метаболизм липидов. МЕТАБОЛІЗМ ЛІПІДІВ. Метаболізм ліпідів 14 Перетворення ліпідів у шлунковокишковому тракті
 Скачать 172.73 Kb. Скачать 172.73 Kb.
|

7 %о) і білками (2 %), які утворюють гідрофільну оболонку навколо гідрофобного ядра.
0,5 г). Розщеплення ліпідів їжі здійснюється шляхом гідролізу за участю ліполітичних ферментів шлунково-кишкового тракту: ліпази триацилгліцеролів, фосфоліпаз, естераз холестеридів. 50-60 г), а також фосфоліпідами ( |
У ротовій порожнині ліпіди через відсутність у складі слини ліполітичних ферментів не зазнають перетворень, їхнє травлення починається у шлунку, де є ліпаза триацигліцеролів. Але активна вона тільки в новонароджених ссавців і в дітей грудного віку, у шлунку яких величина рН середовища (5-5,5) відповідає рН-оптимуму цього ензиму. У кислому середовищі шлунка дорослої людини ліпаза неактивна й тому гідролітичне розщеплення жирів та інших ліпідів практично здійснюється в тонкому кишечнику.
Верхній відділ тонкого кишечнику - дванадцятипала кишка в комплексі з підшлунковою залозою і печінкою - є "центром" травлення всіх складових компонентів їжі, у тому числі й ліпідів.
Поява хімусу й пов'язана з цим ацидифікація у дванадцятипалій кишці викликає секрецію гормонів секретину й холецистокініну клітинами слизової оболонки тонкого кишечнику. Секретин стимулює секрецію панкреатичного соку з високою концентрацією бікарбонатів (у людини до 150 ммоль/л) і незначним вмістом ферментів. У результаті нейтралізації бікарбонатами кислого середовища утворюється оптимальні для гідролізу жирів умови:
У панкреатичному секреті, що продовжує при цьому надходити у дванадцятипалу кишку, змінюється співвідношення неорганічних і органічних складових з переважанням останніх, зокрема білків-ферментів. Неактивні ліполітичні ферменти активуються в просвіті дванадцятипалої кишки. Активатором ліпази триацилгліцеролів є білок коліпаза, який секретується разом із ліпазою. Необхідними умовами для каталітичної активності ліпази, крім наявності коліпази й нейтральної реакції середовища, є також присутність емульгаторів. Оскільки реакція гідролізу ліпідів може відбуватися тільки на межі розподілу між водною та ліпідною фазами, ефективність каталізу залежить від величини доступної поверхні розподілу, тобто від ступеня емульгування жиру. Чим дрібніші краплини жиру, тим доступніша поверхня для гідролізу. Тому, зокрема, ідеальним субстратом для ліпази є молоко - тонка емульсія жиру у воді. Основними природними емульгаторами жирів є жовчні кислоти.
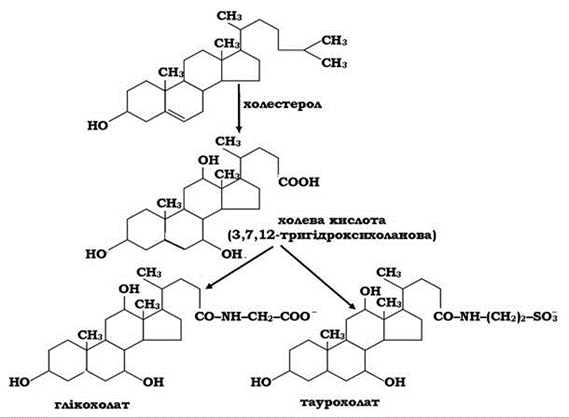
Жовчні кислоти за своєю природою належать до стероїдів, похідних холестеролу. Із більш ніж 30 відомих жовчних кислот найважливішими є холева й хенодезоксихолева кислоти та їхні похідні - дезоксихолева й літохолева. Синтезуються жовчні кислоти в печінці у вигляді кон'югатів із гліцином і таурином (рис. 14.1). Кон'юговані з гліцином жовчні кислоти - глікохолати - характер - ні для ссавців. У людини вміст глікохолатів становить близько 75 %, частка таурохолатів - 25 %, проте у приматів та інших тварин, навпаки, переважають таурохолати.
Кон'югати жовчних кислот накопичуються в жовчному міхурі, і за концентрації приблизно 2 ммоль/л у складі жовчі разом із
 фосфоліпідами та холестеролом утворюють міцели, утримуючи таким чином холестерол у розчинному стані. При порушенні умов міцелоутворення (підвищенні вмісту холестеролу або зниженні концентрації жовчних кислот чи фосфоліпідів) холестерол може осаджуватися в жовчних протоках і утворювати жовчні конкременти.
фосфоліпідами та холестеролом утворюють міцели, утримуючи таким чином холестерол у розчинному стані. При порушенні умов міцелоутворення (підвищенні вмісту холестеролу або зниженні концентрації жовчних кислот чи фосфоліпідів) холестерол може осаджуватися в жовчних протоках і утворювати жовчні конкременти. У складі жовчі кон'югати жовчних кислот легко іонізуються, утворюючи солі переважно з Na+. Завдяки амфіпа- тичним властивостям солей жовчних кислот та їхніх кон'югатів, зумовлених гідрофільністю залишків гліцину й таурину і гідрофобністю стероїдного ядра, кон'югати жовчних кислот та їхні солі є ефективними природними детергентами. Емульгуванню жирів їжі сприяють також фосфоліпіди, моноацилгліцероли та перистальтика кишечнику. За дії емульгаторів поверхневий натяг жирових крапель зменшується й вони розпадаються на дрібніші структури - міцели.
