Нейрофизиологические исследования бесконтактного внесенсорного взаимодействия как объективной реальности энергоинформационных св. Нейрофизиологические исследования бесконтактного внесенсорного в. О. И. Коёкина Рассматриваются полученные на протяжении последних лет результаты нейрофизиологических исследований бесконтактных внесенсорных связей и возможность их рассмотрения как межиндивидуального энергоинформа
 Скачать 113.5 Kb. Скачать 113.5 Kb.
|

|
Нейрофизиологические исследования бесконтактного внесенсорного взаимодействия как объективной реальности энергоинформационных связей виртуального мозга О.И.Коёкина Рассматриваются полученные на протяжении последних лет результаты нейрофизиологических исследований бесконтактных внесенсорных связей и возможность их рассмотрения как межиндивидуального энергоинформационного взаимодействия в виртуальной реальности. В конкретных случаях межиндивидуальные энергоинформационные связи определяются в процессе регистрации биопотенциалов мозга одновременно у индуктора и реципиента путем применения математического аппарата спектрального и корреляционного анализа. Показано, что динамические структуры межиндивидуальных энергоинформационных связей попеременно объединяют различные области мозга как индуктора, так и реципиента, образуя виртуальный мозг со свойствами виртуальной реальности, при этом структуры связей между областями виртуального мозга в определенном частотном диапазоне отличаются от таковых как у индуктора, так и у реципиента в том же диапазоне. Наиболее обоснованные c методической точки зрения нейрофизиологические исследования по уточнению возможности бесконтактного внесенсорного взаимодействия проводились еще в 60-х годах в рамках научной тематики АМН, связанной с обнаружением слабых реакций центральной нервной системы на внешний раздражитель. Под внесенсорным бесконтактным взаимодействием тогда понимали возможность реакции одного испытуемого (реципиента) в ответ на какие-либо, например, стрессовые или болевые, реакции другого испытуемого (индуктора) при отсутствии известных сенсорных контактов. Полная сенсорная разобщенность между ними достигалась путем удаления реципиента от индуктора на несколько километров, при этом исследования проводились в помещениях, экранированных от электромагнитных помех и оборудованных для регистрации электроэнцефалограммы (ЭЭГ). В этих условиях было показано статистически достоверно, что у реципиента возникают слабые реакции в виде неспецифической активации биоритмов мозга с некоторым запаздыванием в секундах по отношению к началу реакций у индуктора. Исследователи подчеркивали, однако, что длительные латентные периоды, скорее всего, означают характерную для подпороговых сигналов задержку ответных реакций центральной нервной системы [1]. В настоящее время в публикуемых работах такой вид взаимодействия обычно называют энергоинформационным, а лиц, участвующих в нем - экстрасенсами, телепатами, сенситивами, биоэнергетиками, целителями и т.д. В данной статье мы останавливаемся на определении бесконтактного внесенсорного воздействия (восприятия, взаимодействия) как энергоинформационного, учитывая роль индуктора, как генератора, и реципиента, как приемника, этого воздействия. Цель работы - получение фактического материала для изучения характеристик биоритмов мозга, связанных со специфическими способностями участвующих лиц. Несмотря на своевременность и актуальность проблемы, на настоящий момент имеется довольно ограниченное количество опубликованных работ, связанныx с нейрофизиологическими исследованиями такого взаимодействия. Большинство исследователей останавливает свое внимание на объективизации состояния мозга самого экстрасенса в процессе “экстрасенсорного” восприятия. Эта задача решалась в ходе опытов регистрации ЭЭГ разными путями: во время биолокации и кожно-оптического гнозиса [2], диагностики заболеваний по фотографии человека [3], направленного воздействия и медитации [4], при дистантном взаимодействии, оцениваемом по тепловому потоку с поверхностей ладоней рук [5], во время сеансов установления телепатической