Биосинтез белка_студентам_материал_2020. Нуклеиновые кислоты Нуклеиновые кислоты
 Скачать 0.83 Mb. Скачать 0.83 Mb.
|

|
Биосинтез белка и его значение в биотехнологии. Создание рекомбинантных организмов. Нуклеиновые кислоты Нуклеиновые кислоты – это высокомолекулярные биополимеры, мономерами которых являются нуклеотиды. Функция нуклеиновых кислот заключается в хранении, реализации и передаче генетической (наследственной) информации в живых организмах. Общая формула нуклеотида Остаток фосфорной кислоты + углевод (пентоза) + азотистое основание Два нуклеотида соединяются фосфодиэфирной связью, по аналогии как две аминокислоты – пептидной. Существует два типа нуклеиновых кислот: • ДНК (дезоксирибонуклеиновая кислота) • РНК (рибонуклеиновая кислота). ДНК: Остаток фосфорной кислоты + дезоксирибоза + азотистое основание (аденин, гуанин, цитозин, тимин) РНК: Остаток фосфорной кислоты + рибоза + азотистое основание (аденин, гуанин, цитозин, урацил) Основания принято обозначать первой буквой их названия. Различают пуриновые (А, Г) и пиримидиновые (Т, У, Ц) основания.  Строение ДНК. Молекулы ДНК в основном находятся в ядрах клеток, но небольшое их количество содержится в митохондриях и пластидах. ДНК человеческой клетки умещается в ядре диаметром 0.005 мм. Если полностью развернуть спираль ДНК, то длина составит почти два метра. Все ДНК одного тела, вытянутые в нить, выстроились бы на расстояние, равное 40-ка расстояниям от Земли до Солнца. ДНК человека на девяносто восемь процентов такая же, как и у шимпанзе, и даже на 50 процентов такая же, как у банана. По меньшей мере 8 процентов генома человека порождено вирусами, чей генетический код объединился с нашим в течении около 40 миллионов лет эволюции приматов. Нуклеиновые кислоты подобно белкам имеют • первичную структуру – последовательность нуклеотидов. Расположение нуклеотидов важно, так как задает последовательность аминокислот в кодируемых белках. • вторичную структуру – две комплементарные цепи • третичную – пространственную структуру, которую и установили Уотсон и Крик (в 1953 г – этот год считается годом выделения молекулярной биологии в самостоятельную науку). Молекула ДНК состоит из двух спирально закрученных цепей, которые по всей длине соединены друг с другом водородными связями. Такую структуру, свойственную только молекулам ДНК, называют двойной спиралью.  