Липиды. Липиды, их строение, свойства и функции
 Скачать 1.31 Mb. Скачать 1.31 Mb.
|

|
Липиды, их строение, свойства и функции Липиды — это сложные эфиры трёхатомного спирта глицерина и высших жирных кислот. В каждом из них есть кислотный остаток СООН, он, теряя атом водорода, соединяется с глицерином, а с остатком соединяется углеродная цепочка. Липиды — низкомолекулярные гидрофобные органические соединения. 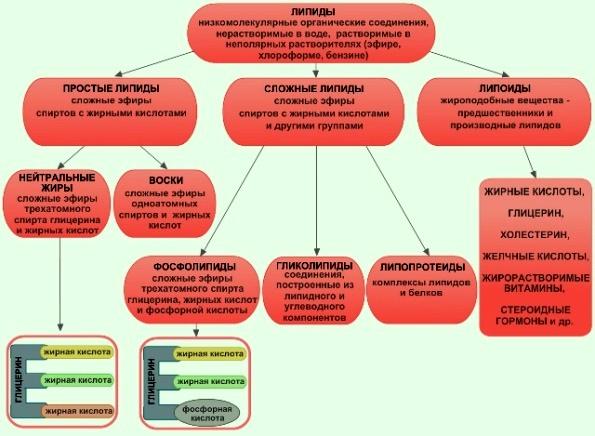 К липидам относятся жиры, воски, стероиды, фосфолипиды, терпены, гликолипиды, липопротеиды. Нейтральные жиры (триглицериды) представляют собой соединения высокомолекулярных жирных кислот и трехатомного спирта глицерина. В цитоплазме клеток триглицериды откладываются в виде жировых капель. Воски — пластичные вещества, обладающие водоотталкивающими свойствами. У насекомых они служат материалом для постройки сот. Восковой налет на поверхности листьев, стеблей, плодов защищает растения от механических повреждений, ультрафиолетового излучения и играет важную роль в регуляции водного баланса. Фосфолипиды — представители класса жироподобных веществ, являющиеся сложными эфирами глицерина и жирных кислот, содержащие остаток фосфорной кислоты. Они формируют основу всех биологических мембран. По своей структуре фосфолипиды сходны с жирами, но в их молекуле один или два остатка жирных кислот замещены остатком фосфорной кислоты. Гликолипиды — вещества, образующиеся в результате соединения углеводов и липидов. Углеводные компоненты гликолипидных молекул полярны, и это определяет их роль: подобно фосфолипидам гликолипиды входят в состав клеточных мембран. Функции липидов: 1. Энергетическая функция — одна из важнейших функций жиров. При окислении жиров выделяется в два раза больше энергии, чем при окислении такой же массы углеводов и белков. 2. Фосфолипиды выполняют структурную функцию — они образуют все плазматические мембраны в клетке. 3. Запасающую функцию в живых организмах выполняют жиры. Они откладываются про запас в семенах и плодах растений, в жировой клетчатке животных. 3. Жиры могут служить источником воды, так как при окислении 1 г жира образуется более 1 г воды. Поэтому некоторые животные могут долгое время выдерживать без воды (верблюды в пустыне — до двух недель, а медведи зимой — более двух месяцев). 4. Защитная функция проявляется в том, что запасы жира защищают внутренние органы от травм. 5. Подкожный жир сохраняет тепло и выполняет теплоизоляционную функцию. 