связи [6], ясновидения в режиме ретрокогниции [7], целительства - экстрасенсорной диагностики и “скрытого” воздействия [8] и др. При этом, накопленный нейрофизиологический опыт давал возможность предполагать, что ключ к пониманию нейродинамических процессов, обеспечивающих у экстрасенсов повышенную, по сравнению с другими, чувствительность, может быть заложен в представлениях о неосознаваемых психических процессах и способах переработки информации, отражающихся в нейродинамическом кодировании биоритмами мозга. Эти нейродинамические процессы представляются в виде феноменов синхронизации и перераспределения очагов ритмической активности в коре головного мозга, что полностью согласуется с ранее проведенными исследованиями, показавшими, что в соответствии с поведенческими актами и психическими реакциями осуществляется сонастройка множества дистантно разрозненных нервных образований [9]. К настоящему времени было установлено также, что подсознательный процесс, по каким бы законам он ни протекал, находит свое отражение в электрической активности мозга. Он готовится постепенно на неосознаваемом уровне и уже тогда может быть выявлен электрографически, хотя лишь только на последнем этапе достигает сознания и вербализуется [10]. В качестве основы субъективных переживаний был рассмотрен механизм взаимодействия жестких и гибких связей, которые определяются совпадением частотных характеристик и соотношением фаз колебаний биопотенциалов мозга. Было показано, что фокусы взаимодействия со специфической для каждого вида психической деятельности топографией возникают за счет конвергенции гибких связей на разных частотах, определяя в то же время свою внутреннюю структуру жесткими связями [11]. Таким образом, благодаря применению топического картирования ЭЭГ и распределения функциональных связей между областями мозга по показателям кросскорреляции и когерентности биопотенциалов, стало возможным изучение специфичности нейродинамических структур, сопутствующим не только осознаваемым, но и неосознаваемым процессам. Полученные благодаря использованию моделей интеллектуальной деятельности и гипнотического состояния сознания нейрофизиологические данные позволили также определить уровень сознания как фактор, модулирующий организацию активационных процессов в коре головного мозга [2]. Исследования динамики ЭЭГ у экстрасенсов показали значительное возрастание степени синхронизации корковых потенциалов разных диапазонов частот, особенно в теменно-центральных областях коры, хотя при невысоких значениях средней когерентности ее максимум мог наблюдаться и в лобных областях [12]. Установлено, что локальная синхронизация в специфических и в проекционных сенсорных зонах коры соответствует более успешной реализации экстрасенсорного восприятия, тогда как глобальная синхронизация (по всей коре) отражает уровень напряженности интеллектуальной работы, препятствующей этому виду восприятия. При биолокации и кожно-оптическом гнозисе очаги активности чаще выражены в задних, реже в передних и центральных зонах, и более вариабельны в правом полушарии. Успешность деятельности коррелирует с возникновением очагов повышенной активности в передних областях левого полушария и в задних правого. Этот эффект связывают с процессами селекции информации и ее осознаванием с участием лобного левого очага [12]. При исследовании реакций активации на ЭЭГ как у индуктора, так и у реципиента найдено [6], что при бесконтактной внесенсорной связи, возникающей на значительном расстоянии (Москва-Ленинград), оптимальным типом кортикальной активационной структуры является смещение фокуса максимальной активации сначала в лобно-правые, а далее в задне-левые отделы, но в момент операционального действия у профессионалов наблюдается полная сопряженная инверсия градиентов кортикальной активации от лобно-левой ориентации к задне-правой, что отражает перестройку режима мозговой деятельности, повидимому, направленную на процесс осознавания, как показано в предыдущей работе. При визуальном анализе ЭЭГ у лиц, обладающих экстрасенсорными способностями, большинство