Правила Чаргаффа: 1) В любой молекуле ДНК количество аденина равно числу тимина, а число гуанина= числу цитозина. А = Т, Г = Ц. 2) А + Г = Т + Ц Между аденином и тимином всегда возникают две, а между гуанином и цитозином — три водородные связи, В связи с этим обнаруживается важная закономерность: против аденина одной цепи всегда располагается тимин другой цепи, против гуанина — цитозин и наоборот. 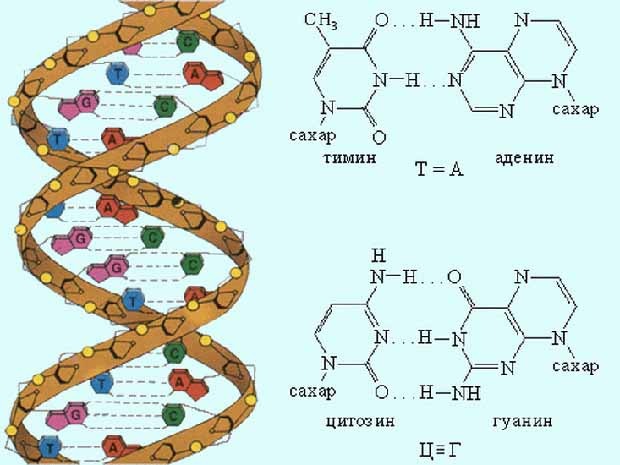 Таким образом, пары нуклеотидов аденин и тимин, а также гуанин и цитозин строго соответствуют друг другу и являются комплементарными. Три нуклеотида – триплет или кодон несут информацию об одной аминокислоте. Основным свойствомДНКявляется ее способность к репликации. 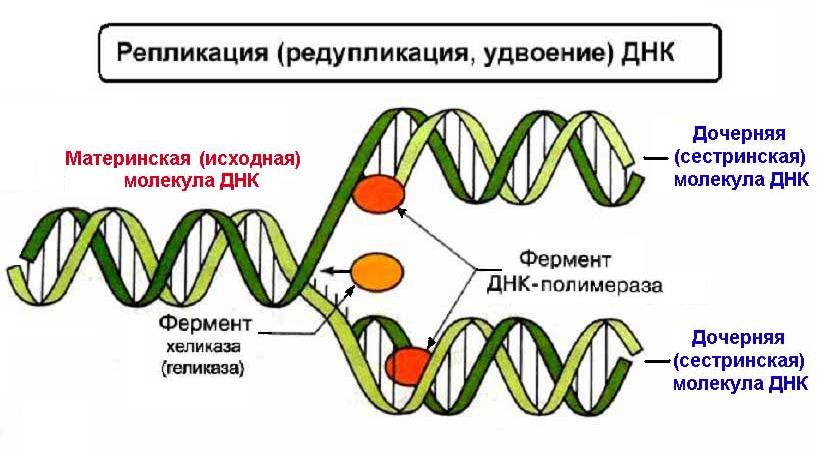 Репликация — это процесс самоудвоения молекул ДНК, происходящий под контролем ферментов. Репликация осуществляется перед каждым делением ядра. Топоизомеразы облегчают расплетание цепей ДНК в двойной спирали (снимают сверхспирализацию некоторых участков ДНК путем внесения одно- или двухцепочечного разрыва с последующим восстановлением – лигированием). Топоизомеразы расплетают небольшие участки ДНК непосредственно перед репликативной вилкой и «подготавливают» ДНК к расплетанию. ДНК-хеликазы (ДНК-геликазы) разделяют цепи двухцепочечной молекулы ДНК, используя энергию гидролиза высокоэнергетических соединений (АТФ или ГТФ). Образуется две материнские цепи ДНК. Далее на каждой из цепей под действием ДНК-полимеразы синтезируется дочерняя цепь ДНК. В итоге дочерние молекулы ДНК являются точными копиями материнских. Это полуконсервативный механизм. Экспериментально это было доказано Мезельсоном и Сталем (1958). E.coli выращивали в среде, содержащей в качестве источника азота только его тяжелый изотоп 15N (изотопы – атомы, которые имеют одинаковое число протонов, но разное – нейтронов, т.е. одинаковый порядковый номер, но разное массовое число). Эта метка включалась в ДНК, вследствие чего ДНК в клетках, выращенных среде с 15N, имела большую молекулярную массу на единицу объема (т.е. большую плотность), чем ДНК в клетках, выращенных в обычных условиях, в присутствии легкого изотопа 14N. Поэтому, если клетки после длительного выращивания на среде с 15N отмывали и переносили в среду с 14N вместо 15N, то это должно было привести к появлению молекул ДНК с меньшей плотностью. Различающиеся по плотности молекулы можно разделяли ультрацентрифугированием. Оказалось, что в клетках появилась гибридная («средняя») по плотности ДНК, у которой одна цепь была “тяжелой”, а другая - “легкой”. 