МембраныМикротрубочкиМитохондрии. Надмембранные структуры поверхностного аппарата. Клеточные оболочки граммположительных и граммотрицательных бактерий. Гликокаликс
 Скачать 346 Kb. Скачать 346 Kb.
|

|
Надмембранные структуры поверхностного аппарата. Клеточные оболочки граммположительных и граммотрицательных бактерий. Гликокаликс Надмембранный комплекс поверхностного аппарата клеток характеризуется многообразием строения .У прокариот надмембранный комплекс представлен клеточной стенкой различной толщины, основу которой составляет сложный гликопротеин муреин (у архебактерий - псевдомуреин). У целого ряда эубактерий наружная часть надмембранного комплекса состоит из еще одной мембраны с большим содержанием липополисахаридов.У эукариот универсальным компонентом надмембранного комплекса являются углеводы - компоненты гликолипидов и гликопротеинов плазмалеммы. Благодаря этому его исходно называли гликокаликсом (от греч. гликос - сладкий, углевод и лат. каллум - толстая кожа, оболочка). Кроме углеводов, в состав гликокаликса относят периферические белки над билипидным слоем. Более сложные варианты надмембранного комплекса встречаются у растений (клеточная стенка из целлюлозы), грибов и членистоногих (наружный покров из хитина). 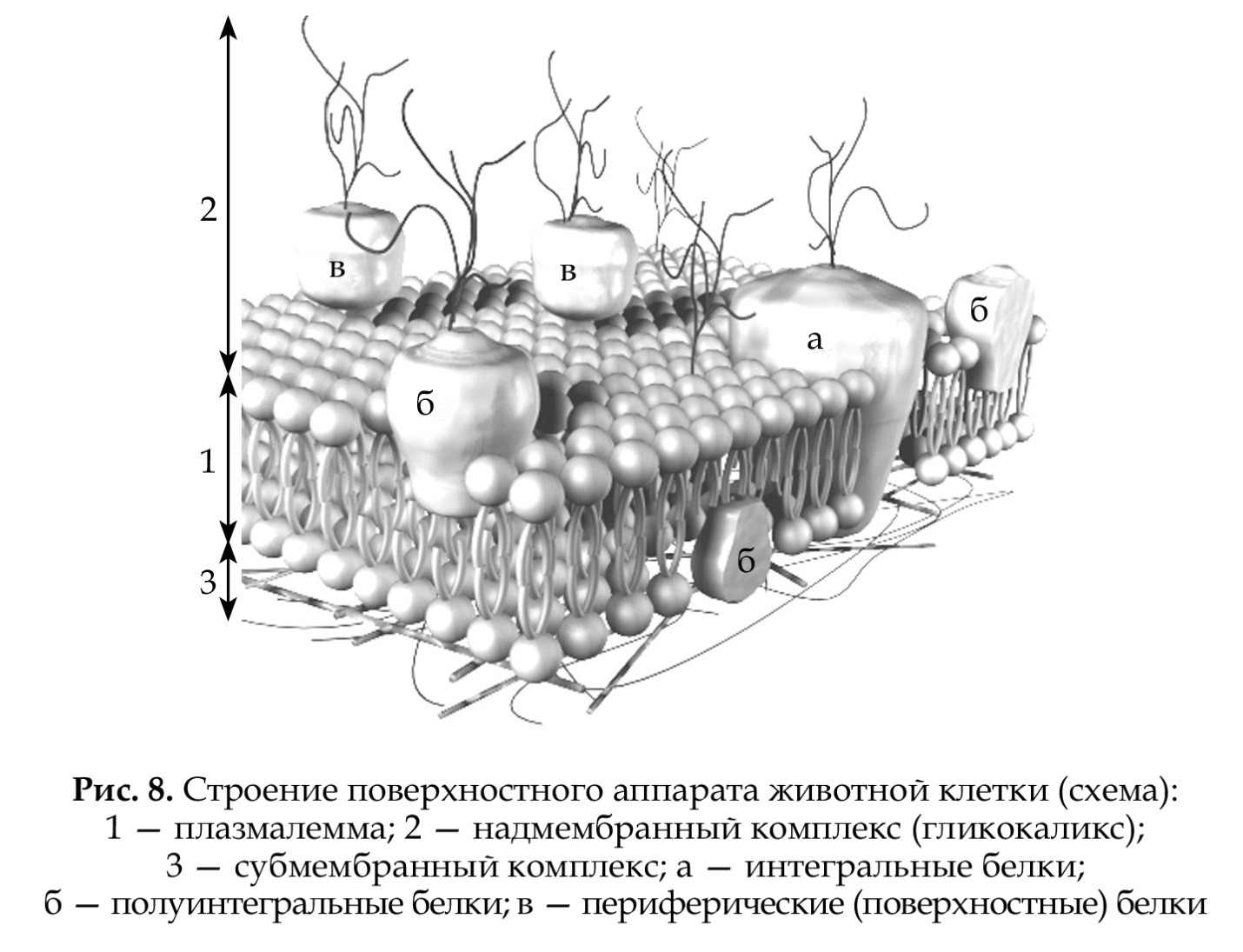 Традиционно все бактерии разделяют на две части, используя простой эмпирический критерий, называемый окраской по Граму, основанный на последовательной обработке бактерий тремя красителями (кристаллическим фиолетовым, йодом, сафранином). У грамположительных бактерий клеточные стенки содержат мало липидов, а у грамотрицательных клеточные стенки, напротив богаты липидами. И у тех и у других жесткий каркас образуется в результате ковалентного связывания полисахаридных и пептидных цепей. Однако, к этой структурной основе присоединяются некоторые специфические для данной бактерии компоненты, которые у грамположительных и грамотрицательных бактерий имеют разный состав. Грамотрица́тельные бакте́рии — это бактерии, которые не окрашиваются кристаллическим фиолетовым при окрашивании по Граму. В отличие от грамположительных бактерий, которые сохранят фиолетовую окраску даже после промывания обесцвечивающим растворителем (спирт), грамотрицательные полностью обесцвечиваются. После промывания растворителем при окрашивании по Граму добавляется контрастный краситель (обычно сафранин), который окрашивает все грамотрицательные бактерии в красный или розовый цвет. Это происходит из-за наличия внешней мембраны, препятствующей проникновению красителя внутрь клетки. Из-за своей более мощной и непроницаемой клеточной стенки грамотрицательные бактерии более устойчивы к антителам, чем грамположительные Характеристика
Грамположительные бактерии Грамположительные бактерии -бактерии, которые сохраняют окраску, не обесцвечиваются при промывке при использовании окраски микроорганизмов по методу Грама.