6. Миелин, покрывающий отростки нервных клеток, изолирует их, ускоряя передачу нервных импульсов (электроизоляционная функция). 7. Некоторые гормоны (кортизон, альдостерон, тестостерон, прогестерон) имеют стероидную природу и выполняют регуляторную функцию. 8. Воски выполняют смазывающую функцию. Они покрывают листья и плоды многих растений, кожу, шерсть, перья животных и защищают их от намокания. Пчёлы используют воск как строительный материал для сот. Основу биологических мембран составляют фосфолипиды. Это амфифильные молекулы, то есть молекулы, имеющие гидрофобную и гидрофильную части. Гидрофильная часть взаимодействует с водой, а гидрофобные «прячутся» от воды. В результате могут образовываться разные типы структур — мицеллы (их образуют молекулы мыла), липосомы и бислои — их образуют фосфолипиды. В состав липидов мембран входят в основном фосфолипиды, сфингомиелины и холестерин.  Таким образом, в составе молекул всех фосфолипидов имеются гидрофобная (остатки жирных кислот) и гидрофильная (фосфорная кислота и присоединенные к ней группы) части, поэтому такие молекулы могут контактировать как с полярными, так и с неполярными растворителями (такие вещества называют амфифильными). Поскольку у большинства молекул фосфолипидов имеется гидрофильная голова и два гидрофобных хвоста, то их часто схематично представляют в виде кружка (гидрофильная часть) с двумя хвостами (гидрофобная часть). Гидрофобные слои смотрят друг на друга, а гидрофильные головы – в водную среду. Такие слои являются основой всех биологических мембран. Таким образом, одной из основных функцией фосфолипидов является структурная функция — формирование биологических мембран. 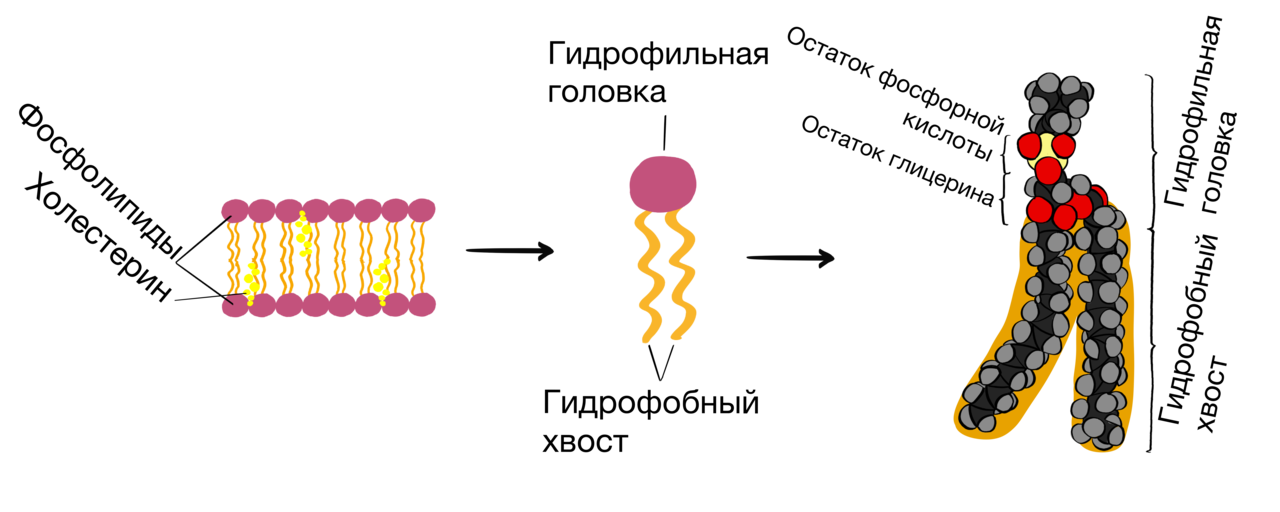 Стероиды Особое место среди липидов занимают стероиды — полициклический спирт холестерол (чаще называемый холестерин) и его производные. Холестерин и его эфиры с жирными кислотами входят в состав биологических мембран клеток животных, придавая им определенную «жесткость» (структурная функция). Белки в липидном слое Если основная роль липидов в составе мембран заключается в стабилизации бислоя, то белки отвечают за функциональную активность мембран. Одни из них обеспечивают транспорт определённых молекул и ионов, другие являются ферментами, третьи участвуют в связывании цитоскелета с внеклеточным матриксом или служат рецепторами для гормонов, медиаторов, эйкозаноидов, липопротеинов, оксида азота (NО). Белки определяют особенности функционирования каждой мембраны.
