Опорно-двигательная система коллоквиум. коллок 4 опорно-двигательная. Опорнодвигательная система клетки
 Скачать 4.67 Mb. Скачать 4.67 Mb.
|

|
Коллоквиум № 4 "Опорно-двигательная система клетки". Основные вопросы: 1.Дать сравнительную характеристику молекулярно-структурной организации промежуточных филаментов, микрофиламентов и микротрубочек. Назвать основные белки и параметры их агрегации. Промежуточные филаменты(тонофибриллы) строятся из фибриллярных мономерных белков. Они имеют толщину 8-10 нм, пронизывают всю клетку, образуя систему наподобие каркаса, функционально связанную с системой микротрубочек (см. ниже); при разрушении системы микротрубочек распадается и система промежуточных филаментов. Основное положение по периферии клетки или в середине. Тканеспецифичны: Кератины – характерны для эпителиальных клеток, найдено около 30 форм. Выработка кератинов ведѐт к кератинизации (ороговению) клеток эпителия и, в итоге, превращению их в роговые чешуи. Кератинизация эпителиальных клеток усиливает барьерно-защитную функцию кожных эпителиев высших позвоночных. В зависимости от степени ороговения (от интенсивности синтеза кератинов в клетках) выделяют мягкую и жесткую кератинизацию. Так, эпидермис кожи человека – пример мягкой кератинизации, а волосы и ногти – пример жѐсткой кератинизации. Виментин – входит в состав цитоскелета клеток мезенхимного происхождения, например, всех клеток соединительной ткани и