имеют однослойную клеточную мембрану, без внешней мембраны. Большая часть патогенных для человека микроорганизмов относится к грамположительным. (стрептококки и стафилококки, являются кокками (шарообразными бактериями). Остальные — палочковидные и делятся далее по возможности образовывать споры. Неспорообразующие: Corynebacterium и Листерия; спорообразующие: Бациллы и Клостридии. Спорообразующие можно разделить на факультативных анаэробов Бациллы и облигатных анаэробов Клостридий. Гликокаликс - это обогащенная углеводами периферическая зона на поверхности большинства эукариотических клеток ( клеточная оболочка ). При использовании различных красителей, например рутениевого красного, эту зону можно отчетливо видеть в электронном микроскопе. Микротрубочки. Их строение. Расположение микротрубочковой субмембранной системы. Пластичность и динамичность микротрубочковой системы. Особенности тубулин-динеиновой системы ресничек и жгутиков Микротрубочки — белковые внутриклеточные структуры, входящие в состав цитоскелета.Микротрубочки представляют собой полые внутри цилиндры диаметром 25 нм. Длина их может быть от нескольких микрометров до, вероятно, нескольких миллиметров в аксонах нервных клеток. Их стенка образована димерами тубулина. Микротрубочки, подобно актиновым микрофиламентам, полярны: на одном конце происходит самосборка микротрубочки, на другом — разборка. В клетках микротрубочки играют роль структурных во многих клеточных процессах. Микротрубочки — это структуры, в которых 13 протофиламентов, состоящих из гетеродимеров α- и β-тубулина, уложены по окружности полого цилиндра. Внешний диаметр цилиндра около 25 нм, внутренний — около 15.Один из концов микротрубочки, называемый плюс-концом, постоянно присоединяет к себе свободный тубулин. От противоположного конца — минус-конца — тубулиновые единицы отщепляются. В образовании микротрубочки выделяют три фазы: замедленная фаза, или нуклеация. Это этап зарождения микротрубочки, когда молекулы тубулина начинают соединяться в более крупные образования. Такое соединение происходит медленнее, чем присоединение тубулина к уже собранной микротрубочке, поэтому фаза и называется замедленной; фаза полимеризации, или элонгация. Если концентрация свободного тубулина высока, его полимеризация происходит быстрее, чем деполимеризация на минус-конце, за счет чего микротрубочка удлиняется. По мере её роста концентрация тубулина падает до критической и скорость роста замедляется вплоть до вступления в следующую фазу; фаза стабильного состояния. Деполимеризация уравновешивает полимеризацию, и рост микротрубочки останавливается. Лабораторные исследования показывают, что сборка микротрубочек из тубулинов происходит только в присутствии гуанозинтрифосфата и ионов магния. Микротрубочки являются очень динамичными структурами, которые могут достаточно быстро возникать и разбираться. При использовании электронных систем усиления сигнала в световом микроскопе можно видеть, что в живой клетке микротрубочки растут, укорачиваются, исчезают, т.