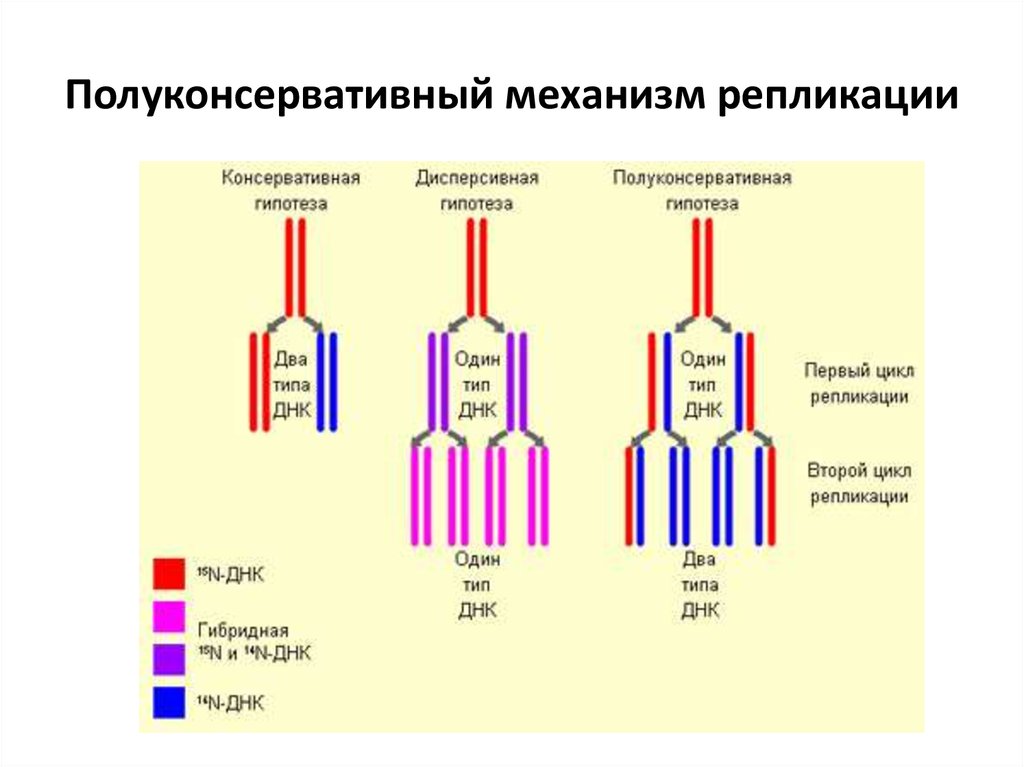 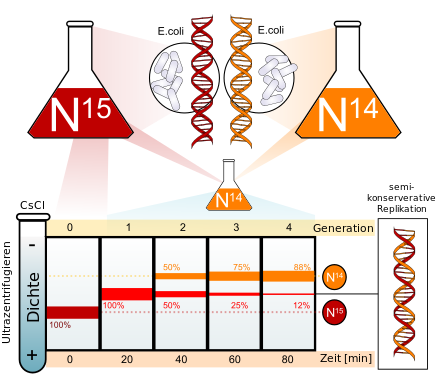 Биологический смысл репликации заключается в точной передаче наследственной информации от материнских клеток дочерним. Особенностью процесса репликации является возможность исправления ошибок (корректорская правка). ДНК-полимераза каждый раз дважды проверяет правильность нуклеотида (измеряет расстояние) в своем активном центре, и ее полимеразная активность включается лишь тогда, когда эта комплементарность установлена. С другой стороны, каждый вновь встроенный нуклеотид также проверяется на соответствие своей паре в активном центре фермента. Если размер образовавшейся пары нуклеотидов не соответствует истинному (когда основания противоположных нуклеотидов не комплементарны друг другу), с помощью своей 3’→5’-экзонуклеазной активности фермент вырезает некомплементарный нуклеотид и ищет ему замену. Дополнительным механизмом, уменьшающим ошибки репликации, служит репарация ДНК. В результате частота ошибочного включения нуклеотидов в образующуюся при репликации цепь ДНК крайне низка (10-8—10-10). Синтез дочерней цепи ДНК всегда антипаралелен материнской (принцип антипаралельности). Синтез всегда происходит в направлении 3→5 (принцип униполярности). Синтез комплиментарных цепей – наоборот. Ген – участок молекулы ДНК, содержащий информацию о первичной структуре белка. ДНК может повреждаться различными мутагенами и радиацией. Они вызывают различные изменения в структуре ДНК, что мешает репликации и транскрипции и вызывает мутации. Некоторые мутагены используют в химиотерапии для подавления быстрорастущих клеток рака. Фрагменты ДНК можно разделить с помощью электрофореза. Поскольку любая молекула ДНК в водном растворе отрицательно заряжена, смесь фрагментов ДНК различных размеров можно разделить с помощью электрофореза. ДНК помещают в гель (агарозный, полиакриламидный), который помещают в постоянное электрическое поле. Молекулы ДНК будут двигаться к положительному электроду (аноду), причем их скорости будут зависеть от длины молекулы: чем она длиннее, тем сильнее ей мешает двигаться гель и, соответственно, тем ниже скорость. После электрофореза смеси фрагментов разных длин в геле образуют полосы, соответствующие фрагментам одной и той же длины. С помощью маркеров (смесей фрагментов ДНК известных длин) можно установить длину молекул в образце. Визуализовать результаты электрофореза можно двумя способами: • добавлением в гель веществ, флуоресцирующих в присутствии ДНК (традиционно использовался довольно токсичный бромистый этидий; в последнее время в обиход входят более безопасные вещества). Бромистый