исследователей отмечают общее снижение индекса альфа-ритма и его амплитуды, расценивая это наблюдение как преобладание процессов визуализации в экстрасенсорном восприятии с участием лимбико-стволовой активности. На этом фоне нарастание количества коротких альфа-вспышек или генерализованных заостренных одиночных альфа-волн, больше в правом полушарии, указывают, по мнению авторов, на наличие усиленно функционирующих очагов, больше справа, в области таламуса. Другие наблюдаемые изменения ЭЭГ в виде периодического появления групп тета-волн в лобных отделах, усиления или ослабления бета-ритма указывают на участие, определяемых с помощью специальной компьютерной программы по локализации очагов активности, лобно-базальных, мезодиэнцефальных структур, по-видимому, обеспечивающих при вовлечении отдельных областей гипоталамуса, гиппокампа и других структур системы свода, сферу эмоциональной деятельности [5,8]. Разнообразные методические приемы для выяснения особенностей межполушарной асимметрии мозга при экстрасенсорном восприятии и бесконтактном воздействии также позволили раскрыть различные особенности межполушарной асимметрии в работе головного мозга, более частую встречаемость леворукости у сенситивов и другие показатели повышенной активности правого полушария [13]. Нередко высказывается предположение о том, что переход в измененное состояние сознания (ИСС), характерное для сенситива, сходно с переходом в гипнотическое состояние сознания. Отмечается также, что состояния релаксации и гипнотического транса способствуют усилению и концентрации специфических очагов активности, и высокогипнабельные испытуемые показывают более совершенную нейрофизиологическую основу для внесенсорного бесконтактного взаимодействия [12]. Что касается оказания бесконтактного энергоинформационного воздействия, то с помощью специально разработанной методики, направленной на синхронизацию активной деятельности одновременно у нескольких лиц в гипнотическом состоянии сознания, показана возможность воспроизводимости влияния на динамические характеристики (скорость вращения) отдельных механических устройств типа “вертушки” [14]. Особенностями ИСС у сенситива, очевидно, является снижение порогов восприятия, и не исключено, что по нескольким видам ощущений, а также своеобразный характер осознания этих ощущений в виде образов, нередко более ярких, чем реальные. Однако, эти образы, как правило, продуцируются в зависимости от социального и образовательного положения сенситива, трансформируя неясные подсознательные ощущения в более для него привычные понятийные формы. Например, у народного целителя-сенситива, не имеющего медицинского образования, образы, характеризующие нарушение здоровья, могут быть представлены такими понятийными формами, как “медуза” при наличии отека в тканях, “плотная паутина”, пронизывающая орган, - при затяжных заболеваниях и т.д. Переход от одного состояния сознания к другому может происходить скачкообразно. Впервые это было обнаружено по изменениям сверхмедленных колебаний потенциалов мозга в момент перехода от обычного бодрствования в гипнотическое состояние сознания [15]. Как было показано, при профессиональной работе индуктора во время внесенсорного бесконтактного взаимодействия переход от одного вида деятельности к другому происходит динамически и произвольно управляется. Этот процесс отражается в попеременной инверсии лобно-затылочных отношений правого и левого полушарий, свидетельствующей о переключении интуитивных неосознаваемых на осознаваемые реакции. Чтобы определить это состояние как ИСС, необходимо было бы выделить стабилизирующие процессы, характеризующие ИСС. Однако, в отношении профессиональной деятельности индукторов, судя по данным ЭЭГ, отсутствует стабилизация тех проявлений межполушарной асимметрии, которая характерна как для интуитивных, так и для осознаваемых процессов. У индуктора, находящегося в рабочем состоянии, оба типа реакций сосуществуют динамически, сменяя друг друга. При этом, большинство исследователей отмечают преобладание реакций активации в коре обоих полушарий с более