е. постоянно находятся в динамической нестабильности. Оказалось, что среднее время полужизни цитоплазматических микротрубочек составляет всего лишь 5 минут. Так за 15 мин около 80% всей популяции микротрубочек обновляется. В составе веретена деления микротрубоски имеют время жизни около 15-20 с. Однако 10-20% микротрубочек остаются относительно стабильными достаточно долгое время (до нескольких часов). За ретроградный транспорт в аксоне отвечает другой белок — цитоплазматический динеин . Он состоит из двух тяжелых цепей — головок, взаимодействующих с микротрубочками, нескольких промежуточных и легких цепей, которые связываются с мембранными вакуолями. Цитоплазматический динеин является моторным белком, переносящим грузы к минус-концу микротрубочек. Динеины также делятся на два класса: цитозольные, участвующие в переносе вакуолей и хромосом, и аксонемные, отвечающие за движение ресничек и жгутиков. Строение и движение ресничек. Микротрубочки цитоплазмы Реснички — многочисленные цитоплазматические выросты на поверхности плазмалеммы. Встречаются на поверхности клеток большинства животных и некоторых растений. Ресничка представляет собой тонкий цилиндрический вырост цитоплазмы с постоянным диаметром 300 нм. Этот вырост от основания до самой его верхушки покрыт плазматической мембраной. Внутри выроста расположена аксонема, сложная структура, состоящая в основном из микротрубочек. Нижняя, проксимальная часть реснички, базальное тельце, погружена в цитоплазму. Диаметры аксонемы и базального тельца одинаковы (около 200 нм). Клетка может двигатся вперед и назад ,вбок и по кругу.Движение ресничек сводится к колебаниям в одной плоскости,вызывающим перемещение клетки или тока жидкости,если реснички принадлежат некоторой эпителиальной ткани. Митохондрии. Общая морфология. Ультраструктура митохондрий Митохондрия - двумембранный сферический или эллипсоидный органоид диаметром обычно около 1 микрона. Характерна для большинства эукариотических клеток, как автотрофов так и гетеротрофов (грибы).Энергетическая станция клетки; основная функция — окисление органических соединений и использование освобождающейся при их распаде энергии для генерации электрического потенциала, синтеза АТФ и термогенеза. Эти три процесса осуществляются за счёт движения электронов по электронно-транспортной цепи белков внутренней мембраны. Количество митохондрий в клетках различных организмов существенно отличается[1]: так, одноклеточные зелёные водоросли эвглены имеют лишь одну гигантскую митохондрию; у кишечных анаэробных энтамёб и некоторых других паразитических простейших митохондрии отсутствуют. В специализированных клетках органов животных содержатся сотни и даже тысячи митохондрий (мозг, сердце, мышцы). Ультраструктура митохондрий Митохондрии ограничены двумя мембранами (рис. 205). Наружняя митохондриальная мембрана отделяет ее от гиалоплазмы. Обычно она имеет ровные контуры, не образует впячиваний или складок. На нее приходится около 7% от площади всех клеточных мембран. Толщина этой мембраны около 7 нм, она не бывает связана ни с какими другими мембранами цитоплазмы и замкнута сама на себя, так что представляет собой мембранный мешок. Наружную мембрану от внутренней отделяет межмембранное пространство шириной около 10—20 нм. Внутренняя мембрана (толщиной около 7 нм) ограничивает собственно внутреннее содержимое митохондрии, ее матрикс, или митоплазму. Характерной чертой внутренних мембран митохондрий является их способность образовывать многочисленные впячивания внутрь митохондрий. Такие впячивания чаще всего имеют вид крист . 