Мембранные белки, контактирующие с гидрофобной частью липидного бислоя, должны быть амфифильными. Те участки белка, которые взаимодействуют с углеводородными цепями жирных кислот, содержат преимущественно 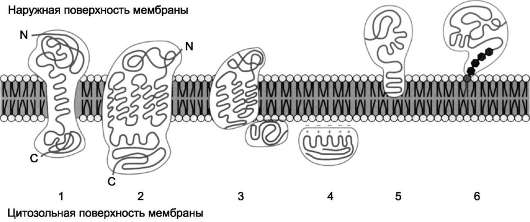 Рис. 5-10. Расположение (локализация) белков в мембранах. Трансмембранные белки, например: 1 - гли-кофорин А; 2 - рецептор адреналина. Поверхностные белки: 3 - белки, связанные с интегральными белками, например, фермент сукцинатдегидрогеназа; 4 - белки, присоединённые к полярным «головкам» липидного слоя, например, протеинкиназа С; 5 - белки, «заякоренные» в мембране с помощью короткого гидрофобного концевого домена, например, цитохром b5; 6 - «заякоренные» белки, ковалентно соединённые с липидом мембраны (например, фермент щелочная фосфатаза). Участки белка, находящиеся в области полярных «головок», обогащены гидрофильными аминокислотными остатками. Белки мембран различаются по своему положению в мембране. Они могут глубоко проникать в липидный бислой или даже пронизывать его - интегральные белки, либо разными способами прикрепляться к мембране - поверхностные белки. Поверхностные белки Поверхностные белки часто прикрепляются к мембране, взаимодействуя с интегральными белками или поверхностными участками ли-пидного слоя. Белки, образующие комплексы с интегральными белками мембраны Ряд пищеварительных ферментов, участвующих в гидролизе крахмала и белков, прикрепляется к интегральным белкам мембран микроворсинок кишечника. Примерами таких комплексов могут быть сахараза-изомальтаза и мальтаза-гликоамилаза. Возможно, связь этих пищеварительных ферментов с мембраной позволяет с высокой скоростью гидролизовать субстраты и усваивать продукты гидролиза клеткой.
Белки, связанные с полярными «головками» липидов мембран Полярные или заряженные домены белковой молекулы могут взаимодействовать с полярными «головками» липидов, образуя ионные и водородные связи. Кроме того, множество растворимых в цитозоле белков при определённых условиях могут связываться с поверхностью мембраны на непродолжительное время. Иногда связывание белка - необходимое условие проявления ферментативной активности. К таким белкам, например, относят протеинкиназу С, факторы свёртывания крови. Закрепление с помощью мембранного «якоря» «Якорем» может быть неполярный домен белка, построенный из аминокислот с гидрофобными радикалами. Примером такого белка может служить цитохром b5 мембраны ЭР. Этот белок участвует в окислительно-восстановительных реакциях, как переносчик электронов. Роль мембранного «якоря» может выполнять также ковалентно связанный с белком остаток жирной кислоты (миристиновой - С14 или пальмитиновой - С16). Белки, связанные с жирными кислотами, локализованы в основном на внутренней поверхности плазматической мембраны. Миристиновая кислота присоединяется к N-концевому глицину с образованием амидной связи. Пальмитиновая кислота образует тиоэ-фирную связь с цистеином или сложноэфирную с остатками серина и треонина. Небольшая группа белков может взаимодействовать с наружной поверхностью клетки с помощью ковалентно присоединённого к С-концу белка фосфатидилинозитолгликана. Этот «якорь» - часто единственное связующее звено между белком и мембраной, поэтому при действии фосфолипазы С этот белок отделяется от мембраны. Трансмембранные (интегральные) белки Некоторые из трансмембранных белков пронизывают мембрану один раз (гликофо-рин), другие имеют несколько участков (доменов), последовательно пересекающих бислой.
Трансмембранные домены, пронизывающие бислой, имеют конформацию α-спирали. Полярные остатки аминокислот обращены внутрь глобулы, а неполярные контактируют с мембранными липидами. Такие белки называют «вывернутыми» по сравнению с растворимыми в воде белками, в которых большинство гидрофобных остатков аминокислот спрятано внутрь, а гидрофильные располагаются на поверхности. Радикалы заряженных аминокислот в составе этих доменов лишены заряда и протонированы (-СООН) или депротонированы (-NH2). Гликозилированные белки Поверхностные белки или домены интегральных белков, расположенные на наружной поверхности всех мембран, почти всегда гликозилированы. Олигосахаридные остатки могут быть присоединены через амидную группу аспарагина или гидроксильные группы серина и треонина. Олигосахаридные остатки защищают белок от протеолиза, участвуют в узнавании лигандов или адгезии. Латеральная диффузия белков Некоторые мембранные белки перемещаются вдоль бислоя (латеральная диффузия) или поворачиваются вокруг оси, перпендикулярно его поверхности. Например, фермент фосфолипаза А2, связываясь с цитоплазматической поверхностью мембраны, может латерально перемещаться по поверхности бислоя и гидролизовать несколько тысяч фосфолипидов в минуту до тех пор, пока не отделится от мембраны. Латеральная диффузия интегральных белков в мембране ограничена, это связано с их большими размерами, взаимодействием с другими мембранными белками, элементами цитоскелета или внеклеточного матрикса. Белки мембран не совершают перемещений с одной стороны мембраны на другую («флип-флоп» перескоки), подобно фосфолипидам. Транспорт веществ через мембрану Классификация видов транспорта: Относительно затрат энергии в процессе переноса веществ транспорт подразделяется на активный и пассивный. Активный транспорт осуществляется с затратой энергии (за счёт гидролиза АТФ) и