубочек протягиваются от одной центриоли к другой, образуя веретено деления. В это время хромосомы спирализуются и вследствие этого утолщаются. Они хорошо видны в световом микроскопе, особенно после окрашивания. Считывание генетической информации с молекул ДНК становится невозможным: синтез РНК прекращается, ядрышко исчезает. В профазе хромосомы расщепляются, но хроматиды все еще остаются скрепленными попарно в зоне центромеры. Центромеры тоже оказывают организующее воздействие на нити веретена, которые теперь тянутся от центриоли к центромере и от нее к другой центриоли. Метафаза. В метафазе спирализация хромосом достигает максимума, и укороченные хромосомы устремляются к экватору клетки, располагаясь на равном расстоянии от полюсов. Образуется экваториальная, или метафазная, пластинка. На этой стадии митоза отчетливо видна структура хромосом, их легко сосчитать и изучить их индивидуальные особенности. В каждой хромосоме имеется область первичной перетяжки — центромера, к которой во время митоза присоединяются нить веретена деления и плечи. На стадии метафазы хромосома состоит из двух хроматид, соединенных между собой только в области центромеры. В анафазе вязкость цитоплазмы уменьшается, центромеры разъединяются, и с этого момента хроматиды становятся самостоятельными хромосомами. Нити веретена деления, прикрепленные к центромерам, тянут хромосомы к полюсам клетки, а плечи хромосом при этом пассивно следуют за центромерой. Таким образом, в анафазе хроматиды удвоенных еще в интерфазе хромосом точно расходятся к полюсам клетки. В этот момент в клетке находятся два диплоидных набора хромосом (4n4с). Таблица 1. Митотический цикл и митоз
В телофазе хромосомы раскручиваются, деспирализуются. Из мембранных структур цитоплазмы образуется ядерная оболочка. В это время восстанавливается ядрышко. На этом завершается деление ядра (кариокинез), затем происходит деление тела клетки (или цитокинез). При делении животных клеток на их поверхности в плоскости экватора появляется борозда, постепенно углубляющаяся и разделяющая клетку на две половины - дочерние клетки, в каждой их которых имеется по ядру. У растений деление происходит путем образования так называемой клеточной пластинки, разделяющей цитоплазму: она возникает в экваториальной области веретена, а затем растет во все стороны, достигая клеточной стенки (т.е. растет изнутри кнаружи). Клеточная пластинка формируется из материала, поставляемого эндоплазматической сетью. Затем каждая из дочерних клеток образует на своей стороне клеточную мембрану и, наконец, на обеих сторонах пластинки образуются целлюлозные клеточные стенки. Особенности протекания митоза у животных и растений приведены в таблице 2. Таблица 2. Особенности митоза у растений и у животных
Так из одной клетки формируются две дочерние, в которых наследственная информация точно копирует информацию, содержавшуюся в материнской клетке. Начиная с первого митотического деления оплодотворенной яйцеклетки (зиготы) все дочерние клетки, образовавшиеся в результате митоза, содержат одинаковый набор хромосом и одни и те же гены. Следовательно, митоз - это способ деления клеток, заключающийся в точном распределении генетического материала между дочерними клетками. В результате митоза обе дочерние клетки получают диплоидный набор хромосом. Весь процесс митоза занимает в большинстве случаев от 1 до 2 часов. Частота митоза в разных тканях и у разных видов различна. Например, в красном костном мозге человека, где каждую секунду образуется 10 млн эритроцитов, в каждую секунду должно происходить 10 млн. митозов. А в нервной ткани митозы крайне редки: так, в центральной нервной системе клетки в основном перестают делиться уже в первые месяцы после рождения; а в красном костном мозге, в эпителиальной выстилке пищеварительного тракта и в эпителии почечных канальцев они делятся до конца жизни. Регуляция митоза, вопрос о пусковом механизме митоза. Факторы, побуждающие клетку к митозу точно не известны. Но полагают, что большую роль играет фактор соотношения объемов ядра