Лекция 1 Анатомия и физиология женской половой системы
 Скачать 273.66 Kb. Скачать 273.66 Kb.
|

|
Лекция №1 Анатомия и физиология женской половой системы Женская репродуктивная система предназначена для выполнения нескольких функций. Она производит женские яйцеклетки, необходимые для размножения, которые называются зрелыми яйцеклетками или ооцитами (незрелые яйцеклетки). Система транспортирует зрелые яйцеклетки на место их оплодотворения. Оплодотворение, под которым имеется в виду оплодотворение яйцеклетки сперматозоидом, обычно происходит в фаллопиевых трубах. Следующим шагом для оплодотворенного яйца является его вживление в стенку матки, что является началом беременности. Если оплодотворения и/или вживления не происходит, система менструирует (ежемесячный выброс выстилающей ткани матки). Кроме этого, женская репродуктивная система производит женские половые гормоны, которые поддерживают репродуктивный цикл. Женская репродуктивная система включает в себя как внутренние, так и наружные органы. Функции внешних женских репродуктивных органов (гениталий) двояки: сделать возможным проникновение спермы в тело и защитить внутренние половые органы от попадания инфекций. Основные наружные половые органы женской репродуктивной системы включают в себя: Большие половые губы: это кожные складки, состоящие из подкожно-жировой ткани, содержат потовые и сальные железы. Большие половые губы имеют потовые и сальные железы. В задней спайке имеются большие сальные железы – Бартолиновые. Они выделяют секрет, который увлажняют вход во влагалище. У Бартолиновых желез выводной проток располагают в преддверии влагалища, поэтому они первыми реагируют на проникновение инфекции. Функция больших половых губ защитная, они прикрывают малые половые губы. После наступления пубертатного возраста, большие половые губы покрыты волосами. Малые половые губы: это кожные складки, обеднены подкожно жировой клетчаткой, но богаты сосудами и нервными окончаниями. Имеют гладкий синюшно-красный цвет. Они находятся внутри больших половых губ и окружают вход во влагалище (канал, который соединяет нижнюю часть матки с наружной стороной тела) и уретрой (трубка, по которой моча выходит из мочевого пузыря наружу). Малые половые губы находятся всегда в сомкнутом состоянии, функция их защитная – защищает уретру от инфекции. Клитор: это рудимент полового члена. Различают: головку, тело и ножки клитора (малые половые губы). Тело клитора также состоит из губчатого тела и пещеристого вещества. Головка содержит сосуды и нервные окончания, от чего орган чувствителен к прикосновению. А при акушерских травмах дает массивное кровотечение. При местном обезболивании достигается незначительный анестезиологический эффект, что затрудняет эффективно провести гемостаз. Внутренние женские репродуктивные органы включают в себя: Влагалище: это фиброзно-мышечная трубка. Располагающая между мочеиспускательным каналом и прямой кишкой, длиной 7-8 см. Вокруг шейки матки влагалище плотно спаяно и образуются своды: передний, задний, и два боковых. Задний свод очень глубокий, что позволяет скапливаться сперме и секрету из матки. Слизистая влагалища представлена многослойным плоским эпителием, она образует поперечные складки, которые с возрастом сглаживаются. Флора влагалища палочковая. Палочки (Додерляйна) – это лактабактерии, которые продуцируют молочную кислоту, которая является барьером для проникновения инфекции. По количеству лактабактерий различают чистоту влагалища: I, II, III, IV. I и II степени чистоты – здоровая женщина, преоблодают во влагалищном мазке палочки, патогенной флоры нет. III и IVстепени чистоты – требует санации влагалища, т.к. вместо палочек, флора кокковая или отсутствует. Присоединяется патогенная инфекция: грибы, трихомонады и т.д. Среда во влагалище слабо кислая, это благоприятно сперматозоидов. Матка: это полый орган грушевидной формы, который является местом развития плода, длиной 7-8 см. Располагается в малом тазу между мочевым пузырем и прямой кишкой. Имеет физиологический перегиб в месте перешейка кпереди (к мочевому пузырю). Различают: тело матки, дно, переднюю и заднюю стенки, два ребра, шейка и перешеек. В шейке матки различают влагалищную и над влагалищную части. Сообщаются они между собой посредством цервикального канала, длиною до 1,2 см. Канал, проходящий через шейку матки, позволяет сперме попадать внутрь, а менструальной крови – выходить наружу. Стенка матки представлена: слизистый слой – эндометрий, состоит из функционального и базального слоев. Из них базальный остается неизменным, функциональный обновляется ежемесячно. Средний слой – мышечный – миометрий, представлен поперечными и круговыми мышцами, что позволяет ритмично сокращаться матке в родовую деятельность. Покрыта матка собственной брюшиной - периметрием. Маточные трубы: это парные органы, которые отходят от углов матки, длиной 12 см и служат яйцеводом. Различаю: маточный отдел (истмический), суженный (истмикоцервикальный), расширенный (ампулярный). Ампулярный отдел заканчивается бахромками (фимбриями), которые находятся все время в движении, они создают турбулентное течение, что не позволяет яйцеклетке уйти в брюшную полость, а попадать только в