крови. Десмин – присутствует в мышечных клетках. Глиальный фибриллярный белок – входит в состав цитоскелета клеток нейроглии,в астроцитах. Периферин – входит в состав цитоскелета периферических и центральных нейронов,держит форму. Белки нейрофиламентов – встречаются в аксонах нервных клеток. Белки ядерной ламины – разновидность белков ядерного матрикса, формирующих ламину. Промежуточные филаменты способны образовывать полимеры, состоящие из нескольких разновидностей белков), например, комплексы виментина с десмином или виментина с глиальными белками. Белки промежуточных филаментов не обладают свойством сократимости, поэтому данные структуры являются чисто опорными, скелетными. Микрофиламенты представляют собой сложные надмолекулярные комплексы, в состав которых входят как опорные, так и сократительные белки. Это позволяет микрофиламентам выполнять опорно-двигательную функцию и обеспечивать механизм мышечного сокращения. Микрофиламенты входят в состав миофибрилл мышечных клеток, а также образуют правильную гексагональную сеть в цитоплазме немышечных клеток – так называемые стресс-фибриллы, сокращение которых приводит к амебоидному движению клеток. Есть у всех клеток,включая растительные. Разберем молекулярную структуру микрофиламента. Основной белок системы микрофиламентов – актин. В мономерной форме молекула актина представляет собой глобулярный белок (G-актин). Фибриллярная (нитчатая) форма актина (F-актин) образуется