и цитоплазмы (ядерно-плазменное соотношение). По некоторым данным, отмирающие клетки продуцируют вещества, способные стимулировать деление клетки. Белковые факторы, отвечающие за переход в фазу М, первоначально были идентифицированы на основе экспериментов по слиянию клеток. Слияние клетки, находящейся в любой стадии клеточного цикла, с клеткой находящейся в М фазе, приводит к вхождению ядра первой клетки в М фазу. Это означает, что в клетке находящейся в М фазе существует цитоплазматический фактор способный активировать М фазу. Позднее этот фактор был вторично обнаружен в экспериментах по переносу цитоплазмы между ооцитами лягушки, находящимися на различных стадиях развития, и был назван "фактором созревания" MPF (maturation promoting factor). Дальнейшее изучение MPF показало, что этот белковый комплекс детерминирует все события М-фазы. На рисунке показано, что распад ядерной мембраны, конденсация хромосом, сборка веретена, цитокинез регулируются MPF. Митоз тормозится высокой температурой, высокими дозами ионизирующей радиации, действием растительных ядов. Один из таких ядов называется колхицин. С его помощью можно остановить митоз на стадии метафазной пластинки, что позволяет подсчитать число хромосом и дать каждой из них индивидуальную характеристику, т. е. провести кариотипирование. Амитоз (от греч. а – отриц. частица и митоз) - прямое деление интерфазного ядра путем перешнуровывания без преобразования хромосом. При амитозе не происходит равномерное расхождение хроматид к полюсам. И это деление не обеспечивает образование генетически равноценных ядер и клеток. По сравнению с митозом амитоз более кратковременный и экономичный процесс. Амитотическое деление может осуществляться несколькими способами. Наиболее распространенный тип амитоза – это перешнуровывание ядра на две части. Этот процесс начинается с разделения ядрышка. Перетяжка углубляется, и ядро разделяется надвое. После этого начинается разделение цитоплазмы, однако это происходит не всегда. Если амитоз ограничивается только делением ядра, то это приводит к образованию дву- и многоядерных клеток. При амитозе может также происходить почкование и фрагментация ядер. Клетка, претерпевшая амитоз, в последующем не способна вступить в нормальный митотический цикл. Амитоз встречается в клетках различных тканей растений и животных. У растений амитотическое деление довольно часто встречается в эндосперме, в специализирующихся клетках корешков и в клетках запасающих тканей. Амитоз также наблюдается в высокоспециализированных клетках с ослабленной жизнеспособностью или дегенерирующих, при различных патологических процессах, таких как злокачественный рост, воспаление и т. п. Кроме митоза в клетках некоторых органов растений и животных встречаются и другие типы деления: эндомитоз и политения. При эндомитозе не формируется веретено деления и сохраняется ядерная оболочка, вследствие чего образуются полиплоидные клетки с увеличенным числом хромосом. Политения рассматривается как частный случай эндомитоза, поскольку после многократной репликации ДНК все хроматиновые нити (хроматиды) плотно прилегают друг к другу и соединены общей центромерой, образуя гигантские политенные хромосомы.  3. Мейоз и его значение При образовании гамет, т.е. половых клеток – сперматозоидов и яйцеклеток – происходит деление клетки, называемое мейозом. Мейоз (от греч. meiosis – уменьшение) - это особый способ деления клеток, в результате которого происходит редукция (уменьшение) числа хромосом и переход клеток из диплоидного состояния 2n в гаплоидное n. Этот вид деления был впервые описан В. Флемингом в 1882 г. у животных и Э. Страсбургером в 1888 г. у растений. Мейоз включает два последовательных деления: первое (редукционное) и второе (эквационное). В каждом делении выделяют 4 фазы: профаза, метафаза, анафаза, телофаза. Все фазы первого мейотического деления обозначают цифрой I, а все фазы второго деления — цифрой II. Мейозу предшествует интерфаза, в процессе которой происходит удвоение ДНК и клетки вступают в мейоз с хромосомным