просвет маточной трубы. Стенки маточной трубы представлены тремя слоями: 1) слизистая – цилиндрический эпителий – мерцательный; 2) средний слой – круговые мышцы; 3) собственная брюшина. Оплодотворенное яйцо движется в матку, где оно вживляется в выстилающую матку ткань. Яичники: это парные половые органы женщины, размером с миндаль, одновременно с внешней и внутренней секрецией: вырабатывает женские половые клетки – яйцеклетки, и половые гормоны – эстрагены и прогестероны. Состоят яичники из мозгового слоя, где проходят сосуды и нервы, корковый слой, где располагаются преморбитные и доминантный фолликулы, желтое тело. Белочная оболочка и собственная брющина. В женских железах - яичниках - синтезируются эстрогены, а также в небольшом количестве тестостерон, который является предшественником эстрогенов. Прогестерон, женский половой гормон, синтезируется желтым телом яичника, образующимся и осуществляющим свою функциональную активность после наступления овуляции.Женские половые гормоны – эстрогены (эстрол, эстриол и эстрадиол) выступают регуляторами овариально-менструального цикла, а при наступлении беременности – регуляторами ее нормального течения. Эстрогены оказывают влияние на: развитие половых органов; выработку яйцеклеток; обуславливают подготовку яйцеклеток к оплодотворению, матки – к беременности, молочных желез – к кормлению ребенка; регулируют формирование женской фигуры и особенностей скелета; обеспечивают внутриутробное развитие на всех этапах. Кроме этого эстрогены увеличивают синтез гликогена в печени и отложение жира в организме. Эстрогены, попадая из яичников в кровь, транспортируются по организму при помощи белков-переносчиков. Разрушаются эстрогены в печени при помощи печеночных ферментов и выводятся из организма с мочой. Прогестерон или гормон желтого тела синтезируется в яичниках и в плаценте при беременности. Он помогает сохранению беременности, подготавливает внутреннюю слизистую оболочку матки для имплантации в нее оплодотворенной яйцеклетки, подавляет действие эстрогенов и сокращение матки, способствует развитию железистой ткани молочных желез, под его воздействием повышается базальная температура. Разрушается прогестерон в печени и выводится с мочой. Кроме этого в яичниках вырабатывается некоторое количество андрогенов. Менструальный цикл - генетически детерминированные, циклически повторяющиеся изменения в организме женщины, особенно в звеньях репродуктивной системы, клиническим проявлением которых служат кровяные выделения из половых путей (менструация). Менструальный цикл устанавливается после менархе (первой менструации) и сохраняется в течение репродуктивного (детородного) периода жизни женщины до менопаузы (последней менструации). Циклические изменения в организме женщины направлены на возможность воспроизводства потомства и носят двухфазный характер: 1-я (фолликулярная) фаза цикла определяется ростом и созреванием фолликула и яйцеклетки в яичнике, после чего происходят разрыв фолликула и выход из него яйцеклетки - овуляция; 2-я (лютеиновая) фаза связана с образованием желтого тела. Одновременно в циклическом режиме происходят последовательные изменения в эндометрии: регенерация и пролиферация функционального слоя, сменяющаяся секреторной трансформацией желез. Изменения в эндометрии заканчиваются десквамацией функционального слоя (менструацией). Биологическое значение изменений, которые происходят на протяжении менструального цикла в яичниках и эндометрии, состоит в обеспечении репродуктивной функции после созревания яйцеклетки, ее оплодотворения и имплантации зародыша в матке. Если оплодотворения яйцеклетки не происходит, функциональный слой эндометрия отторгается, из половых путей появляются кровяные выделения, а в репродуктивной системе вновь и в той же последовательности происходят процессы, направленные на обеспечение созревания яйцеклетки. Менструация - это повторяющиеся с определенными интервалами кровяные выделения из половых путей на протяжении всего репродуктивного периода, исключая беременность и лактацию. Менструация начинается в конце лютеиновой фазы менструального цикла в результате отторжения функционального слоя эндометрия. Первая менструация (menarhe) возникает в возрасте 10-12 лет. В течение последующих 1-1,5 лет менструации могут быть нерегулярными, и лишь затем устанавливается регулярный менструальный цикл. Первый день менструации условно принимается за 1-й день менструального цикла, а продолжительность цикла рассчитывается как интервал между первыми днями двух следующих одна за другой менструаций. Внешние параметры нормального менструального цикла: • длительность - от 21 до 35 дней (у 60% женщин средняя продолжительность цикла составляет 28 дней); • продолжительность менструальных выделений - от 3 до 7 дней; • величина кровопотери в менструальные дни - 40-60 мл (в среднем 50 мл). Процессы, обеспечивающие нормальное течение менструального цикла, регулируются единой функционально-связанной нейроэндокринной системой, включающей центральные (интегрирующие) отделы, периферические (эффекторные) структуры, а также промежуточные звенья. Функционирование репродуктивной системы обеспечивается строго генетически запрограммированным взаимодействием пяти основных уровней, каждый из которых регулируется вышележащими структурами по принципу прямой и обратной, положительной и отрицательной взаимосвязи (рис. 1). 