четкими очагами во время успешного взаимодействия. Наблюдения за работой целителей с пациентами показали также, что подобные дискретные изменения состояния сознания происходят и у пациентов. Более того, в некоторых случаях сами пациенты начинают обнаруживать у себя после процедур лечения у целителей новые, ранее не замечавшиеся способности к необычному для них восприятию. Эти и подобные им факты, побудившие к проведению одновременной регистрации биопотенциалов мозга как у целителя, так и у пациента в процессе целительской деятельности, открыли область исследований, которую мы отнесли к виртуальной реальности. Речь идет о взаимодействии между отдельными структурами мозга целителя (как индуктора), с одной стороны, и отдельными областями мозга пациента (как реципиента), с другой стороны. Это взаимодействие выражается в синхронизации биопотенциалов мозга в узкополосном спектре, присущем каждый раз определенной паре образований мозга целителя и пациента. Динамическая природа этого взаимодействия выражается в попеременной смене частоты, фазы, областей мозга. Физический носитель биоинформации остается неизвестным. Однако, согласно выделенным критериям, описанным ранее [4,16], можно судить о наличии между ними биоинформационных связей. При этом вновь возникающая динамическая структура межиндивидуального биоинформационного взаимодействия формируется как виртуальная реальность, имеющая свою область существования, свои особенности и свои ограничения. Сложность межиндивидуального взаимодействия сопоставима с работой отдельного головного мозга и повторяет ее закономерности, что позволяет рассматривать эту реальность как виртуальный мозг, который функционирует, включая в деятельность разные области мозга как целителя, так и пациента. Последующие исследования показали, что биоинформационные связи виртуального мозга весьма динамичны. Виртуальный мозг имеет свои свойства и свои закономерности развития биоинформационных связей, их взаимодействие относительно независимо от индивидуальной работы мозга целителя и пациента в отдельности. Чтобы показать это, остановимся коротко на методике проведения исследования. В группу исследования входили лица, владеющие экстрасенсорной диагностикой, целительством и другими способами биоэнергоинформационного взаимодействия - 18 целителей (назовем их индукторами) и их пациенты (назовем их реципиентами). Во время проведения исследований был использован прием одновременного съема биоэлектрической активности мозга у двух лиц (индуктора и реципиента). Использовали 8-канальную систему монополярных отведений от лобных - F3, F4, центральных - C3, C4, теменных - P3, P4 и затылочных - O1, O2 областей коры соответственно левого и правого полушарий по отношению к индифферентным электродам в виде объединенных пар ушных электродов у каждого участника исследования. Биопотенциалы мозга через аналогоцифровой преобразователь вводили в компьютер для проведения дальнейшей математической обработки данных. Оба участника находились рядом, на расстоянии около 80 см друг от друга, но без непосредственных контактов. Возможность их возникновения во время регистрации ЭЭГ контролировали по возникновению артефактов и путем непосредственного наблюдения за участниками. В процессе записи ЭЭГ индуктору давали следующие установки: - расслабленного мышечного тонуса и рассеянного внимания (фон); - экстрасенсорной диагностики состояния реципиента; - биокоррекции состояния реципиента. Реципиенту рекомендовали принять удобное положение тела, закрыть глаза и поддерживать рассеянное внимание во время работы индуктора. Полученные данные ЭЭГ подвергали визуальной оценке, исключали артефакты и проводили обработку с последующим картированием полученных результатов. Основная цель обработки заключалась в сопоставлении корреляционных связей по отдельным частотам активности мозга, одновременно зарегистрированной у двух лиц. При этом под биоинформационной связью понимали наличие математически выраженной корреляционной зависимости между потенциалами мозга двух участников (с абсолютным значением коэффициента выше 0,6). Аналогичные