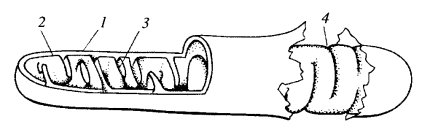 1 — внешняя мембрана; 2 — внутренняя мембрана; 3 — впячивания внутренней мембраны - кристы; 4 — места впячиваний, вид с поверхности внутренней мембраны Общая поверхность внутренней мембраны митохондрии в печеночной клетке составляет примерно треть поверхности всех клеточных мембран. Митохондрии клеток сердечной мышцы содержат втрое больше крист, чем печеночные митохондрии. Это может отражать различия в функциональных нагрузках митохондрий разных клеток. Расстояние между мембранами в кристе составляет около 10-20 нм. . Митохондриальные кристы, отходящие от внутренней мембраны и простирающиеся в сторону матрикса, обычно не полностью перегораживают полость митохондрии, не нарушают непрерывности заполняющего ее матрикса. Ориентация крист по отношению к длинной оси митохондрии различна для разных клеток. Так, может быть перпендикулярная ориентация (клетки печени, почек) крист; в некоторых клетках (сердечная мышца) наблюдается продольное расположение крист. Часто кристы могут ветвиться или образовывать пальцевидные отростки, изгибаться и не иметь выраженной ориентации . У простейших, одноклеточных водорослей, в некоторых клетках высших растений и животных выросты внутренней мембраны имеют вид трубок (трубчатые кристы). Матрикс митохондрий имеет тонкозернистое гомогенное строение, в нем иногда выявляются тонкие собранные в клубок нити (около 2—3 нм) и гранулы около 15—20 нм. Известно, что нити матрикса митохондрий представляют собой молекулы ДНК в составе митохондриального нуклеоида, а мелкие гранулы — митохондриальные рибосомы. Кроме того, в матриксе встречаются крупные (20—40 нм) плотные гранулы, это — места отложения солей магния и кальция. 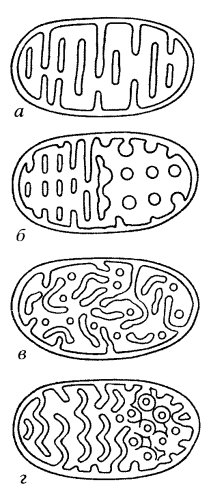 а — пластинчатые кристы (печень); б — перфорированные кристы (летательная мышца мухи); в — трубчатые кристы; г — волнистые кристы (амеба). а — пластинчатые кристы (печень); б — перфорированные кристы (летательная мышца мухи); в — трубчатые кристы; г — волнистые кристы (амеба).Функции митохондрий. Увеличение числа митохондрий. Авторепродукция митохондрий. Хондриом Функции митохондрий: 1) играют роль энергетических станций клеток. в них протекают процессы окислительного фосфорилирования (ферментативного окисления различных веществ с последующим накоплением энергии в виде молекул аденозинтрифосфата - АТФ) ; 2) хранят наследственный материал в виде митохондриальной ДНК. митохондрии для своей работы нуждаются в белкаx, закодированных в генах ядерной ДНК, так как собственная митохондриальная ДНК может обеспечить митохондрии лишь несколькими белками. Увеличение числа митохондрий. Митохондрии могут увеличивать свою численность особенно при делении клеток или при увеличении функциональной нагрузки клетки. Происходит постоянное обновление митохондрий. Например, в печени средняя продолжительность жизни митохондрий составляет около 10 дней.Основная масса экспериментальных данных говорит о том, что увеличение числа митохондрий происходит путем роста и деления предшествующих митохондрий. Было обнаружено , что в течение клеточного цикла митохондрии могут вырастать до нескольких мкм, а затем фрагментироваться, делиться на более мелкие тельца.