этидий светится оранжевым светом при облучении ультрафиолетом, причем при связывании с ДНК интенсивность свечения возрастает на несколько порядков. • использовать радиоактивные изотопы, их надо предварительно включить в состав анализируемой ДНК. В этом случае на гель сверху кладут фотопластинку, которая засвечивается над полосами ДНК за счет радиоактивного излучения. Это метод называется авторадиографией. 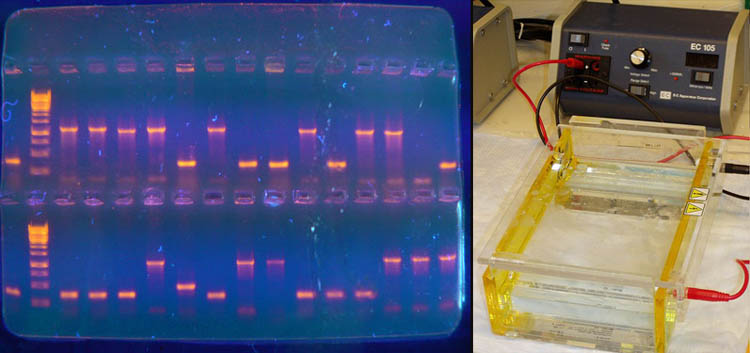 Кроме «обычного» электрофореза в пластине из геля, в некоторых случаях используют капиллярный электрофорез, который проводят в очень тонкой трубочке, наполненной гелем (обычно полиакриламидным). Разрешающая способность такого электрофореза значительно выше: с его помощью можно разделять молекулы ДНК, отличающиеся по длине всего на один нуклеотид. РНК РНК состоит из одной цепи, закрученной в спираль (исключение – некоторые вирусы содержат двухцепочечные РНК). Молекулы РНК короче, чем молекулы ДНК. Типы РНК 1) Информационные, или матричные, РНК (иРНК) составляют около 5% всей клеточной РНК. Они синтезируются на участке одной из цепей молекулы ДНК и передают информацию о структуре белка из ядра клеток к рибосомам, где эта информация реализуется. В зависимости от объема копируемой информации молекула иРНК может иметь различную длину. иРНК это копия участка ДНК. Молекулы иРНК относительно нестабильные – быстро распадаются на нуклеотиды. Срок их жизни составляет до несколько часов. Транскрипция – синтез иРНК на ДНК. Трансляция – синтез белка на иРНК. Основная догма молекулярной биологии: ДНК → РНК → белок.  2) Транспортные РНК (тРНК) образуются в ядре на ДНК, затем переходят в цитоплазму. Они составляют около 10% клеточной РНК и являются самыми небольшими по размеру РНК, состоящими из 70— 100 нуклеотидов. Каждая тРНК присоединяет определенную аминокислоту и транспортирует ее к месту синтеза белка в рибосоме. Там молекула тРНК узнает соответствующий участок иРНК. Таким образом, определяется порядок расположения аминокислотных остатков в молекуле белка, который синтезируется. Все известные тРНК за счет комплементарного взаимодействия образуют вторичную структуру, по форме напоминающую лист клевера. В молекуле тРНК есть два активных участка: триплет-антикодон на одном конце и акцепторный конец на другом. 