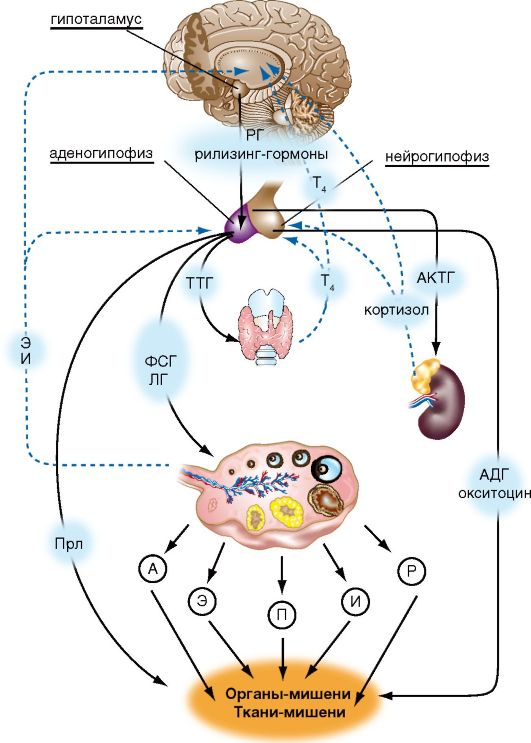 Рис. 1 Гормональная регуляция в системе гипоталамус - гипофиз - периферические эндокринные железы - органы мишени (схема): РГ - рилизинг-гормоны; ТТГ - тиреотропный гормон; АКТГ - адренокотикотропный гормон; ФСГ - фолликулостимулирующий гормон; ЛГ - лютеинизирующий гормон; Прл - пролактин; П - прогестерон; Э - эстрогены; А - андрогены; Р - релаксин; И - ингибин; Т4 - тироксин, АДГ - антидиуретический гормон (вазопрессин). Первым (высшим) уровнем регуляции репродуктивной системы являются кора головного мозга и экстрагипоталамические церебральные структуры. Адекватное состояние ЦНС обеспечивает нормальное функционирование всех нижележащих звеньев репродуктивной системы. Различные органические и функциональные изменения в коре и подкорковых структурах могут приводить к нарушениям менструального цикла. Хорошо известна возможность прекращения менструаций при сильных стрессах (потеря близких людей, условия военного времени и т.д.) или без очевидных внешних воздействий при общей психической неуравновешенности ("ложная беременность" - задержка менструации при сильном желании беременности или, наоборот, при ее боязни). Специфические нейроны головного мозга получают информацию о состоянии как внешней, так и внутренней среды. Внутреннее воздействие осуществляется с помощью специфических рецепторов к стероидным гормонам яичников (эстрогенам, прогестерону, андрогенам), находящимся в ЦНС. В ответ на воздействие факторов внешней среды на кору головного мозга и экстрагипоталамические структуры происходят синтез, выделение и метаболизм нейротрансмиттеров и нейропептидов. В свою очередь, ней-ротрансмиттеры и нейропептиды влияют на синтез и выделение гормонов нейросекреторными ядрами гипоталамуса. К важнейшим нейротрансмиттерам, т.е. веществам-передатчикам нервных импульсов, относятся норадреналин, дофамин, γ-аминомасляная кислота (ГАМК), ацетилхолин, серотонин и мелатонин. Норадреналин, аце-тилхолин и ГАМК стимулируют выброс гонадотропного рилизинг-гормона (ГнРГ) гипоталамусом. Дофамин и серотонин уменьшают частоту и снижают амплитуду выработки ГнРГ в течение менструального цикла. Нейропептиды (эндогенные опиоидные пептиды, нейропептид Y, га-ланин) также участвуют в регуляции функции репродуктивной системы. Опиоидные пептиды (эндорфины, энкефалины, динорфины), связываясь с опиатными рецепторами, приводят к подавлению синтеза ГнРГ в гипоталамусе. Вторым уровнем регуляции репродуктивной функции является гипоталамус. Несмотря на малые размеры, гипоталамус участвует в регуляции полового поведения, осуществляет контроль за вегетососудистыми реакциями, температурой тела и другими жизненно важными функциями организма. Гипофизотропная зона гипоталамуса представлена группами нейронов, составляющих нейросекреторные ядра: вентромедиальное, дорсомедиаль-ное, аркуатное, супраоптическое, паравентрикулярное. Эти клетки имеют свойства как нейронов (воспроизводящие электрические импульсы), так и эндокринных клеток, вырабатывающих специфические нейросекреты с диаметрально противоположными эффектами (либерины и статины). Ли-берины, или рилизинг-факторы, стимулируют освобождение соответствующих тропных гормонов в передней доле гипофиза. Статины оказывают ингибирующее действие на их выделение. В настоящее время известны семь либеринов, по своей природе являющихся декапептидами: тиреолиберин, кортиколиберин, соматолиберин, меланолиберин, фоллиберин, люлибе-рин, пролактолиберин, а также три статина: меланостатин, соматостатин, пролактостатин, или пролактинингибирующий фактор. Люлиберин, или рилизинг-гормон лютеинизирующего гормона (РГЛГ), выделен, синтезирован и подробно описан. Выделить и синтезировать фолликулостимулирующий рилизинг-гормон до настоящего времени не удалось. Однако установлено, что РГЛГ и его синтетические аналоги стимулируют выделение гонадотрофами не только ЛГ, но и ФСГ. В связи с этим принят один термин для гонадотропных либеринов - "гонадотропин-рилизинг-гормон" (ГнРГ), по сути, являющийся синонимом люлиберина (РГЛГ). Основное место секреции ГнРГ - аркуатные, супраоптические и пара-вентрикулярные ядра гипоталамуса. Аркуатные ядра воспроизводят секреторный сигнал с частотой приблизительно 1 импульс в 1-3 ч, т.е. в пульсирующем или цирхоральном режиме (circhoral - вокруг часа). Эти импульсы имеют определенную амплитуду и вызывают периодическое поступление ГнРГ через портальную систему кровотока к клеткам аденогипофиза. В зависимости от частоты и амплитуды импульсов ГнРГ в аденогипофизе происходит преимущественная секреция ЛГ или ФСГ, что, в свою очередь, вызывает морфологические и секреторные изменения в яичниках. Гипоталамо-гипофизарная область имеет особую сосудистую сеть, которая называется портальной системой. Особенностью данной сосудистой сети является возможность передачи информации как от гипоталамуса к гипофизу, так и обратно (от гипофиза к гипоталамусу). Регуляция выделения пролактина в большей степени находится под статиновым влиянием. Дофамин, образующийся в гипоталамусе, тормозит