набором 2n4с (n — хромосомы, с — хроматиды). Профаза I мейоза отличается значительной продолжительностью и сложностью. Ее условно разделяют на пять последовательных стадий: лептотена, зиготена, пахитена, диплотена и диакинез. Каждая из этих стадий обладает своими отличительными особенностями. Лептотена (стадия тонких нитей). Для этой стадии характерно наличие тонких и длинных хромосомных нитей. Число хромосомных нитей соответствует диплоидному числу хромосом. Каждая хромосомная нить состоит из двух хроматид, соединенных общим участком — центромерой. Хроматиды очень близко сближены, и поэтому каждая хромосома кажется одиночной. Зиготена (стадия соединения нитей). Моментом перехода лептотены в зиготену считают начало синапса. Синапс – процесс тесной конъюгации двух гомологичных хромосом. Подобная конъюгация отличается высокой точностью. Конъюгация часто начинается с того, что гомологичные концы двух хромосом сближаются на ядерной мембране, а затем процесс соединения гомологов распространяется вдоль хромосом от обоих концов. В других случаях синапс может начаться во внутренних участках хромосом и продолжаться по направлению к их концам. В результате каждый ген входит с соприкосновение с гомологичным ему геном той же хромосомы. Такой тесный контакт между гомологичными участками хроматид обеспечивается благодаря специализированной структуре – синаптонемальному комплексу. Синаптонемальный комплекс представляет собой длинное белковое образование, напоминающее веревочную лестницу, к противоположным сторонам которого плотно прилегают два гомолога. Пахитена (стадия толстых нитей). Как только завершается синапс по всей длине хромосом, клетки вступают в стадию пахитены, на которой они могут оставаться несколько суток. Соединение гомологов становится столь тесным, что уже трудно отличить две отдельные хромосомы. Однако это пары хромосом, которые называют бивалентами. В этой стадии происходит кроссинговер, или перекрест хромосом. Кроссинговер (от англ. crossingover - пересечение, скрещивание) - взаимный обмен гомологичными участками гомологичных хромосом. В результате кроссинговера хромосомы несут комбинации генов в новом сочетании. Например, ребенок родителей, один из которых имеет темные волосы и карие глаза, а другой - светловолосый и голубоглазый, может иметь карие глаза и светлые волосы. Диплотена (стадия двойных нитей). Стадия диплотены начинается с разделения конъюгировавших хромосом. Процесс отталкивания начинается в области центромеры и распространяется к концам. В это время хорошо видно, что бивалент состоит из двух хромосом (откуда и название стадии «двойные нити»), и что каждая хромосома состоит из двух хроматид. Всего в биваленте структурно обособлены четыре хроматиды, поэтому бивалент называют тетрадой. В это же время становится видно, что тела двух гомологичных хромосом переплетаются. Фигуры перекрещенных хромосом напоминают греческую букву «хи» (χ), поэтому места перекреста назвали хиазмами. Наличие хиазм связано с произошедшим кроссинговером. По мере прохождения этой стадии хромосомы как бы раскручиваются, происходит перемещение хиазм от центра к концам хромосом (терминализация хиазм). Это обеспечивает возможность движения хромосом к полюсам в анафазе. Диакинез. Диплотена незаметно переходит в диакинез, завершающую стадию профазы I. На этой стадии биваленты, которые заполняли весь объем ядра, начинают перемещаться ближе к ядерной оболочке. К концу диакинеза контакт между хроматидами сохраняется на одном или обоих концах. Исчезновение оболочки ядра и ядрышек, а также окончательное формирование веретена деления завершают профазу I. Метафаза I. В метафазе I биваленты располагаются в экваториальной плоскости клетки. Нити веретена прикрепляются к центромерам гомологичных хромосом. Анафаза I. В анафазе I к полюсам отходят не хроматиды, как при митозе, а гомологичные хромосомы из каждого бивалента. В этом принципиальное отличие мейоза от митоза. При этом расхождение гомологичных хромосом носит случайный характер. Телофаза I очень короткая, в процессе