подходы широко использовались в отечественных и зарубежных нейрофизиологических исследованиях для оценки связей между структурами головного мозга. Результаты исследований В процессе своей работы индуктор обычно испытывал ряд переходных состояний, позволяющих ему настроиться на реципиента, проявить способность экстрасенсорного восприятия с целью проведения биоинформационной диагностики и воздействия. В этих переходных состояниях сознания анализировали взаимные корреляционные отношения между колебаниями ЭЭГ индукторов и реципиентов в узких диапазонах частот (полосой до 1 Гц). Обычно в фоновой регистрации ЭЭГ наблюдались низкие значения коэффициентов взаимных корреляций (от +0,2 до +0,5) между биопотенциалами мозга индуктора и реципиента. В этом случае структура биоинформационных связей не рассматривалась. В процессе последующей настройки индуктора на реципиента на отдельных частотах наблюдались как положительные, так и отрицательные взаимные корреляционные связи с более высокими, по сравнению с фоном, абсолютными значениями коэффициентов взаимной корреляции (r<-0,6; r>+0,6), что могло быть связано с выраженной синхронизацией отдельных ритмов мозга индуктора и реципиента. При этом положительные и отрицательные значения коэффициентов корреляции отражали соответственно процессы синхронизации биоритмов в фазе и противофазе. Обычно настройка индуктора на реципиента начиналась с формирования противофазных отношений альфа-ритма, который, как известно, является ритмом, наиболее выраженным по амплитуде и мощности в состоянии спокойного бодрствования и сенсорной депривации. В этом диапазоне частот наблюдалась следующая структура биоинформационных связей между индуктором и реципиентом (см. рис.1). 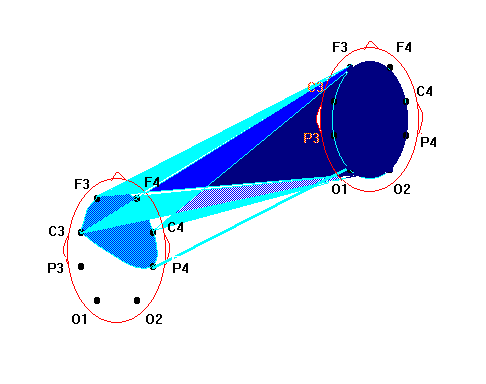 Рис.1. Энергоинформационные связи, определяемые по межиндивидуальной синхронизации биопотенциалов мозга, в диапазоне альфа-ритма в периоде настройки индуктора на реципиента. Коэффициент кросскорреляции [R]>-0,6. (Обозначения областей мозга и подробности описания см. в тексте). На примере, представленном на рис.2, видно, что в процессе диагностики между ЭЭГ двух участников возникали значимые корреляционные связи положительного знака в бета-диапазоне, отражая фазовые взаимоотношения передних отделов левого полушария одного партнера и центральных отделов другого. Эта структура биоинформационных связей сосуществовала одновременно с предыдущей. При этом отрицательная корреляционная зависимость в альфа-диапазоне, повидимому, отражала явление контрастности в механизме экстрасенсорного восприятия. 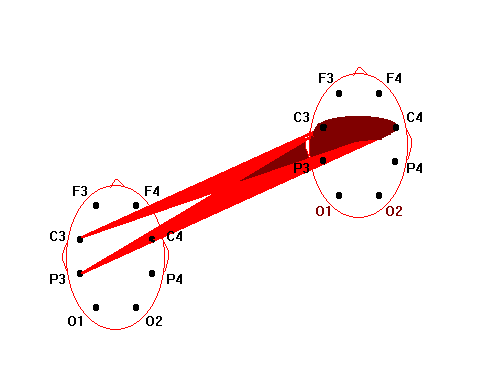 Рис.2. Пример энергоинформационных связей в диапазоне бета-ритма в периоде проведения индуктором диагностики состояния реципиента. Коэффициент кросскорреляции R>0,6. (Подробности см. в тексте). На рис.3 представлена структура биоинформационных связей в тета-диапазоне частотного спектра ЭЭГ, которая возникала в процессе проведения биокоррекции состояния реципиента. 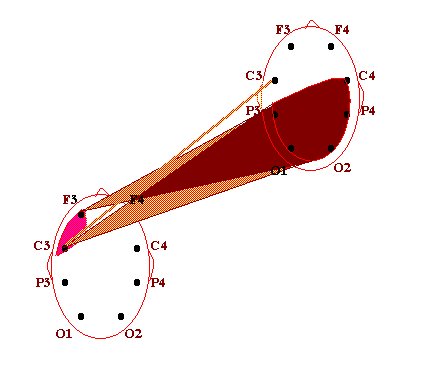 Рис.3. Пример энергоинформационных связей в диапазоне тета-ритма в периоде проведения индуктором коррекции состояния реципиента. Коэффициент кросскорреляции R>0,6. (Подробности см. в тексте). Таким образом, в процессе биоинформационного взаимодействия наблюдалась взаимосвязь между индуктором и реципиентом в виде усиления синхронизации (коэффициент корреляции 0,6-0,9) отдельных частот биоритмов мозга, по сравнению с фоном, где эти значения не превышали уровня 0,5. Для разных областей коры и разных частот корреляционная зависимость между активностью мозга индуктора и реципиента могла иметь одновременно противоположные по знаку значения и изменяться во времени, что свидетельствовало о сложной динамике процессов взаимодействия мозга двух индивидуумов в дискретно изменяющихся состояниях сознания. Чередование частот с положительными и отрицательными коэффициентами корреляции как в одном временном отрезке, так и последовательно в разных отрезках в фазе и противофазе между участниками свидетельствовало о возможных механизмах контрастирования, усиления