путем самосборки молекул глобулярного актина в спиральную ленту толщиной 6-7 нм . Самосборка (полимеризация) начинается при достаточной концентрации молекул актина. Также ингибитор полимеризации актина -белок профилин. При этом актиновая фибрилла обладает полярностью в отношении процесса полимеризации – в ней можно выделить «+»-конец и «-»-конец. На «+»-конце полимеризация преобладает над деполимеризацией, а на «-» конце, наоборот, деполимеризация преобладает над полимеризацией . Таким образом, актиновый филамент растет со стороны «+»-конца, а со стороны «-»-конца убывает. Полимеризация актина носит обратимый характер. Благодаря обратимым изменениям концентрации G- и F-форм меняется и вязкость цитоплазмы: преобладание G-актина соответствует более жидкой ее консистенции (золь),в которой легче двигаются органоиды, а преобладание F-актина – более вязкой (гель). Кроме того, в клетке нити актина стабилизируются массой специфических белков, ассоциированных с F-актином. Так, тропомиозин придаѐт микрофиламентам жѐсткость, фимбрин связывает филаменты в пучки, филамин и альфа-актинин образуют поперечные скрепки между микрофиламентами, создавая сложную трѐхмерную сеть, которая и придает цитоплазме гелеобразное состояние (рис. 69). Как уже упоминалось, микрофиламенты пронизывают всю клетку, образуя особенно густую сеть в кортикальном слое цитоплазмы (под плазматической мембраной) – кортикальный цитоскелет. Эта