против градиента концентрации (иногда данный вид транспорта называют первично-активным). Если же перенос веществ обеспечивает энергия, обусловленная градиентом других ионов, такой транспорт называют сопряженным (иногда используется термин вторично-активный транспорт). Пассивный транспорт осуществляется без затраты энергии, протекает самопроизвольно и по градиенту концентрации. (Градиент в процессе пассивного транспорта снижается). Относительно сопряженности переноса веществ через мембрану выделяют: Унипорт – перенос данного вещества (иона) не зависит от наличия и переноса других веществ и котранспорт – перенос данного вещества (иона) зависит от наличия и переноса других веществ. В этом случае скорость суммарного процесса контролируется наличием и доступностью для систем переноса обоих партнеров транспортного процесса. Котранспорт относительно направления подразделяют на симпорт и антипорт. При симпорте осуществляется однонаправленный и одновременный транспорт, а при антипорте – одновременный, но разнонаправленный перенос веществ. Специфические механизмы переноса веществ, связанные с нарушением структурной целостности мембраны: эндоцитоз и экзоцитоз – важнейшие процессы жизнедеятельности клетки, механизмы высвобождения медиатора в синаптическую щель, фагоцитоз лейкоцитов и др. Пассивный транспорт Пассивный транспорт может происходить непосредственно через фосфолипидный слой, через белки- переносчики или через белковые каналы. Движущая сила может обеспечиваться: разностью концентрации транспортируемого вещества (диффузия) или осмотического давления (осмос) на разных сторонах мембраны. транспорт воды обеспечивается разностью осмотического давления с помощью белков-аквапоринов. разностью электрического потенциала на мембране (если транспортируемое вещество несет заряд); Простая диффузия — перенос веществ через мембрану по градиенту концентрации (из области высокой концентрации в область низкой концентрации) без затрат энергии. происходит по электрохимическому градиенту; скорость линейно зависит от градиента концентрации вещества; не насыщаемый процесс, то есть может ускоряться неограниченно; не расходуется энергия. Путём простой диффузии в клетку проникают гидрофобные вещества (кислород, азот, бензол) и полярные маленькие молекулы (вода, углекислый газ, мочевина). Не проникают полярные относительно крупные молекулы (аминокислоты, моносахариды), заряженные частицы (ионы) и макромолекулы (ДНК, белки). Облегченная диффузия — процесс трансмембранного переноса веществ по градиенту концентрации с участием мембранных белков-переносчиков без затраты энергии. Каналы — белковые молекулы в мембране, создающие гидрофильный проход (облегченная диффузия). Характеристика каналов: селективны (специфичны); изменяют свое состояние; чувствительны к лигандам и заряду мембраны. С помощью облегченной диффузии в клетку попадают органические вещества с высокой полярностью. 