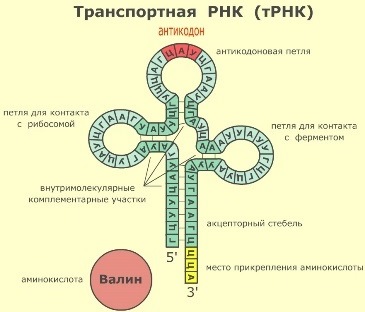 На верхушке «листка» – триплет (три нуклеотида), который по генетическому коду соответствует определенной аминокислоте. Этот триплет называют антикодоном. Он комплементарен соответствующему триплету ДНК (кодону). Поскольку в трёхбуквенных комбинациях используются 4 основания, всего возможны 64 кодона (4³ комбинации). Из них 61 кодирующий и 3 некодирующих (кодоны-терминаторы, свидетельствуют об окончании процесса трансляции). Кодоны-терминаторы в -РНК: УАА, УАГ, УГА, в ДНК: АТТ, АТЦ, АЦТ. Кодоны кодируют 20 стандартных аминокислот, каждой из которых соответствует в большинстве случаев более одного кодона. Возле основания есть участок (акцепторный конец), к которому благодаря ковалентной связи прикрепляется молекула аминокислоты. Молекула тРНК может образовывать и третичную структуру (L – подобную). 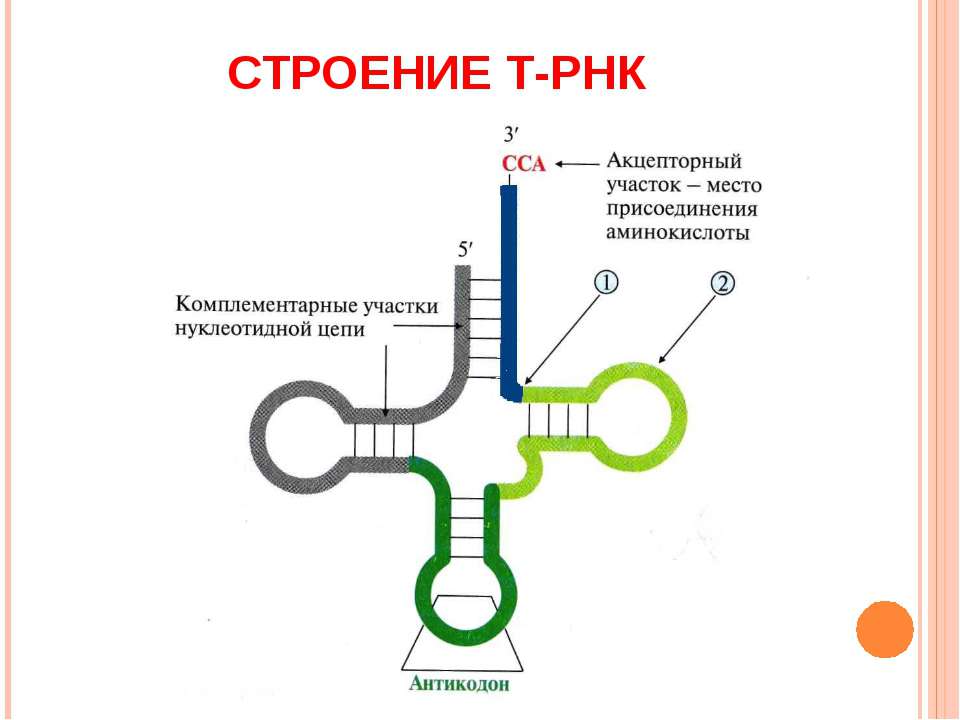 3) Рибосомные РНК (рРНК) синтезируются в основном в ядрышке и составляют примерно 85% всех РНК клетки. Они входят в состав рибосом и участвуют в формировании активного центра рибосомы, где происходит процесс биосинтеза белка. Не принимают участия в передаче наследственной информации. Выполняет структурную функцию – обеспечивает определенное пространственное расположение иРНК и тРНК на рибосоме.  Биосинтез белка Начало процесса трансляции определяет кодон-инициатор (АУГ, в ДНК — ТАЦ), кодирующий аминокислоту метионин. Этот кодон первым входит в рибосому. Впоследствии метионин, если он не предусмотрен в качестве первой аминокислоты данного белка, отщепляется. Каждая т-РНК может переносить только свою аминокислоту. Т-РНК активируется специальными ферментами, присоединяет свою аминокислоту и транспортирует ее в рибосому. Внутри рибосомы в каждый данный момент находится всего два кодона и-РНК. Если антикодон т-РНК является комплементарным кодону и-РНК, то происходит временное присоединение т-РНК с аминокислотой к и-РНК. Ко второму кодону присоединяется вторая т-РНК, несущая свою аминокислоту. Аминокислоты располагаются рядом в большой субъединице рибосомы, и с помощью ферментов между ними устанавливается пептидная связь. Одновременно разрушается связь между первой аминокислотой и ее т-РНК, и т-РНК уходит из рибосомы за следующей аминокислотой. Рибосома перемещается на один триплет, и процесс повторяется. Так постепенно наращивается молекула полипептида, в которой аминокислоты располагаются в строгом соответствии с порядком кодирующих их триплетов (матричный синтез). Одна рибосома способна синтезировать полную полипептидную цепь. Однако, нередко по одной молекуле и-РНК движется несколько рибосом. Такие комплексы называются полирибосомами. После завершения синтеза полипептидная цепочка отделяется от матрицы – молекулы