сеть стабилизирует форму клетки. Один из механизмов движения псевдоподий амебоидной клетки заключается в том, что в месте выпячивания псевдоподий сеть микрофиламентов разбирается, цитоплазма разжижается и начинает натекать на плазматическую мембрану, после чего этот «натѐк» стабилизируется восстановлением кортикальной сети микрофиламентов. Надстраивающаяся сеть микрофиламентов может также прогибать плазматическую мембрану, образуя еѐ выпячивания – тонкие филоподии и более широкие ламеллоподии. Клетка крепится к субстрату при помощи специальных клеточных контактов – фокальных контактов. Самый главный сопутствующий актиновым филаментам белок – миозин. Если связь с другими белками позволяла актину выполнять преимущественно функцию скелетного белка, то в ассоциации с миозином актин участвует в движении. Молекула миозина состоит из двух головок (тяжелый меромиозин) и двух переплетѐнных хвостиков (легкий меромиозин) (рис. 72, а). Особенностью миозина является его способность расщеплять АТФ и за счет освободившейся энергии обратимо менять свою конформацию (третичную структуру); при этом головка миозина меняет угол наклона по отношению к хвостику. Взаимодействуя с актиновыми микрофибриллами, миозин участвует и в амебоидном движении клетки, и в движении органоидов клетки, и в колебании микроворсинок, и в сокращении миофибрилл мышечных клеток. НА КАЖДЫЙ ШАГ(10-20НМ) НУЖНА МОЛЕКУЛА АТФ. В гладких мышцах сократительная сеть построена на основе стрессс-фибрил,которые могут долгое время быть в сокращенном состоянии. Z-полоска это альфа актинин с десмином. Темный диск никогда не меняет свой размер. 