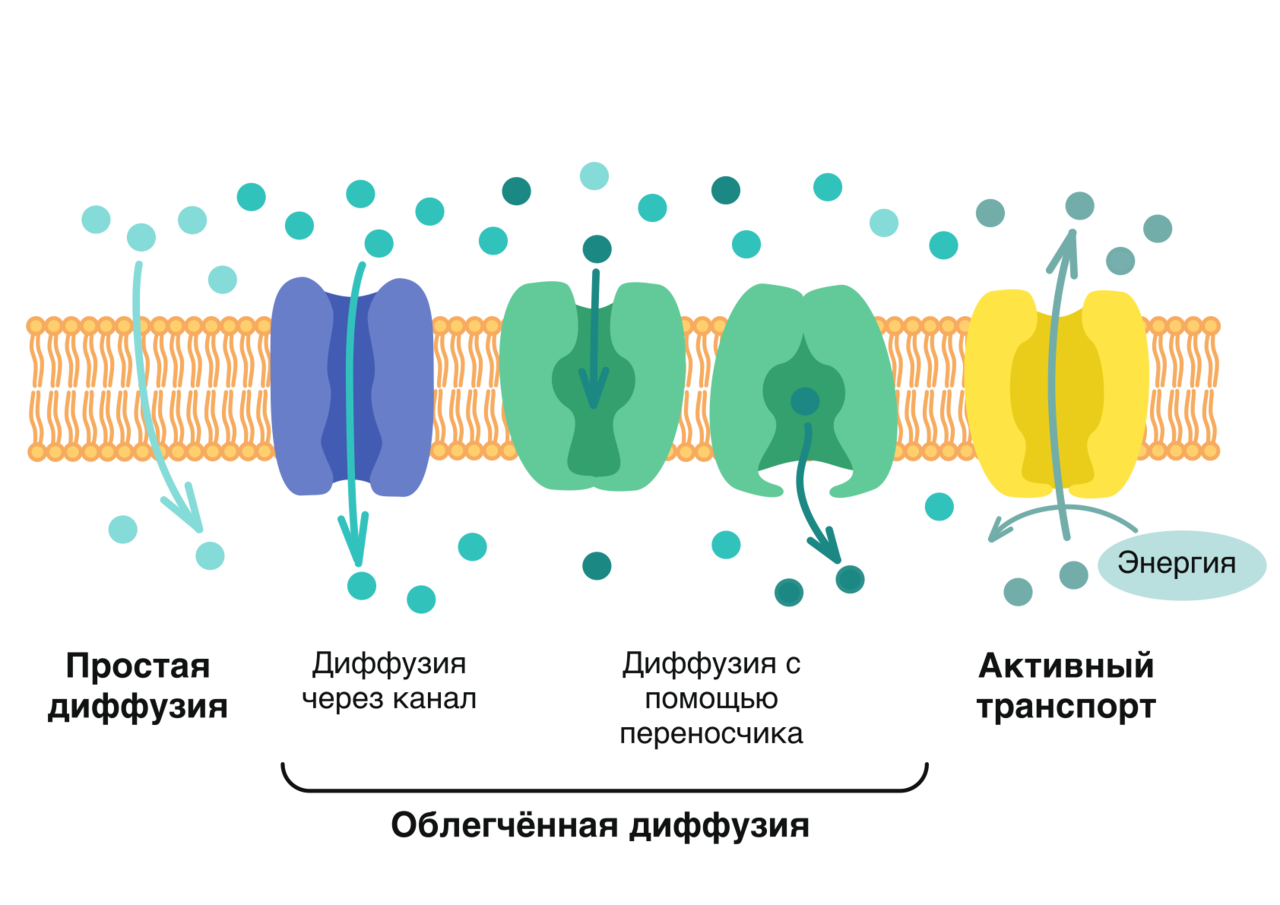 Особенности облегченной диффузии быстрее, чем простая диффузия; происходит по электрохимическому градиенту; с помощью белка-переносчика или белков трансмембранных каналов; ограниченный по скорости и насыщаемый процесс; можно блокировать, связав переносчик — чувствительна к ингибиторам; конкуренция переносимых веществ за переносчик; не расходуется энергия. Активный транспорт Активный транспорт — процесс трансмембранного переноса веществ против их градиента концентрации с затратами энергии. Активный транспорт всегда происходит посредством белков-носителей, называемых транспортерами. Деятельность белкового насоса: зависит от источника метаболической энергии: первичные транспортеры требуют прямого использования АТФ, например, транспортные механизмы для Na, K, Са; вторичные транспортеры — белки, чьи транспортные функции требуют одновременного перемещения иона (обычно Na) по градиенту концентрации, который поддерживается первичными транспортерами (например, транспортные механизмы для глюкозы и аминокислот); 2. вещества транспортируются против их электрохимического градиента 3. происходит только в одном направлении через плазматическую мембрану. Унипорт — перенос через мембрану отдельного растворенного вещества. Более сложная функция переносчиков-транспортировать два растворенных вещества совместно, так что перенос одного зависит от одновременного переноса другого в том же самом направлении (симпорт) или в противоположном направлении (антипорт). Энергия для активного транспорта выделяется при гидролизе АТФ: АТФ = АДФ + Ф + энергия С помощью активного транспорта в клетку проникают ионы (К, Na , Са, Mg) и мономеры (простые сахара, аминокислоты, нуклеотиды). Осуществляется специальными белками-переносчиками, образующими так называемые ионные насосы, или помпы. Наиболее изученным является натрий-калиевый насос в клетках животных, активно выкачивающих ионы Na наружу, поглощая при этом ионы К. Благодаря этому в клетке поддерживается большая концентрация К и меньшая Na по сравнению с окружающей средой. На этот процесс затрачивается энергия АТФ. В результате активного транспорта с помощью мембранного насоса в клетке происходит также регуляция концентрации Mg и Са. Ионный канал образуется одной или несколькими белковыми субъединицами, окружающими центральную пору. Транспорт через ионные каналы всегда пассивен, и его максимальная скорость приблизительно в 1 000 раз больше, чем