и-РНК, сворачивается в спираль и приобретает свойственную ей (вторичную, третичную или четвертичную) структуру. Рибосомы работают очень эффективно: в течение 1с бактериальная рибосома образует полипептидную цепь из 20 аминокислот. Синтез белка (трансляция) можно условно разделить на 2 этапа: активирование аминокислот. Необходимым условием синтеза белка является наличием в системе не свободных, а так называемых активированных аминокислот, располагающих своим внутренним запасом энергии. Активация свободных аминоксилот осуществляется при помощи аминоацил –тРНК–синтетаз в присутствии АТФ (при этом образуется аминоацил-тРНК). собственно процесс трансляции (протекает в рибосоме) инициация (связывание рибосомы с мРНК, нахождение стартового кодона, кодирующего метионин) элонгация (присоединение новых аминоацил-тРНК, образование пептидной связи, шаг рибосомы на новый триплет и т.д.) терминация (вспомогательные белки – факторы терминации – узнают стоп-кодоны и связываются в рибосоме на место тРНК, при этом происходит гидролиз связи синтезированного пептида с тРНК) Пептидная связь образуется между карбоксильной группой одной аминокислоты и аминогруппой другой. Первая аминокислота белковой молекулы имеет свободную аминогруппу (N-конец), а последняя – свободную карбоксильную группу (С-конец). Организация генов прокариот Прокариоты не имеют оформленного ядра. У них есть нуклеоид – кольцевая молекула ДНК. В геноме прокариот выделяют: • независимые гены – гены, с каждого из которых считывается моноцистронная РНК. Цистрон – это последовательность нуклеотидов, кодирующая единую полипептидную цепь или стабильную РНК. Среди независимых генов выделяют гены, содержащие регуляторные области и конститутивные. Конститутивные – гены, транскрипция которых осуществляется непрерывно. Эти гены кодируют структуру конститутивных белков (тех белков, которые синтезируются постоянно, например гистоновые). • гены, объединенные в группы Оперон – совокупность генов, способных включаться и выключаться в зависимости от метаболических потребностей клеток. Кроме нуклеоида в клетках прокариот часто есть плазмиды – кольцевые молекулы ДНК, способные автономно размножаться. От наличия плазмид зависит устойчивость к антибиотикам. Очень важно, что плазмиды из-за своих маленьких размеров могут быть выделены из клеток неповрежденными. 