высвобождение пролактина из лактотрофов аденогипофиза. Увеличению секреции пролактина способствуют тиреолиберин, а также серотонин и эндогенные опиоидные пептиды. Кроме либеринов и статинов, в гипоталамусе (супраоптическом и пара-вентрикулярном ядрах) продуцируются два гормона: окситоцин и вазопрессин (антидиуретический гормон). Гранулы, содержащие данные гормоны, мигрируют от гипоталамуса по аксонам крупноклеточных нейронов и накапливаются в задней доле гипофиза (нейрогипофиз). Третьим уровнем регуляции репродуктивной функции является гипофиз, он состоит из передней, задней и промежуточной (средней) доли. Непосредственное отношение к регуляции репродуктивной функции имеет передняя доля (аденогипофиз). Под воздействием гипоталамуса в аденогипофизе секретируются гонадотропные гормоны - ФСГ (или фоллитропин), ЛГ (или лютропин), пролактин (Прл), АКТГ, соматотропный (СТГ) и тиреотропный (ТТГ) гормоны. Нормальное функционирование репродуктивной системы возможно лишь при сбалансированном выделении каждого из них. Гонадотропные гормоны (ФСГ, ЛГ) передней доли гипофиза находятся под контролем ГнРГ, который стимулирует их секрецию и высвобождение в кровеносное русло. Пульсирующий характер секреции ФСГ, ЛГ является следствием "прямых сигналов" из гипоталамуса. Частота и амплитуда импульсов секреции ГнРГ меняется в зависимости от фаз менструального цикла и влияет на концентрацию и соотношение ФСГ/ЛГ в плазме крови. ФСГ стимулирует в яичнике рост фолликулов и созревание яйцеклетки, пролиферацию гранулезных клеток, образование рецепторов ФСГ и ЛГ на поверхности гранулезных клеток, активность ароматаз в зреющем фолликуле (это усиливает конверсию андрогенов в эстрогены), продукцию ингибина, активна и инсулиноподобных факторов роста. ЛГ способствует образованию андрогенов в текаклетках, обеспечивает овуляцию (совместно с ФСГ), стимулирует синтез прогестерона в лютеини-зированных клетках гранулезы (желтом теле) после овуляции. Пролактин оказывает многообразное действие на организм женщины. Его основная биологическая роль - стимуляция роста молочных желез, регуляция лактации; он также обладает жиромобилизующим и гипотензивным эффектом, осуществляет контроль секреции прогестерона желтым телом путем активации образования в нем рецепторов к ЛГ. Во время беременности и лактации уровень пролактина в крови увеличивается. Ги-перпролактинемия приводит к нарушению роста и созревания фолликулов в яичнике (ановуляции). Задняя доля гипофиза (нейрогипофиз) не является эндокринной железой, а лишь депонирует гормоны гипоталамуса (окситоцин и вазопрессин), которые находятся в организме в виде белкового комплекса. Яичники относятся к четвертому уровню регуляции репродуктивной системы и выполняют две основные функции. В яичниках происходят циклические рост и созревание фолликулов, созревание яйцеклетки, т.е. осуществляются генеративная функция, а также синтез половых стероидов (эстрогенов, андрогенов, прогестерона) - гормональная функция. Основной морфофункциональной единицей яичника является фолликул. При рождении в яичниках девочки находится примерно 2 млн примордиальных фолликулов. Основная их масса (99%) в течение жизни подвергается атрезии (обратное развитие фолликулов). Только очень небольшая их часть (300-400) проходит полный цикл развития - от примордиального до преовуляторного с образованием в последующем желтого тела. Ко времени менархе в яичниках содержится 200-400 тыс. примордиальных фолликулов. Яичниковый цикл состоит из двух фаз: фолликулярной и лютеиновой. Фолликулярная фаза начинается после менструации, связана с ростом и созреванием фолликулов и оканчивается овуляцией. Лютеиновая фаза занимает промежуток после овуляции до начала менструации и связана с образованием, развитием и регрессом желтого тела, клетки которого секретируют прогестерон. В зависимости от степени зрелости выделяют четыре типа фолликула: примордиальный, первичный (преантральный), вторичный (антральный) и зрелый (преовуляторный, доминантный) (рис. 2). 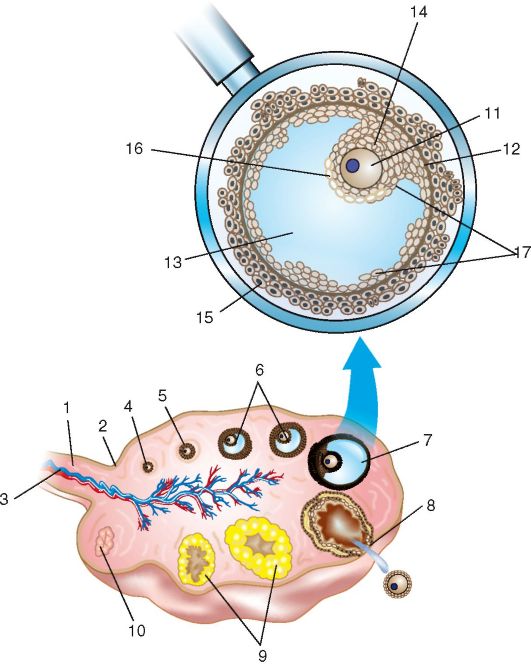 Рис. 2 Строение яичника (схема). Этапы развития доминантного фолликула и желтого тела: 1 - связка яичника; 2 - белочная оболочка; 3 - сосуды яичника (конечная ветвь яичниковой артерии и вены); 4 - примордиальный фолликул; 5 - преантральный фолликул; 6 - антральный фолликул; 7 - преовуляторный фолликул; 8 - овуляция; 9 - желтое тело; 10 - белое тело; 11 - яйцеклетка (ооцит); 12 - базальная мембрана; 13 - фолликулярная жидкость; 14 - яйценосный бугорок; 15 - тека-оболочка; 16 - блестящая оболочка; 17 - гранулезные клетки. Примордиальный фолликул состоит из незрелой яйцеклетки (ооцита) в профазе 2-го мейотического деления, которая окружена одним слоем гранулезных клеток. В преантральном (первичном) фолликуле ооцит увеличивается в размерах. Клетки гранулезного эпителия пролиферируют и округляются, образуя зернистый слой фолликула. Из окружающей стромы формируется соедительнотканная оболочка - тека (theca). Антральный (вторичный) фолликул характеризуется дальнейшим ростом: продолжается пролиферация клеток гранулезного слоя, которые продуцируют фолликулярную жидкость. Образующаяся жидкость оттесняет яйцеклетку к периферии, где клетки