ее идет формирование новых ядер. Хромосомы деконденсируются и деспирализуются. Так заканчивается редукционное деление, и клетка переходит в короткую интерфазу, после которой наступает второе мейотическое деление. От обычной интерфазы эта интерфаза отличается тем, что в ней не происходит синтеза ДНК и дупликации хромосом, хотя синтез РНК, белка и других веществ может происходить. Цитокинез у многих организмов происходит не сразу после деления ядер, так что в одной клетке лежат два ядра более мелких, чем исходное. Затем наступает второе деление мейоза, сходное с обычным митозом. Профаза II очень короткая. Она характеризуется спирализацией хромосом, исчезновением ядерной оболочки, ядрышка, формированием веретена деления. Метафаза II. Хромосомы располагаются в экваториальной плоскости. Центромеры, соединяющие пары хроматид, делятся (в первый и единственный раз в течение мейоза), что свидетельствует о начале анафазы II. В анафазе II хроматиды расходятся и быстро увлекаются нитями веретена от плоскости экватора к противоположным полюсам. Телофаза II. Для этой стадии характерно деспирализация хромосом, образование ядер, цитокинез. В итоге из двух клеток мейоза I в телофазе II образуются четыре клетки с гаплоидным числом хромосом. Описанный процесс типичен для образования мужских половых клеток. Образование женских половых клеток идет аналогично, но при овогенезе развивается лишь одна яйцеклетка, а три мелких направительных (редукционных) тельца впоследствии отмирают. Направительные тельца несут полноценные хромосомные наборы, но практически лишены цитоплазмы и вскоре погибают. Биологический смысл образования этих телец заключается в необходимости сохранения в цитоплазме яйцеклетки максимального количества желтка, потребного для развития будущего зародыша. Таким образом, для мейоза характерно два деления: в ходе первого расходятся хромосомы, в ходе второго - хроматиды. Разновидности мейоза. В зависимости от места в жизненном цикле организма выделяют три основных типа мейоза: зиготный, или начальный, споровый, или промежуточный, гаметный, или конечный. Зиготный тип происходит в зиготе сразу после оплодотворения и приводит к образованию гаплоидного мицелия или таллома, а затем спор и гамет. Этот тип характерен для многих грибов и водорослей. У высших растений наблюдается споровый тип мейоза, который проходит перед цветением и приводит к образованию гаплоидного гаметофита. Позднее в гаметофите образуются гаметы. Для всех многоклеточных животных и ряда низших растений свойственен гаметный, или конечный, тип мейоза. Протекает он в половых органах и приводит к образованию гамет. Биологическое значение мейоза заключается в том, что: поддерживается постоянный кариотип в ряду поколений организмов, размножающихся половым путем (после оплодотворения образуется зигота, содержащая характерный для данного вида набор хромосом). обеспечивается перекомбинация генетического материала как на уровне целых хромосом (новые комбинации хромосом), так и на уровне участков хромосом. 4. Краткий обзор этапов гаметогенезаГаметогенез подразделяется на сперматогенез (процесс образования сперматозоидов у самцов) и оогенез (процесс образования яйцеклетки). По тому, что происходит с ДНК, эти процессы практически не отличаются: одна исходная диплоидная клетка дает четыре гаплоидные. Однако, по тому, что происходит с цитоплазмой, эти процессы кардинально различаются. В яйцеклетке накапливаются питательные вещества, необходимые в дальнейшем для развития зародыша, поэтому яйцеклетка – это очень крупная клетка, и когда она делится, цель – сохранить питательные вещества для будущего зародыша, поэтому деление цитоплазмы несимметрично. Для того чтобы сохранить все запасы цитоплазмы и при этом избавиться от ненужного генетического материала, от цитоплазмы отделяются полярные тельца, которые содержат очень мало цитоплазмы, но позволяют поделить хромосомный набор. Полярные тельца отделяются при первом и втором делении мейоза. Исходная клетка, из которой в последствии образуется зрелая яйцеклетка, называется ооцитом первого порядка. После деления из него образуется ооцит второго порядка и первое полярное тельце. Затем происходит второе деление мейоза, в результате образуется гаплоидный оотид и второе полярное тельце. Первое полярное тельце за это время тоже успевает поделиться, таким образом, всего получается три гаплоидных полярных тельца. В оотиде происходят некоторые процессы созревания и он превращается в яйцеклетку. Она содержащая почти всю цитоплазму исходного ооцита, но гаплоидный набор хромосом. Эти хромосомы уже прошли рекомбинацию, т.