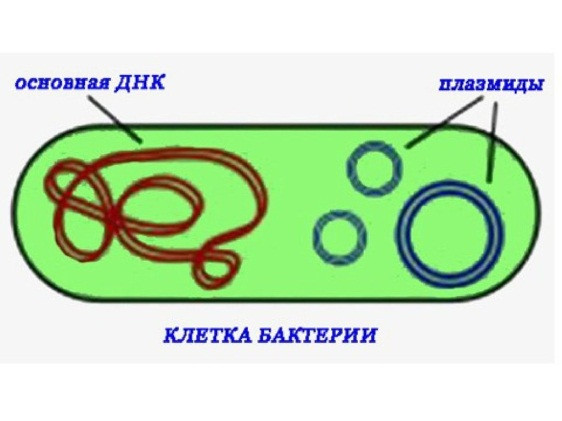 Наследственный материал эукариот Все клетки эукариот имеют ядро, в нем сохраняется наследственная информация. Есть клетки, которые имеют несколько ядер (это клетки больших размеров или клетки с ускоренным обменом веществ). Есть клетки у которых ядро разрушается в процессе развития, они не способны к размножению (тромбоциты, эритроциты). На самом деле наряду с информацией о структуре белка, записанной с помощью генетического кода, для успешного проведения синтеза белка, молекула ДНК должна иметь ряд регуляторных сигналов, записанных в виде специфических нуклеотидных последовательностей. Эти последовательности указывают точки начала транскрипции и трансляции и указывают точки прекращения считывания генетической информации. Гены эукариотических организмов имеют мозаичную структуру. Кодирующие белок участки (экзоны) разделены не кодирующими участками (интронами). В клетках эукариот информационная РНК, которая транскрибируется с гена, кодирующего белок, подвергается расщеплению ферментами эндонуклеазами. В результате этого процесса из м-РНК удаляются все интроны, а экзоны в строго определенной последовательности сшиваются при помощи лигазы. Этот процесс получил название сплайсинга. В результате сплайсинга и последующего созревания м-РНК обладает непрерывной последовательностью нуклеотидов, кодирующей последовательность аминокислот белка, а также содержит все регуляторные сигналы, необходимые для начала и прекращения трансляции. 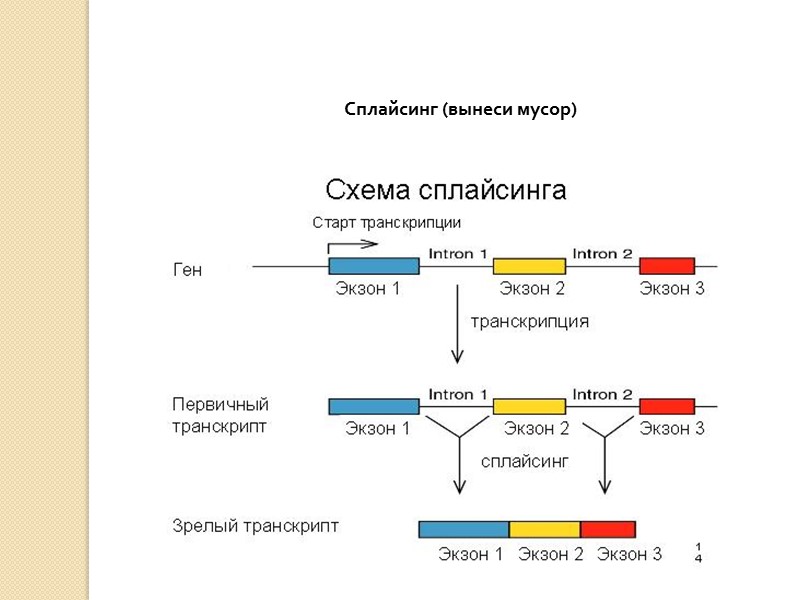 У прокариот отсутствует сплайсинг и их гены не содержат интронов, то есть последовательности нуклеотидов, кодирующие белки у прокариот изначально непрерывны. Следовательно, интроны генов эукариот не могут быть удалены в бактериальных клетках. Поэтому для экспрессии в бактериальной клетке гена человека и других эукариот необходимо, чтобы в бактериальную клетку была введена ДНК, интроны из которой уже удалены. Этого можно достичь, если из клеток человека выделить не сам ген, а транскрибированную с него м-РНК, уже прошедшую сплайсинг, а затем при помощи особого фермента РНК-зависимой-ДНК-полимеразы на матрице м-РНК получит ДНК. Более того регуляторные сигналы в ДНК, которую предполагается ввести в клетку кишечной палочки, должны быть похожими на таковые бактериальной клетки. В 1970 году американские ученые Келли и Смит с сотрудниками выделили первую рестриктазу – фермент, который вызывает гидролиз ДНК в строго определенных местах. Место, в котором рестриктазы разрывают ДНК, определяется последовательностью, как правило, из 6-8 нуклеотидов. В настоящее время описано более 300 различных рестриктаз. Многие из них применяются в генной инженерии. |