Рис. 14.1. Схема синтезу глікохолату и таурохолату
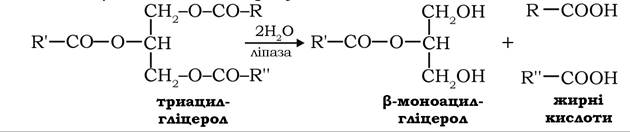
Гідроліз жирів здійснюється на поверхні цих структур за участю ліпази, яка швидко розщеплює ефірні зв'язки в молекулах триацилгліцеролів у α- та γ-положенні з вивільненням жирних кислот і β-моноацилгліцеролу:
Гідроліз ліпідів їжі у дванадцятипалій кишці відбувається під гормональним контролем, зокрема з боку холецистокініну, який вивільняється при появі в кишечнику продуктів травлення ліпідів. Він стимулює секрецію жовчі й ферментів панкреатичної залози, підтримуючи тим самим процес травлення.
Гідроліз фосфоліпідів, що здійснюється одночасно з жирами, каталізується фосфоліпазами, які також секретуються підшлунковою залозою (рис. 14.2):
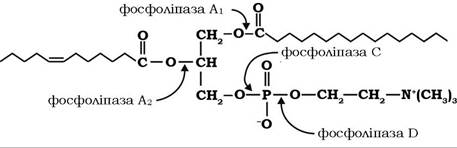
Рис. 14.2. Специфічність дії різних фосфоліпаз
Після активації трипсином фосфоліпази послідовно розщеплюють ефірні зв'язки молекули ліпіду: спочатку фосфоліпазою А2 гідролізується ефірний зв'язок біля вуглецю-2 молекули фосфоліпіду з утворенням лізофосфоліпіду, у подальшому, за участю фосфоліпази А1, С і D, відщеплюється жирна кислота, вивільняються холін, гліцерол і фосфорна кислота.
Холестериди (ефіри холестеролу), які потрапляють в організм з їжею, розщеплюються панкреатичною холестеролестеразою з вивільненням холестеролу й жирних кислот.
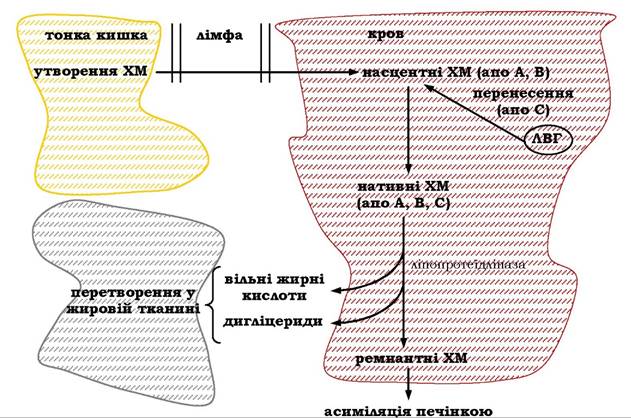
Усі продукти гідролізу ліпідів у тонкому кишечнику разом із жовчними кислотами всмоктуються клітинами слизової тонкої кишки. При цьому молекули гліцеролу, аміноспиртів, фосфорної кислоти, а також коротколанцюгові жирні кислоти вільно, шляхом пасивної дифузії потрапляють у ентероцити, далі в міжклітинний простір і з кров'ю портальної вени надходять у печінку.
Амфіфільні продукти розщеплення ліпідів - вищі жирні кислоти, моноацилгліцероли, жиророзчинні вітаміни разом із жовчними кислотами й холестеролом - утворюють змішані міцели і в такій формі транспортуються в клітини слизової оболонки, де міцели розпадаються на складові.
Вивільнені жовчні кислоти з кров'ю портальної вени потрапляють у печінку й повторно використовуються при гідролізі ліпідів. Ефективність такої реутилізації жовчних кислот, яка дістала назву ентерогепатичної циркуляції жовчних кислот, становить майже 90 %. Інші компоненти змішаних міцел - моноацилгліце- роли, вищі жирні кислоти, холестерол і фосфоліпіди - беруть участь в утворенні ліпід-білкових комплексів - хіломікронів. Ліпідні й білкові компоненти хіломікронів синтезуються на мембранах ендоплазматичного ретикулума клітин слизової оболонки тонкого кишечнику: на мембранах гладенького ретикулума ресинтезуються ліпіди, на рибосомах синтезуються специфічні білки хіломікронів - апопротеїни.
Ресинтез триацилгліцеролів здійснюється шляхом прямої етерифікації β-моноацилгліцеролів, КоА-ефірами жирних кислот. У реакціях так званого моноацилгліцеролового шунта жирні кислоти спочатку активуються синтетазою за участю КоА-SH і далі переносяться ацилтрансферазами на β-моноацилгліцерол:
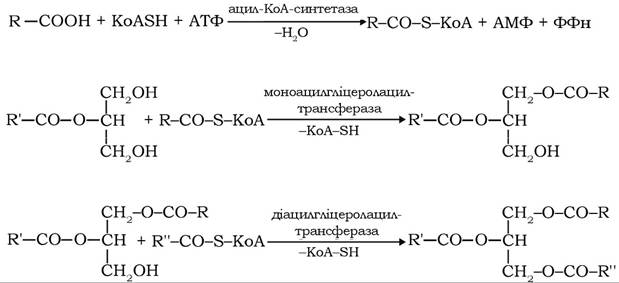
Таким самим шляхом ресинтезуються і деякі фосфоліпіди. Ресинтезовані ліпіди й синтезовані апопротеїни, зокрема апопротеїн В, шляхом ліпід-ліпідних, ліпід-білкових взаємодій утворюють хіломі- крони. Це невеликі сферичні утворення діаметром