и подавления в процессе биоинформационного обмена. Необходимо отметить, что, когда индуктор работает с реципиентом, он подключает обычно все вышеописанные механизмы одновременно, проводя биокоррекцию и используя диагностику в качестве обратной связи для оценки биокоррекции. Образуется сложная динамическая система биоинформационных связей, в которой попеременно участвуют разные области мозга как индуктора, так и реципиента с присущими им спектрами частот. При этом биоэлектрическая активность мозга каждого из участников сохраняет его индивидуальные особенности, относительно независимые от активности мозга другого участника. Таким образом, можно рассматривать три относительно независимые системы мозговых связей: индуктора и реципиента в отдельности и систему межиндивидуальных связей, которая названа виртуальным мозгом. Структура биоинформационных связей виртуального мозга отличается от структур функциональных связей как мозга индуктора, так и реципиента (см. рис.4). 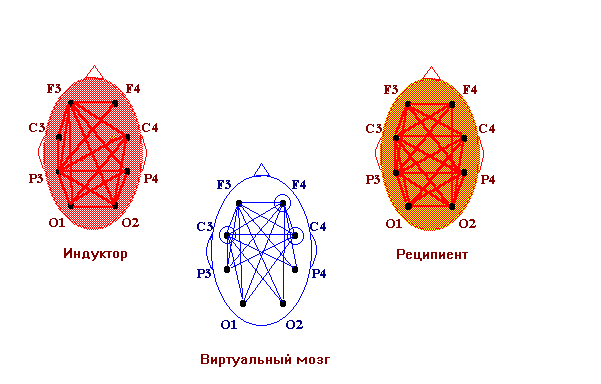 Рис.4. Функциональные связи, определяемые по показателям синхронизации биопотенциалов, между областями мозга у индуктора, у реципиента, и энергоинформационные связи виртуального мозга (пример одновременной регистрации). Наблюдаются различия структур связей, как у отдельных индивидуумов, так и отличия связей виртуального мозга от индивидуальных связей. Виртуальному мозгу принципиально присущи специфические свойства виртуальной реальности, описанные ранее [17]. Эти свойства следующие: - порожденность: виртуальный мозг продуцируется активностью другой реальности, в данном случае, активностью мозга индуктора и реципиента; - актуальность: виртуальный мозг существует актуально - только “здесь и теперь”, в данной виртуальной реальности свое время и пространство, свои законы существования; - интерактивность: виртуальный мозг взаимодействует с другими реальностями, а именно, с функциональными системами мозга индуктора и реципиента. Наряду с этим, проявляются свойства виртуального мозга, общие со свойствами индивидуальной активности мозга. Виртуальный мозг возникает благодаря взаимодействию тех же областей, функциональных центров, которые обеспечивают взаимодействие в индивидуальном мозге, и поэтому имеет сходные многопараметрические характеристики своей деятельности. Существует такая же динамичность биоинформационных связей между областями виртуального мозга, как и функциональных связей между областями индивидуального мозга индуктора или реципиента, однако, отличающаяся по своей структуре, так как каждая биоинформационная связь отражает взаимодействие между какой либо областью мозга индуктора, с одной стороны, и реципиента - с другой. Динамичность биоинформационных связей виртуального мозга проявляется не только в пространстве, но и во времени, что свидетельствует о фазовой и амплитудной зависимости между биоритмами мозга индуктора и реципиента в процессе его существования. Наблюдается зависимость биоэнергоинформационных связей от несущей частоты: в какой-либо узкой полосе частот (в нашем случае, с частотным шагом 1 Гц) возможен переход от одной частоты к другой ближайшего спектра. В виртуальном мозге можно рассматривать межполушарные отношения, возможно также применение алгоритмов для обнаружения глубоких структур виртуального мозга, участвующих в формировании биоинформационных связей. Кроме того, возможно рассмотрение не только биоинформационных связей, но и биоритмов виртуального мозга, поскольку каждая связь формируется на биоритме какой-либо определенной частоты. В отличие от индивидуального, виртуальный мозг образует структуры биоинформационных связей, относительно устойчивые во времени и развивающиеся по правилам взаимодействия между индуктором и реципиентом. При этом, взаимодействие между областями виртуального мозга отличается от взаимодействия между аналогичными областями как у индуктора, так и у реципиента, и зависит от успешности внесенсорного контакта. Таким образом, к исследованию деятельности виртуального мозга применима та методология, которая используется для изучения