зернистого слоя образуют яйценосный бугорок (cumulus oophorus). Соединительнотканная оболочка фолликула отчетливо дифференцируется на наружную и внутреннюю. Внутренняя оболочка (the-ca interna) состоит из 2-4 слоев клеток. Наружная оболочка (theca externa) располагается над внутренней и представлена дифференцированной соединительнотканной стромой. В преовуляторном (доминантном) фолликуле яйцеклетка, находящаяся на яйценосном бугорке, покрыта мембраной, называемой блестящей оболочкой (zona pellucida). В ооците доминантного фолликула возобновляется процесс мейоза. За время созревания в преовуляторном фолликуле происходит стократное увеличение объема фолликулярной жидкости (диаметр фолликула достигает 20 мм). В течение каждого менструального цикла от 3 до 30 примордиальных фолликулов начинают расти, преобразуясь в преантральные (первичные) фолликулы. В последующий менструальный цикл продолжается фолликулогенез и только один фолликул развивается от преантрального до преовуляторного. В процессе роста фолликула от преантрального до антрального гранулезными клетками синтезируется антимюллеров гормон, способствующий его развитию. Остальные первоначально вступившие в рост фолликулы подвергаются атрезии (дегенерации). Овуляция - разрыв преовуляторного (доминантного) фолликула и выход из него яйцеклетки в брюшную полость. Овуляция сопровождается кровотечением из разрушенных капилляров, окружающих текаклетки. После выхода яйцеклетки в оставшуюся полость фолликула быстро врастают образующиеся капилляры. Гранулезные клетки подвергаются лютеинизации, морфологически проявляющейся в увеличении их объема и образовании липидных включений - формируется желтое тело. Желтое тело - транзиторное гормонально-активное образование, функционирующее в течение 14 дней независимо от общей продолжительности менструального цикла. Если беременность не наступила, желтое тело регрессирует, если же происходит оплодотворение, оно функционирует вплоть до образования плаценты (12-я неделя беременности). Гормональная функция яичников. Рост, созревание фолликулов в яичниках и образование желтого тела сопровождаются продукцией половых гормонов как гранулезными клетками фолликула, так и клетками внутренней теки и в меньшей степени - внешней теки. К половым стероидным гормонам относятся эстрогены, прогестерон и андрогены. Исходным материалом для образования всех стероидных гормонов служит холестерин. До 90% стероидных гормонов находятся в связанном состоянии и только 10% несвязанных гормонов оказывают свой биологический эффект. Эстрогены подразделяются на три фракции с различной активностью: эстрадиол, эстриол, эстрон. Эстрон - наименее активная фракция, выделяется яичниками в основном в период старения - в постменопаузе; наиболее активная фракция - эстрадиол, она значима в наступлении и сохранении беременности. Количество половых гормонов меняется на протяжении менструального цикла. По мере роста фолликула увеличивается синтез всех половых гормонов, но преимущественно эстрогенов. В период после овуляции и до начала менструации в яичниках преимущественно синтезируется прогестерон, выделяемый клетками желтого тела. Андрогены (андростендион и тестостерон) продуцируются текаклетками фолликула и межуточными клетками. Их уровень на протяжении менструального цикла не меняется. Попадая в клетки гранулезы, андрогены активно подвергаются ароматизации, приводящей к их конверсии в эстрогены. Помимо стероидных гормонов, яичники выделяют и другие биологически активные соединения: простагландины, окситоцин, вазопрессин, релаксин, эпидермальный фактор роста (ЭФР), инсулиноподобные факторы роста (ИПФР-1 и ИПФР-2). Полагают, что факторы роста способствуют пролиферации клеток гранулезы, росту и созреванию фолликула, селекции доминирующего фолликула. В процессе овуляции определенную роль играют простагландины (F2a и Е2), а также содержащиеся в фолликулярной жидкости протеолитические ферменты, коллагеназа, окситоцин, релаксин. Цикличность деятельности репродуктивной системы определяется принципами прямой и обратной связи, которая обеспечивается специфическими рецепторами к гормонам в каждом из звеньев. Прямая связь состоит в стимулирующем действии гипоталамуса на гипофиз и последующем образовании половых стероидов в яичнике. Обратная связь определяется влиянием повышенной концентрации половых стероидов на вышележащие уровни, блокируя их активность. Во взаимодействии звеньев репродуктивной системы различают "длинную", "короткую" и "ультракороткую" петли. "Длинная" петля - воздействие через рецепторы гипоталамо-гипофизарной системы на выработку половых гормонов. "Короткая" петля определяет связь между гипофизом и гипоталамусом, "ультракороткая" - связь между гипоталамусом и нервными клетками, которые под действием электрических стимулов осуществляют локальную регуляцию с помощью нейротрансмиттеров, нейропептидов, нейромодуляторов. Фолликулярная фаза. Пульсирующая секреция и выделение ГнРГ приводят к высвобождению ФСГ и ЛГ из передней доли гипофиза. ЛГ способствует синтезу андрогенов текаклетками фолликула. ФСГ воздействует на яичники и приводит к росту фолликула и созреванию ооцита. Вместе с этим увеличивающийся уровень ФСГ стимулирует