Кроме того, митохондрии могут сливаться друг с другом. при делении митохондрий: происходит увеличение массы митохондриальных мембран со всеми специфическими компонентами за счет синтеза и включения в них отдельных белков - ферментов и липидов, нарастание массы белков матрикса, а затем происходит деление как бы удвоившейся или многократно увеличившейся структуры. Авторепродукция митохондрий. Двумембранные органеллы обладают полной системой авторепродукции. В митохондриях и пластидах существует ДНК, на которой синтезируются информационные, трансферные и рибосомные РНК и рибосомы, осуществляющие синтез митохондриальных и пластидных белков. Однако, эти системы, хотя и автономны, но ограничены по своим возможностям. ДНК в митохондриях представляет собой циклические молекулы без гистонов и тем самым напоминают бактериальные хромосомы. Найдено, что митохондральная ДНК различных объектов очень однородна.. Все митохондриальные ДНК представляют множественные копии, собранными в группы, кластеры. Так в одной митохондрии печени крысы может содержаться от 1 до 50 циклических молекул . В настоящее время доказано, что большая часть белков митохондрий находится под генетическим контролем со стороны клеточного ядра и синтезируется вне митохондрий. Большинство липидов митохондрий так же синтезируются в цитоплазме. Все это говорит о эндосимбиотическом происхождении митохондрий, о том, что митохондрии представляют собой организмы типа бактерий, находящиеся в симбиозе с эукариотический клеткой. Хондриом. Совокупность всех митохондрий в одной клетке называется хондриомом. Она может быть различной в зависимости от типа клеток. Во многих клетках хондриом состоит из разрозненных многочисленных митохондрий, равномерно расположенных по всей цитоплазме или локализуются группами в местах интенсивной траты АТФ. В обоих этих случаях митохондрии функционируют поодиночке, их кооперативная работа, возможно, координируется какими-то сигналами из цитоплазмы. Существует и совершенно иной тип хондриома, когда вместо мелких одиночных разрозненных митохондрий в клетке располагается одна гигантская разветвленная митохондрия ю.Такие митохондрии встречаются у одноклеточных зеленых водорослей (например у Chlorella). Они образуют сложную митохондриальную сеть или митохондриальный ретикулум Биологический смысл появления такой гигантской разветвленной митохондриальной структуры, объединенной в одно целое своими внешними и внутренними мембранами заключается в том, что в любой точке поверхности внутренней мембраны такой разветвленной митохондрии может идти синтез АТФ, который будет поступать в любую точку цитоплазмы, где в этом есть необходимость. С этих позиций митохондриальный ретикулум представляет собой как бы электрический проводник, кабель, соединяющий отдаленные точки такой системы. Митохондриальный ретикулум оказался очень полезным не только для мелких подвижных клеток, таких как хлорелла, но и для более крупных структурных единиц таких как, например, миофибриллы в скелетных мышцах. Хондриом клеток сердечной мышцы не образует ветвящихся структур, а представлен множеством небольших вытянутых митохондрий, располагающихся без особого порядка между миофибриллами. Однако, все соседние митохондрии стыкуются друг с другом с помощью митохондриальных контактов . |