мозга человека в нейрофизиологии. Однако, необходимо учитывать, что виртуальный мозг - это результат взаимодействия двух лиц - индуктора и реципиента, и за этим стоят специфические особенности виртуальной реальности. Биоритмы виртуального мозга - это только те биоритмы, которые участвуют в образовании биоинформационных связей. Поэтому частотный спектр ритмов виртуального мозга безусловно отличается от спектра индуктора или реципиента, он более сужен. А структура биоинформационных связей является более жестким образованием, по-видимому, с меньшим числом степеней свободы при использовании ограниченных в этих условиях частотных каналов передачи информации. После некоторого рассмотрения свойств биоинформационных связей и особенностей их функционирования можно полагать, что виртуальный мозг - это система отношений между отдельными областями мозга индуктора и реципиента, которые в виртуальном мозге рассматриваются как его части. В целом, виртуальный мозг может рассматриваться как объект для изучения закономерностей развития биоинформационных взаимодействий. Литература 1. Парамонов Ю.В., Зайденман И.А., Мирза Д.Г., Пятницкий А.П. Методологические вопросы экспериментальных исследований по обнаружению слабых реакций центральной нервной системы на внешний раздражитель. В сб. “Проблемы обнаружения слабых реакций нервной системы”, М., АН СССР, 1968, с.8-23 2. Свидерская Н.Е., Королькова Т.А., Селицкий Г.В. Влияние уровня сознания на пространственную организацию корковой активности при психических процессах. Журн. высш. нерв. деят., 1990, т.40, вып.5, с.934-944 3. Добронравова И.С., Лебедева Н.Н. Особенности межполушарных отношений когерентных характеристик ЭЭГ реципиентов и сенситивов при аутогенной деятельности. Парапсихология и психофизика. 1994, N1(13), с.50-59 4. Кукес В.Г., Коёкина О.И., Волков Р.Ю. Межиндивидуальная взаимосвязь биоэлектрической активности мозга при явлениях биоэнергоинформационного обмена. Парапсихология и психофизика. 1994, N1(13), с.42-50 5. Васильева Г.Н., Лытаев С.А., Хлуновский А.Л., Кобрин В.П. Исследование энергоинформационного обмена между субъектами методом электроэнцефалографии. Приборостроение, 1993, т.36, N6, с.65-72 6. Павлова Л.П. Системный подход в электроэнцефалографическом исследовании биотелесвязи в физиологической школе Н.Е.Введенского - А.А.Ухтомского. Парапсихология и психофизика. 1993, N2. С.42-48 7. Н.И.Заикин Н.И., Кривенков С.Г., Корсакова Е.А. Результаты электроэнцефалографических и психофизиологических исследований испытуемых, работающих в режиме ретрокогниции. Парапсихология и психофизика. 1994, №4, с.51-63 8. Благосклонова Н.К., Гусев А.Н., Коптелов Ю.М., Шапкин С.А. Отражение в ЭЭГ экстрасенсорного воздействия. Физиология человека. 1994, Т.20, N3, с.36-44 9. Ливанов М.Н. Пространственная организация процессов головного мозга. М., “Наука”, 1972г. 10. Костандов Э.А., Арзуманов Ю.Л. Физиологические критерии осознаваемости стимула. Журн. высш. нерв. деят., 1990 г., т.40, вып.6, с.1063-1073 11. Иваницкий А.М. Мозговая основа субъективных переживаний: гипотеза информационного синтеза. Журн. высш. нерв. деят., 1996, т.46, вып.2, с.241-252 12. Лебедева Н.Н., Добронравова И.С. Организация ритмов ЭЭГ человека при особых состояниях сознания. Журн. высш. нерв. деят., 1990 г., т.40, вып.5, с.951-962 13. Доброхотова Т.А., Брагина Н.Н., Федорук А.Г. Индивидуальный профиль функциональных асимметрий человека и парапсихология. Парапсихология и психофизика. 1993, N2, с.60-73 14. Ли А.Г., Иванова Т.К. Разработка способов синхронизации усилий в группах операторов для реализации явления телекинеза. Парапсихология и психофизика. 1995, №1, с.22-46 15. Аладжалова Н.А. Психофизиологические аспекты сверхмедленной ритмической активности головного мозга. М., “Наука”, 1979, с.214 16. Коекина О.И., Волков Р.Ю. Нейрофизиологические подходы в исследовании биоэнергоинформационного взаимодействия в процессе целительства. В сб. “Актуальные вопросы традиционной медицины и фармакотерапии”. М., 1995г., с.36-43 17. Носов Н.А. Психологические виртуальные реальности. М., 1994г., с.195 |