продукцию эстрогенов в клетках гранулезы путем ароматизации андрогенов, образовавшихся в текаклетках фолликула, а также способствует секреции ингибина и ИПФР-1-2. Перед овуляцией увеличивается количество рецепторов к ФСГ и ЛГ в клетках теки и гранулезы (рис. 3). Овуляция происходит в середине менструального цикла, через 12-24 ч после достижения пика эстрадиола, вызывающего увеличение частоты и амплитуды секреции ГнРГ и резкий преовуляторный подъем секреции ЛГ по типу "положительной обратной связи". На этом фоне активизируются протеолитические ферменты - коллагеназа и плазмин, разрушающие коллаген стенки фолликула и таким образом уменьшающие ее прочность. Одновременно отмечаемое повышение концентрации простагландина F2a, а также окситоцина индуцирует разрыв фолликула в результате стимуляции ими сокращения гладких мышц и выталкивания ооцита с яйценосным бугорком из полости фолликула. Разрыву фолликула способствует также повышение в нем концентрации простагландина Е2 и релаксина, уменьшающих ригидность его стенок. Лютеиновая фаза. После овуляции уровень ЛГ снижается по отношению к "овуляторному пику". Однако данное количество ЛГ стимулирует процесс лютеинизации гранулезных клеток, оставшихся в фолликуле, а также преимущественную секрецию образовавшимся желтым телом прогестерона. Максимальная секреция прогестерона происходит на 6-8-й день существования желтого тела, что соответствует 20-22-му дню менструального цикла. Постепенно, к 28-30-му дню менструального цикла уровень прогестерона, эстрогенов, ЛГ и ФСГ снижается, желтое тело регрессирует и заменяется соединительной тканью (белое тело). Пятый уровень регуляции репродуктивной функции составляют чувствительные к колебаниям уровня половых стероидов органы-мишени: матка, маточные трубы, слизистая оболочка влагалища, а также молочные железы, волосяные фолликулы, кости, жировая ткань, ЦНС. Стероидные гормоны яичников влияют на обменные процессы в органах и тканях, имеющих специфические рецепторы. Эти рецепторы могут быть 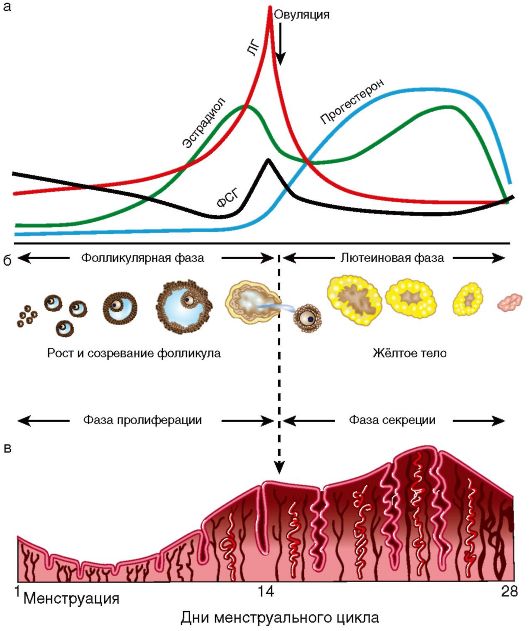 Рис. 3. Гормональная регуляция менструального цикла (схема): а - изменения уровня гормонов; б - изменения в яичнике; в - изменения в эндометрии как цитоплазматическими, так и ядерными. Цитоплазматические рецепторы строгоспецифичны к эстрогенам, прогестерону и тестостерону. Стероиды проникают в клетки-мишени, связываясь со специфическими рецепторами - соответственно к эстрогенам, прогестерону, тестостерону. Образовавшийся комплекс поступает в ядро клетки, где, соединяясь с хроматином, обеспечивает синтез специфических тканевых белков через транскрипцию матричной РНК. Матка состоит из наружного (серозного) покрова, миометрия и эндометрия. Эндометрий морфологически состоит из двух слоев: базального и функционального. Базальный слой в течение менструального цикла существенно не изменяется. Функциональный слой эндометрия претерпевает структурные и морфологические изменения, проявляющиеся последовательной сменой стадий пролиферации, секреции, десквамации с последующей регенерацией. Циклическая секреция половых гормонов (эстрогены, прогестерон) приводит к двухфазным изменениям эндометрия, направленным на восприятие оплодотворенной яйцеклетки. Циклические изменения в эндометрии касаются его функционального (поверхностного) слоя, состоящего из компактных эпителиальных клеток, которые отторгаются во время менструации. Базальный слой, не отторгаемый в этот период, обеспечивает восстановление функционального слоя. В эндометрии в течение менструального цикла происходят следующие изменения: десквамация и отторжение функционального слоя, регенерация, фаза пролиферации и фаза секреции. Трансформация эндометрия происходит под влиянием стероидных гормонов: фаза пролиферации - под преимущественным действием эстрогенов, фаза секреции - под влиянием прогестерона и эстрогенов. Фаза пролиферации (соответствует фолликулярной фазе в яичниках) продолжается в среднем 12-14 дней, начиная с 5-го дня цикла. В этот период образуется новый поверхностный слой с вытянутыми трубчатыми железами, выстланными цилиндрическим эпителием с повышенной митотической активностью. Толщина функционального слоя эндометрия составляет 8 мм. Фаза секреции (лютеиновая фаза в яичниках) связана с активностью желтого тела, длится 14±1 день. В этот период эпителий желез эндометрия начинает вырабатывать секрет, содержащий кислые гликозаминогликаны, гликопротеиды, гликоген Активность секреции становится наивысшей на 20-21-й день менструального цикла. К этому времени в эндометрии обнаруживается максимальное количество протеолитических ферментов, а в строме происходят децидуальные превращения. Отмечается резкая васкуляризация стромы - спиральные артерии функционального слоя извиты, образуют "клубки", вены расширены. Такие изменения в эндометрии, отмечаемые на 20-22-й день (6-8-й день после овуляции) 28-дневного менструального цикла, обеспечивают наилучшие условия для имплантации оплодотворенной яйцеклетки. К 24-27-му дню в связи с началом регресса желтого тела и снижением концентрации продуцируемого им прогестерона трофика эндометрия нарушается, постепенно в нем нарастают дегенеративные изменения. Из зернистых клеток стромы эндометрия