у белков-переносчиков. Ионные каналы высоко селективны для определенных ионов Ионные каналы изменяют свою активность (открываются в ответ на определенный стимул (раздражитель), который может быть электрическим, химическим или механическим). Электрочувствительные каналы, осуществляя перенос Na, К или Са, имеют большое значение для возбудимых клеток (например, нервных и мышечных). Лигандчувствительные каналы распространены во всех органах и тканях и участвуют в восприятии клетками химических раздражителей. Натрий, каливый насос Натрий-калиевый насос транспортирует натрий из клетки, и калий в клетку в повторяющемся цикле конформационных изменений (формы). В каждом цикле, три иона натрия покидают клетку, в то время как два иона калия проникают в неё. Этот процесс выполняется пошагово: 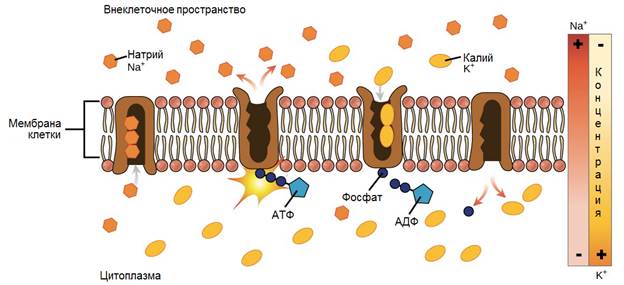 1. Сначала насос-переносчик открыт для внутренней стороны клетки. В этом расположении насос-переносчик легко соединяется с Na+ ионами и связывает три из них. 2. Когда Na+ ионы связываются, они запускают насос-переносчик для гидролиза (разложения) АТФ. Одна фосфатная группа из АТФ прикрепляется к насосу-переносчику, который затем называется фосфорилированным. АДФ выпускается как побочный продукт. 3. Фосфорилирование приводит к изменению положения насоса-переносчика, его переориентации, от чего он открывается в сторону внеклеточного пространства. В этой конформации, насос-переносчик больше не связывается с Na+ ионами, поэтому три Na+ иона высвобождаются наружу клетки. 4. В этом направленном наружу расположении, насос-переносчик меняет конформацию и теперь легко связывается с K+ ионам. Он связывае два из них, и это вызовает удаление фосфатной группы, прикрепленной к насосу. 5. С исчезновением фосфатной группы, насос-переносчик снова возращается к исходному положению, направив себя внутрь клетки. 6. В своем обращенном внутрь положении, насос-переносчик больше не связывается с K+ ионами, поэтому два K+ иона будут высвобождаться в цитоплазму. Затем насос возвращается в то расположение, в котором он находился на этапе 1, и цикл может начаться снова. Эндоцитоз Транспорт макромолекул, их комплексов и крупных частиц внутрь клетки происходит совершенно иным путем — посредством эндоцитоза. Выведение тех же грузов из клетки называется экзоцитозом. При эндоцитозе (эндо — внутрь) определенный участок плазмалеммы захватывает и как бы обволакивает внеклеточный материал, заключая его в мембранную вакуоль, возникшую вследствие впячивания мембраны. В дальнейшем такая вакуоль соединяется с лизосомой, ферменты которой расщепляют макромолекулы до мономеров. 