е. если исходно клетки содержат одну хромосому от мамы, одну от папы, то в зрелой яйцеклетке в каждой хромосоме чередуются куски, полученные от одного и второго родителя. При сперматогенезе цитоплазма исходного сперматоцита первого порядка делится (первое деление мейоза) поровну между клетками, давая сперматоциты второго порядка. Второе деление мейоза приводит к образованию гаплоидных сперматоцитов второго порядка. Затем происходит созревание без деления клетки, большая часть цитоплазмы отбрасывается, и получаются сперматозоиды, содержащие гаплоидный набор хромосом очень мало цитоплазмы. Оплодотворенное яйцо называют зиготой (от греч. зиготос – соединенный вместе). Амфимиксис – обычный тип полового процесса, при котором происходит слияние ядер женских и мужских гаплоидных гамет и образование диплоидной зиготы, из которой развивается зародыш. После оплодотворения происходит деление клетки, восстановившей диплоидный набор хромосом, первое и несколько последующих делений яйцеклетки происходят без увеличения размера клеток, поэтому процесс называется дроблением яйцеклетки.  Нерегулярные типы полового размножения. Апомиксис – развитие зародыша нового организма без слияния половых клеток (гамет): партеногенез – развитие яйцеклетки без оплодотворения: гиногенез – из неоплодотворенной яйцеклетки, андрогенез – из ядер спермиев, апогаметия (апогамия) – из других клеток женского гаметофита (синергид, антипод); апоспория – из нередуцированных соматических клеток спорофита или материнской споры; адвентивная эмбриония – из соматических клеток нуцеллуса или внутреннего интегумента семяпочек. Процесс мейоза лежит в основе полового размножения, поскольку приводит к гаплоидному числу хромосом в гаметах. У диплоидных организмов генетическая информация хранится в парных гомологичных хромосомах, причем один гомолог происходит от матери, а другой – от отца. В результате мейоза гаплоидные гаметы содержат как материнские, так и отцовские хромосомы. Благодаря кроссинговеру между этими гомологами в профазе I мейоза, генетическая изменчивость гамет становится более высокой. Особенно важную роль играет мейоз в жизненном цикле грибов и растений. У многих грибов (например, дрожжей) преобладающей фазой жизненного цикла считаются вегетативные гаплоидные клетки, которые размножаются путем митоза. У многоклеточных растений чередуются диплоидная стадия спорофита и гаплоидная стадия гаметофита. У разных систематических групп преобладает та или иная стадия, а мейоз с последующим оплодотворением яйцеклетки служит «мостом», соединяющим поколения спорофитов и гаметофитов. При половом размножении в жизненном цикле высших растений и животных преобладает диплоидная стадия (спорофит), гаплоидное состояние (гаметофит) свойственно только половым клеткам. Литература Айала, Ф. Современная генетика / Ф. Айала, Дж. Кайгер. – М.: Мир, 1987. – Т.1. – 295 с; Т.2. – 368 с; Т.3. Алиханян, С. И. Общая генетика / С. И. Алиханян, А. П. Акифьев, Л. С. Чернин.– М.: Высш. шк., 1985. Бокуть, С. Б. Молекулярная биология: молекулярные механизмы хранениия, воспроизведения и реализации генетической информации / С. Б. Бокуть, Н. В. Герасимович, А. А. Милютин. – Мн.:Высш. шк., 2005. Дубинин, Н. П. Общая генетика / Н. П. Дубинин. – М.: Наука, 1986. Жимулев, И. Ф. Общая и молекулярная генетика / И. Ф. Жимулев. –Новосибирск: Изд-во Новосибирского ун-та, 2002. Жученко, А. А. Генетика / А. А Жученко, Ю. Л. Гужов, В. А. Пухальский. – М.: Колос, 2004. | |||||||||||||||||||||||