выделяются гранулы, содержащие релаксин, подготавливающий менструальное отторжение слизистой оболочки. В поверхностных участках компактного слоя отмечаются лакунарные расширения капилляров и кровоизлияния в строму, что можно обнаружить за 1 сут до начала менструации. Менструация включает десквамацию, отторжение и регенерацию функционального слоя эндометрия. В связи с регрессом желтого тела и резким снижением содержания половых стероидов в эндометрии нарастает гипоксия. Началу менструации способствует длительный спазм артерий, приводящий к стазу крови и образованию тромбов. Гипоксию тканей (тканевой ацидоз) усугубляют повышенная проницаемость эндотелия, ломкость стенок сосудов, многочисленные мелкие кровоизлияния и массивная лейкоцитарная инфильтрация. Выделяемые из лейкоцитов лизосомальные протеолитические ферменты усиливают расплавление тканевых элементов. Вслед за длительным спазмом сосудов наступает их паретическое расширение с усиленным притоком крови. При этом отмечаются рост гидростатического давления в микроциркуляторном русле и разрыв стенок сосудов, которые к этому времени в значительной степени утрачивают механическую прочность. На этом фоне и происходит активная десквамация некротизированных участков функционального слоя эндометрия. К концу 1-х суток менструации отторгается 2/3 функционального слоя, а его полная десквамация обычно заканчивается на 3-й день менструального цикла. Регенерация эндометрия начинается сразу после отторжения некротизированного функционального слоя. Основой для регенерации являются эпителиальные клетки стромы базального слоя. В физиологических условиях уже на 4-й день цикла вся раневая поверхность слизистой оболочки оказывается эпителизированной. Далее опять следуют циклические изменения эндометрия - фазы пролиферации и секреции. Последовательные изменения на протяжении цикла в эндометрии - пролиферация, секреция и менструация - зависят не только от циклических колебаний уровня половых стероидов в крови, но и от состояния тканевых рецепторов к этим гормонам. Концентрация ядерных рецепторов эстрадиола увеличивается до середины цикла, достигая пика к позднему периоду фазы пролиферации эндометрия. После овуляции наступает быстрое снижение концентрации ядерных рецепторов эстрадиола, продолжающееся до поздней секреторной фазы, когда их экспрессия становится значительно ниже, чем в начале цикла. Функциональное состояние маточных труб варьирует в зависимости от фазы менструального цикла. Так, в лютеиновую фазу цикла активируются реснитчатый аппарат мерцательного эпителия и сократительная активность мышечного слоя, направленные на оптимальный транспорт половых гамет в полость матки. Изменения в экстрагенитальных органах-мишенях Все половые гормоны не только определяют функциональные изменения в самой репродуктивной системе, но и активно влияют на обменные процессы в других органах и тканях, имеющих рецепторы к половым стероидам. В коже под влиянием эстрадиола и тестостерона активизируется синтез коллагена, что способствует поддержанию ее эластичности. Повышенная сальность, акне, фолликулиты, пористость кожи и избыточное оволосение возникают при повышении уровня андрогенов. В костях эстрогены, прогестерон и андрогены поддерживают нормальное ремоделирование, предупреждая костную резорбцию. Баланс половых стероидов влияет на метаболизм и распределение жировой ткани в женском организме. С воздействием половых гормонов на рецепторы в ЦНС и структурах гиппокампа связано изменение эмоциональной сферы и вегетативных реакций у женщины в дни, предшествующие менструации, - феномен "менструальной волны". Этот феномен проявляется разбалансированием процессов активации и торможения в коре головного мозга, колебаниями симпатической и парасимпатической нервной системы (особенно влияющей на сердечно-сосудистую систему). Внешними проявлениями данных колебаний служат изменения настроения и раздражительность. У здоровых женщин эти изменения не выходят за пределы физиологических границ. Влияние щитовидной железы и надпочечников на репродуктивную функцию Щитовидная железа вырабатывает два йодаминокислотных гормона - трийодтиронин (Т3) и тироксин (Т4), которые являются важнейшими регуляторами метаболизма, развития и дифференцировки всех тканей организма, особенно тироксин. Гормоны щитовидной железы оказывают определенное влияние на белково-синтетическую функцию печени, стимулируя образование глобулина, связывающего половые стероиды. Это отражается на балансе свободных (активных) и связанных стероидов яичника (эстрогенов, андрогенов). При недостатке Т3 и Т4 повышается секреция тиреолиберина, активизирующего не только тиреотрофы, но и лактотрофы гипофиза, что нередко становится причиной гиперпролактинемии. Параллельно снижается секреция ЛГ и ФСГ с торможением фолликуло- и стероидогенеза в яичниках. Возрастание уровня Т3 и Т4 сопровождается значительным увеличением концентрации глобулина, связывающего половые гормоны в печени и приводящего к уменьшению свободной фракции эстрогенов. Гипоэстрогения, в свою очередь, приводит к нарушению созревания фолликулов. Надпочечники. В норме продукция андрогенов - андростендиона и тестостерона - в надпочечниках оказывается такой же, как