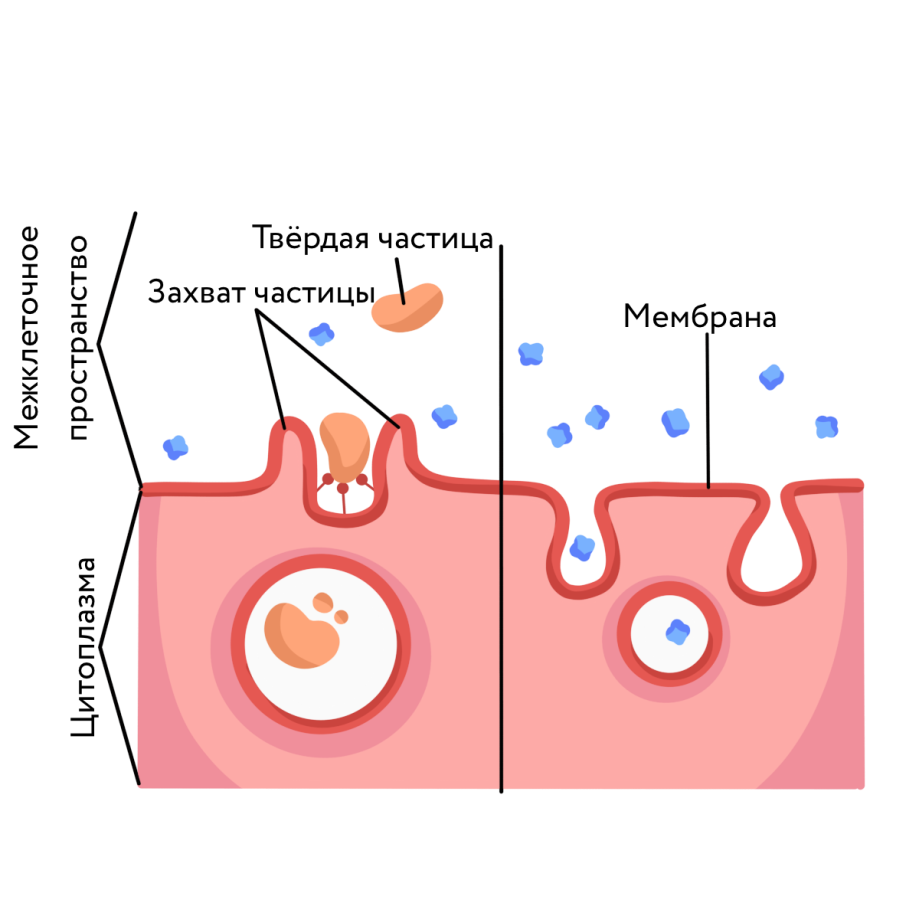 Опосредуемый рецепторами эндоцитоз представляет собой избирательный концентрирующий механизм, позволяющий клеткам захватывать большие количества специфических лигандов без поглощения соответственно большего объема внеклеточной жидкости. При этом поглощаются макромолекулы, для которых на плазмалемме существует ограниченное число связывающих участков. Эти рецепторы, обладают высоким сродством к определенным веществам. Они выборочно поглощают их из среды клетки и концентрируют. При этом жидкости и растворенные в ней посторонние, несвязывающиеся с рецепторами молекулы, почти не поглощаются. Так обеспечивается эффективное поступление специфических молекул в клетку. Везикулы, образующиеся при таком эндоцитозе, формируются в месте инвагинаций плазмалеммы, покрытых с цитоплазматической стороны волокнистым материалом — мембранным белком клатрином. Предварительно на мембране образуются окаймленные клатрином ямки, которые могут занимать до 2% поверхности некоторых клеток. С помощью окаймленных ямок, в которых располагаются соответствующие рецепторы, интернализуются ряд веществ, например, липопротеины низкой плотности, факторы роста, гормоны и пр. 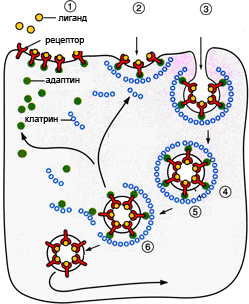 Рисунок 1. Этапы эндоцитоза, опосредованного рецепторами. 1. Связывание лиганда с рецептором. 2. Формирование окаймленной ямки. 3. Образование клатриновой везикулы. 4–6. Внутриклеточная утилизация. Экзоцитоз Экзоцитоз (экзо — наружу) — процесс, обратный эндоцитозу. Благодаря ему клетка выводит внутриклеточные продукты или непереваренные остатки, заключенные в вакуоли или пузырьки. Пузырек подходит к цитоплазматической мембране, сливается с ней, а его содержимое выделяется в окружающую среду. Так выводятся пищеварительные ферменты, гормоны, гемицеллюлоза и др. 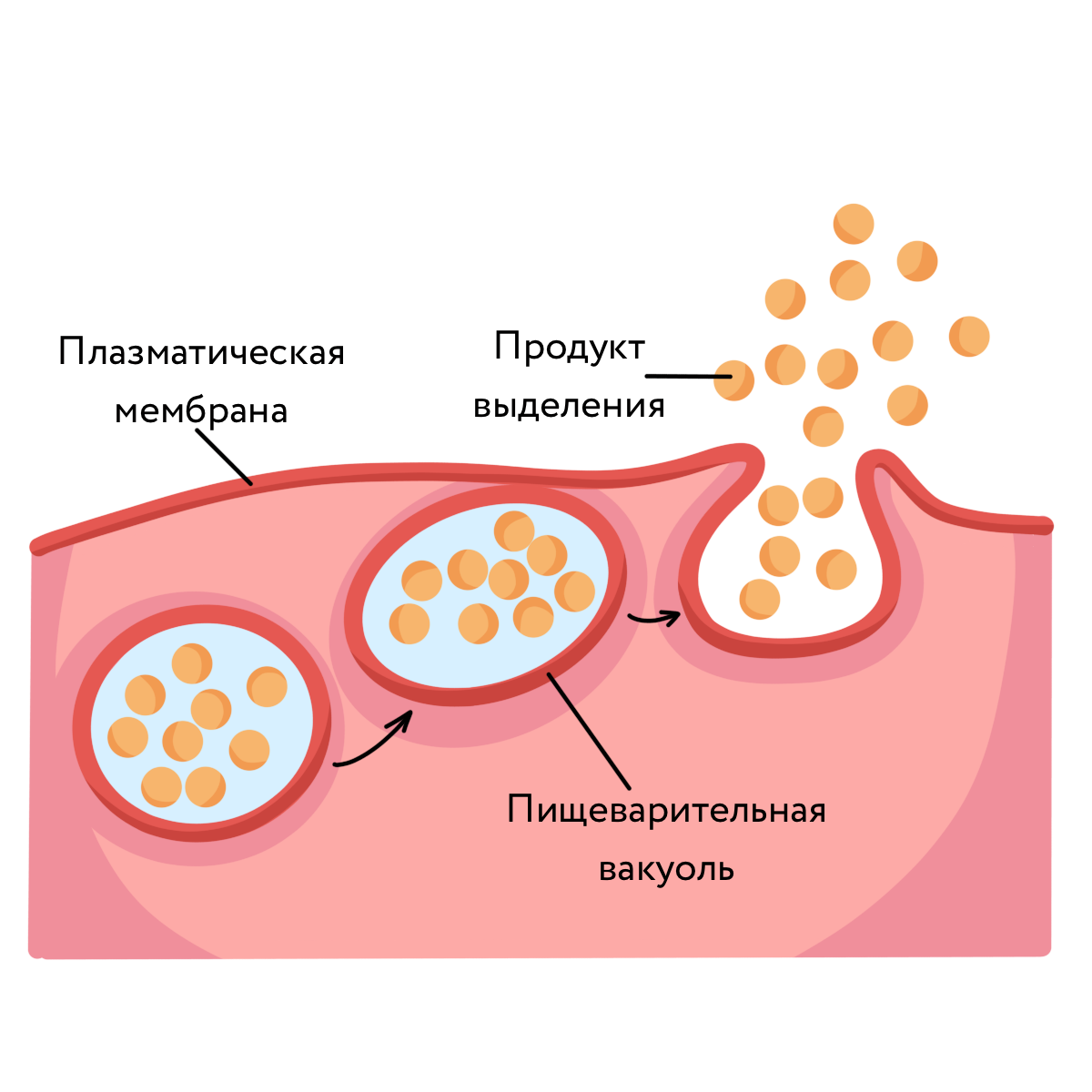 Таким образом, биологические мембраны как основные структурные элементы клетки служат не просто физическими границами, а представляют собой динамичные функциональные поверхности. На мембранах органелл осуществляются многочисленные биохимические процессы, такие как активное поглощение веществ, преобразование энергии, синтез АТФ и др. Фагоцитоз Осуществляется фагоцитами (циркулирующими в крови моноцитами и нейтрофилами и в тканях – макрофагами). Поглощают субстрат (твердые частицы), связываются с рецепторами цитоплазмы мембраны специальных клеток, затем происходит активирование мембраны и ее втягивание внутрь клетки при помощи актина. Фагоцитарный процесс интенсифицирует при опсонизации фагоцитарных частиц, т.е. фиксации на поверхности объектов опсонинов, к которым у фагоцитов есть специфичные рецепторы (Fc – рецепторы) – фрагменты антител, компоненты системы комплемента, фибронектин. |