и в яичниках. В надпочечниках происходит образование ДГЭА и ДГЭА-С, тогда как в яичниках эти андрогены практически не синтезируются. ДГЭА-С, секретируемый в наибольшем (по сравнению с другими надпочечниковыми андрогенами) количестве, обладает относительно невысокой андрогенной активностью и служит своего рода резервной формой андрогенов. Надпочечниковые андрогены наряду с андрогенами яичникового происхождения являются субстратом для внегонадной продукции эстрогенов. Оценка состояния репродуктивной системы по данным тестов функциональной диагностики 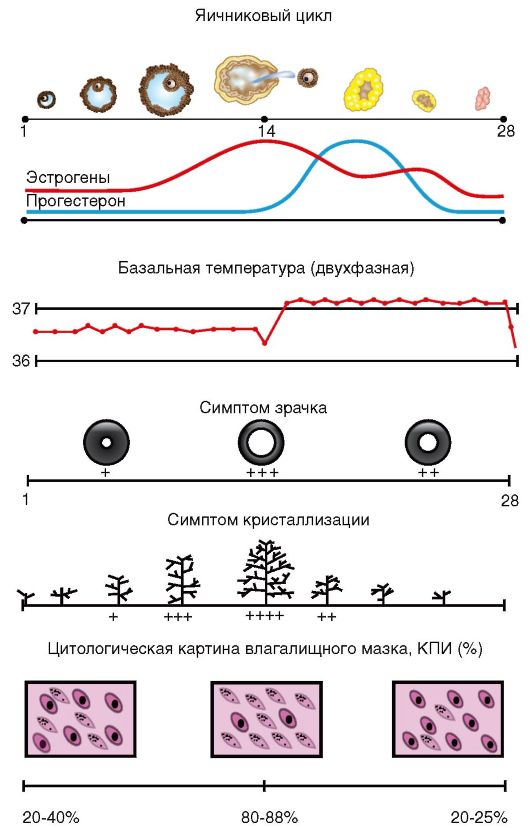 Рис. 4. Тесты функциональной диагностики при двухфазном менструальном цикле Тест базальной температуры основан на способности прогестерона (в возросшей концентрации) непосредственно воздействовать на центр терморегуляции в гипоталамусе. Под влиянием прогестерона во 2-ю (лютеиновую) фазу менструального цикла происходит транзиторная гипертермическая реакция. Пациентка ежедневно измеряет температуру в прямой кишке утром, не вставая с постели. Результаты отображают графически. При нормальном двухфазном менструальном цикле базальная температура в 1-ю (фолликулярную) фазу менструального цикла не превышает 37 °C, во 2-ю (лютеиновую) фазу отмечается повышение ректальной температуры на 0,4-0,8 °C по сравнению с исходным значением. В день менструации или за 1 сут до ее начала желтое тело в яичнике регрессирует, уровень прогестерона уменьшается, в связи с чем базальная температура снижается до исходных значений. Стойкий двухфазный цикл (базальную температуру нужно измерять на протяжении 2-3 менструальных циклов) свидетельствует о произошедшей овуляции и функциональной полноценности желтого тела. Отсутствие подъема температуры во 2-ю фазу цикла указывает на отсутствие овуляции (ановуляцию); запаздывание подъема, его кратковременность (повышение температуры на 2-7 дней) или недостаточный подъем (на 0,2-0,3 °C) - на неполноценную функцию желтого тела, т.е. недостаточность выработки прогестерона. Ложноположительный результат (повышение базальной температуры при отсутствии желтого тела) возможен при острых и хронических инфекциях, при некоторых изменениях ЦНС, сопровождающихся повышенной возбудимостью. Симптом "зрачка" отражает количество и состояние слизистого секрета в канале шейки матки, которые зависят от эстрогенной насыщенности организма. Феномен "зрачка" основан на расширении наружного зева цервикального канала из-за накопления в нем прозрачной стекловидной слизи и оценивается при осмотре шейки матки с помощью влагалищных зеркал. В зависимости от выраженности симптом "зрачка" оценивается по трем степеням: +, ++, +++. Синтез цервикальной слизи в течение 1-й фазы менструального цикла увеличивается и становится максимальным непосредственно перед овуляцией, что связано с прогрессирующим увеличением уровня эстрогенов в данный период. В преовуляторные дни расширенное наружное отверстие канала шейки матки напоминает зрачок (+++). Во 2-ю фазу менструального цикла количество эстрогенов уменьшается, в яичниках преимущественно вырабатывается прогестерон, поэтому количество слизи уменьшается (+), а перед менструацией и вовсе отсутствует (-). Тест нельзя использовать при патологических изменениях шейки матки. Симптом кристаллизации цервикальной слизи (феномен "папоротника") При высушивании максимально выражен во время овуляции, затем кристаллизация постепенно уменьшается, а перед менструацией вообще отсутствует. Кристаллизация слизи, высушенной на воздухе, оценивается также в баллах (от 1 до 3). Симптом натяжения шеечной слизи прямо пропорционален уровню эстрогенов в женском организме. Для проведения пробы корнцангом извлекают слизь из шеечного канала, бранши инструмента медленно раздвигают, определяя степень натяжения (расстояния, на котором слизь "разрывается"). Максимальное растяжение цервикальной слизи (до 10-12 см) происходит в период наибольшей концентрации эстрогенов - в середине менструального цикла, что соответствует овуляции. На слизь могут негативно влиять воспалительные процессы в половых органах, а также нарушения гормонального баланса. Кариопикнотический индекс (КПИ). Под влиянием эстрогенов происходит пролиферация клеток базального слоя многослойного плоского эпителия влагалища, в связи с чем в поверхностном слое увеличивается количество ороговевающих (отшелушивающихся, отмирающих) клеток. Первой стадией гибели клеток являются изменения их ядра (кариопикноз). КПИ - это отношение числа клеток с пикнотическим ядром (т.е. ороговевающих) к общему числу эпителиальных клеток в мазке, выраженное в процентах. В начале фолликулярной фазы менструального цикла КПИ составляет 20- 40%, в преовуляторные дни повышается до 80-88%, что связано с прогрессирующим увеличением уровня эстрогенов. В лютеиновой фазе цикла уровень эстрогенов снижается, следовательно, КПИ уменьшается до 20- 25%. Таким образом, количественные соотношения клеточных элементов в мазках слизистой оболочки влагалища позволяют судить о насыщенности организма эстрогенами. В настоящее время, особенно в программе экстракорпорального оплодотворения (ЭКО), созревание фолликула, овуляция и образование желтого тела определяются при динамическом УЗИ. |