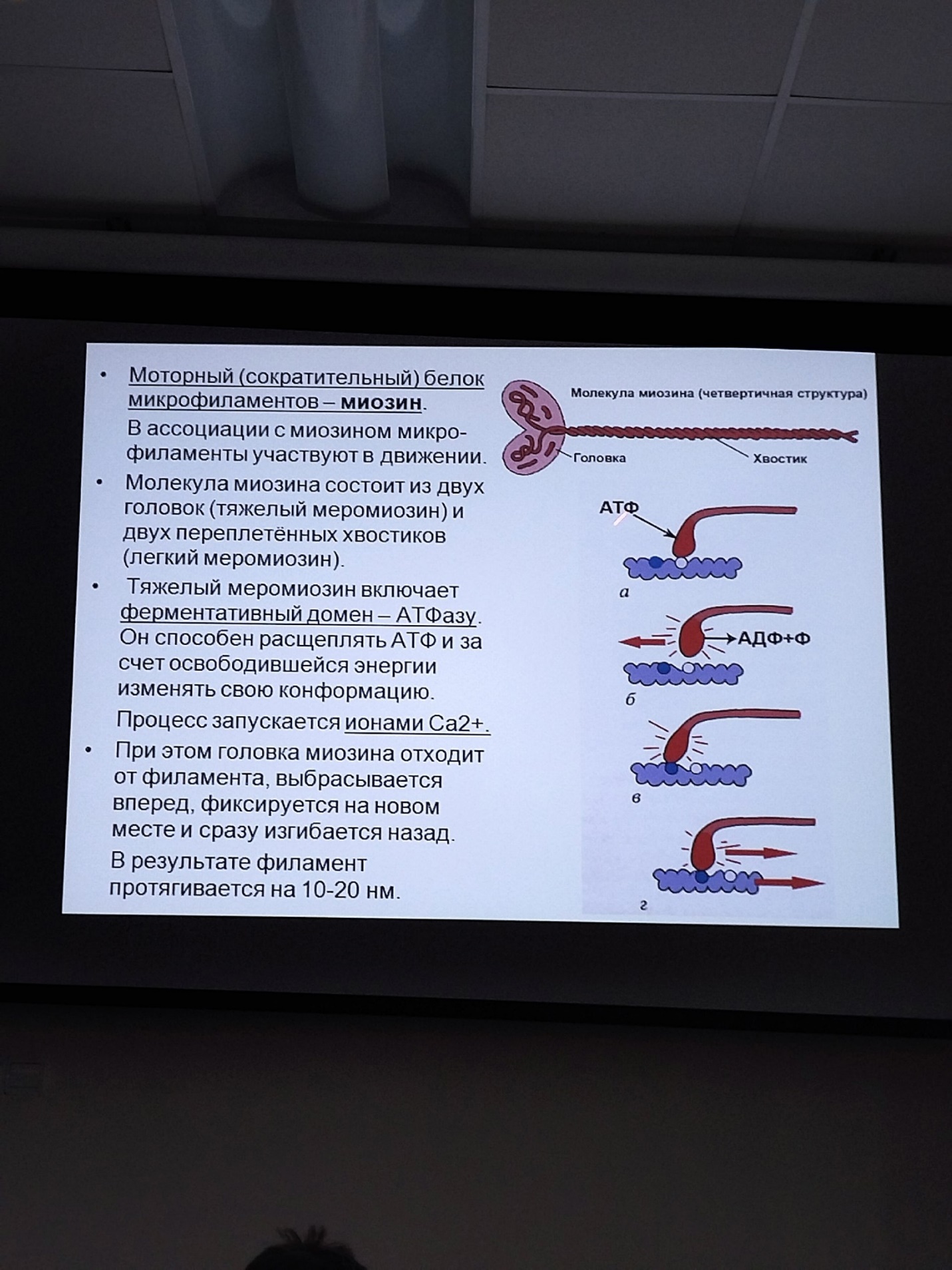 процесс запускается ионами кальция, они «закупоривает» процесс запускается ионами кальция, они «закупоривает»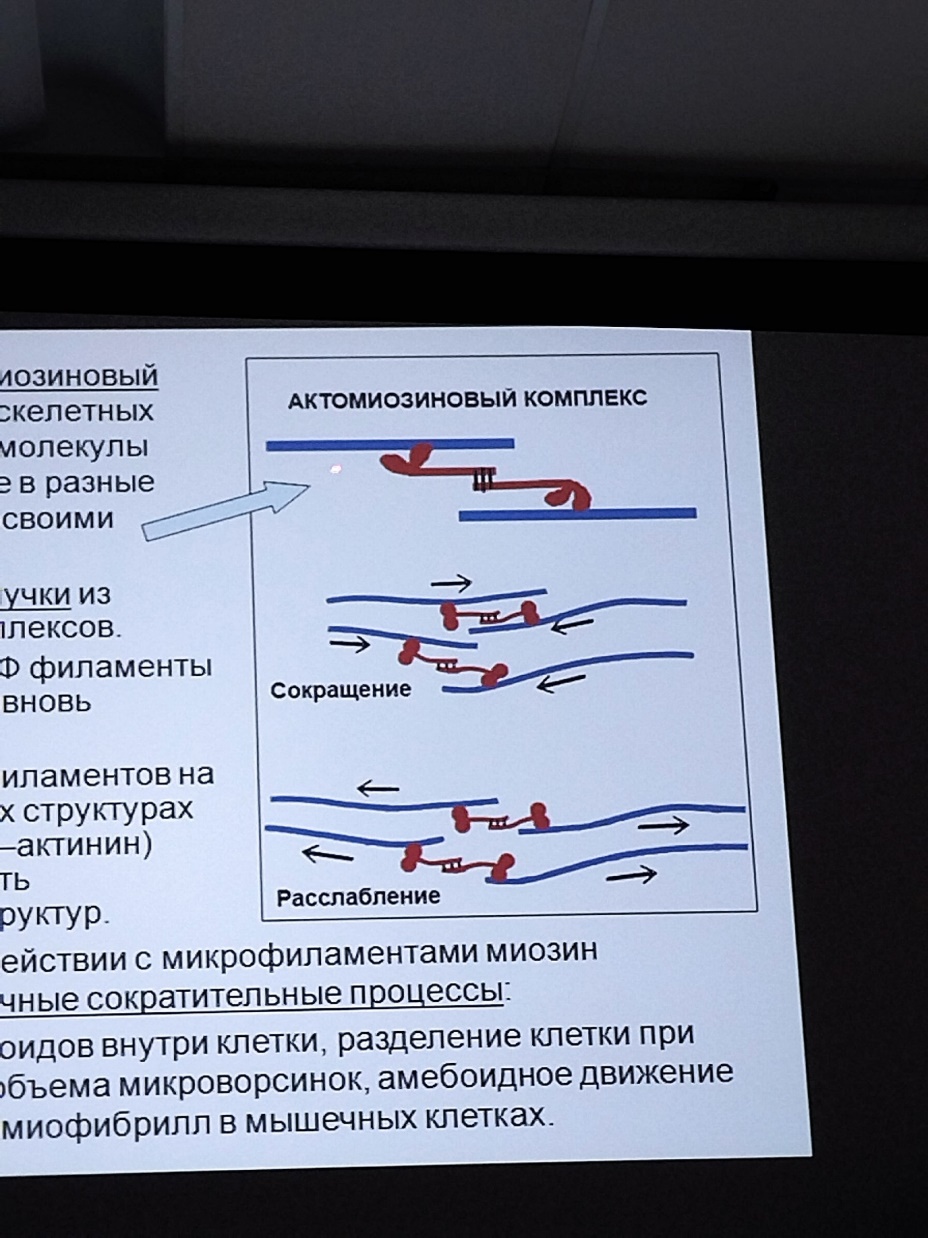 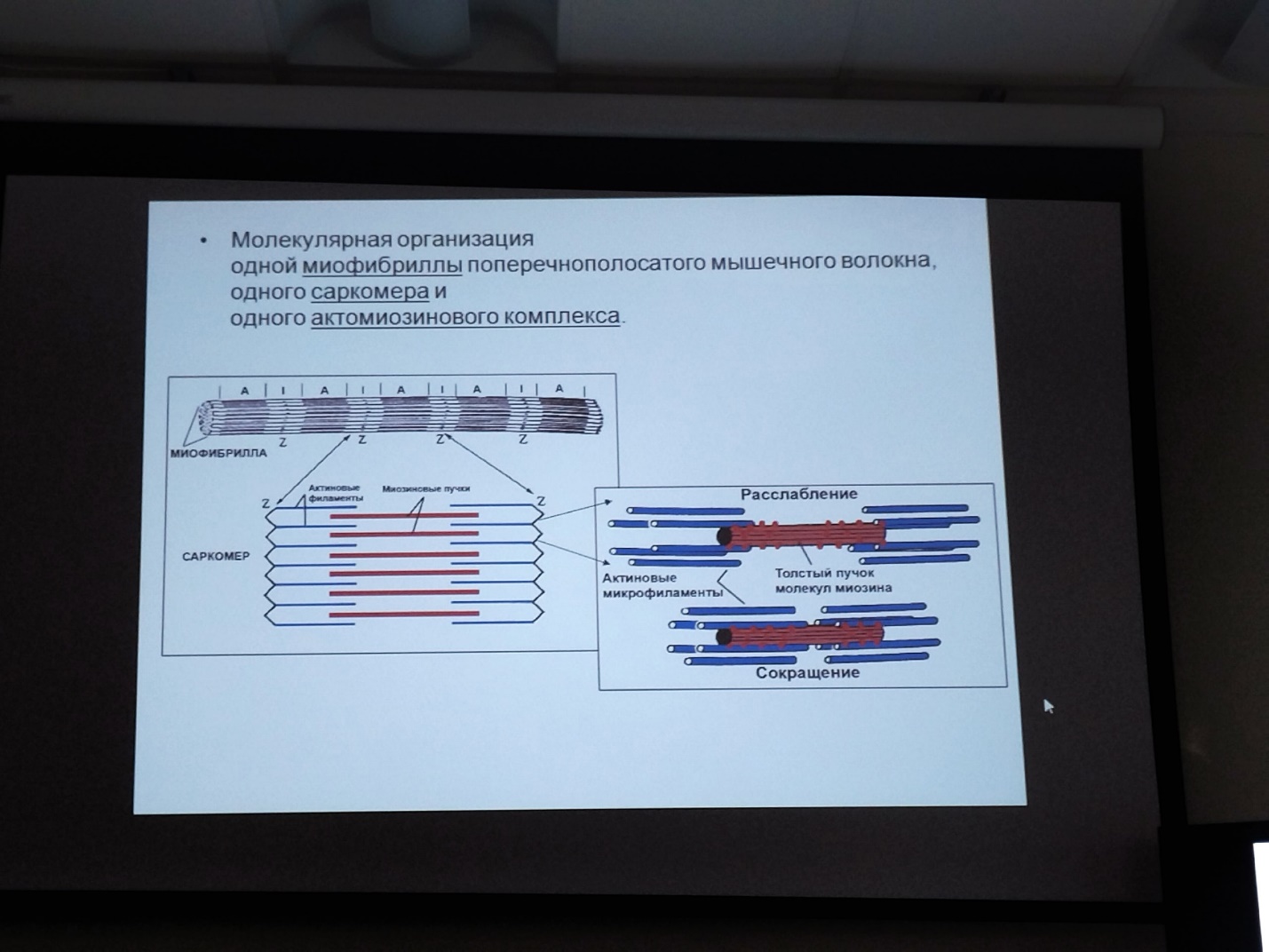 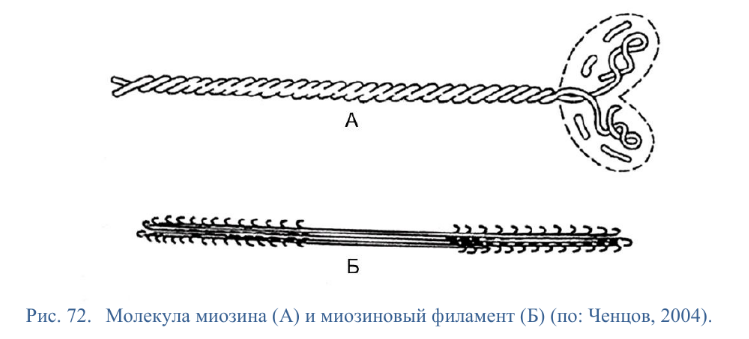 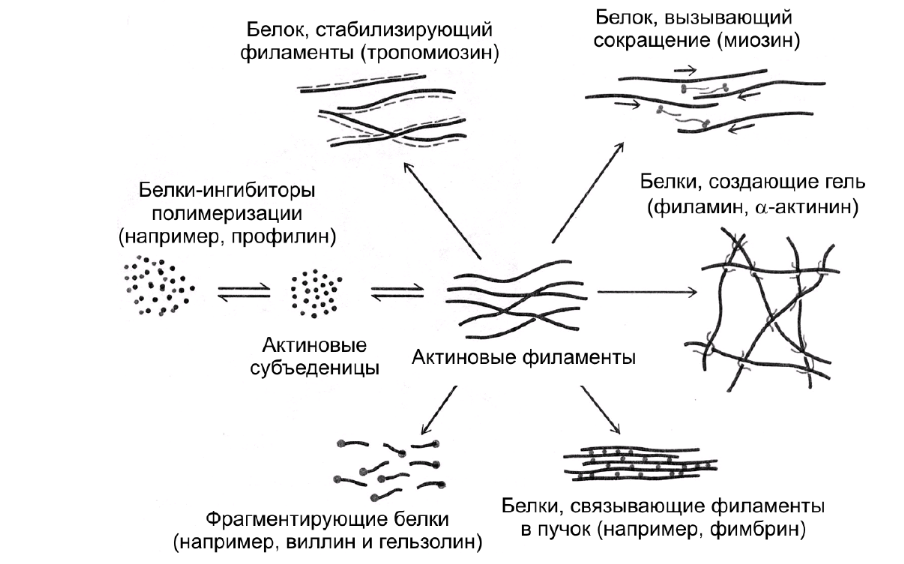 + тропонин Микротрубочки представляют собой полые цилиндры диаметром 22 нм, построенные из глобулярного белка тубулина. Присутствуют у всех эукариотных ктлеток. Существует две разновидности тубулина – альфа- и бета- тубулин. Есть гама-тубулин(учавствует только в сборке микротрубочек центриолей). Одна глобула альфа-тубулина и одна глобула бета-тубулина объединяются в димер, а димеры, в свою очередь, полимеризуются с образованием микротрубочки (рис. 77). 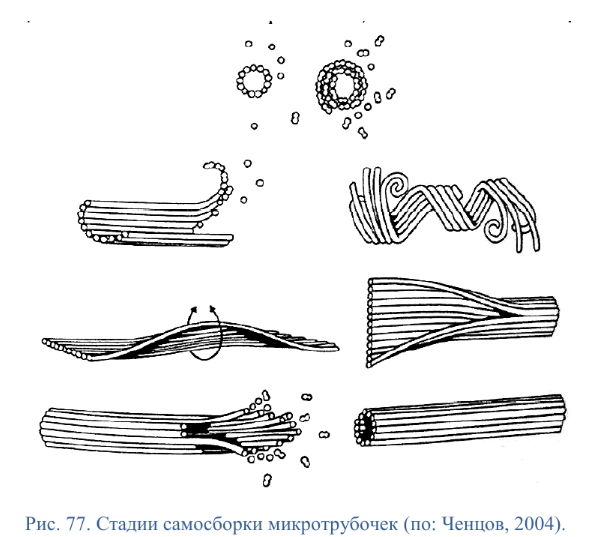 Микротрубочки, как и актиновые микрофиламенты, представляют собой самособирающиеся, динамичные, полярные структуры. У микротрубочки имеется быстро растущий «+»-конец и медленно растущий или разбирающийся «-»-конец. В покоящейся клетке все микротрубочки подвергаются полной сборке-разборке каждые 15-20 минут, а в процессе деления клетки среднее время жизни микротрубочки – 30 секунд. В составе микротрубочек, помимо тубулина, обнаруживается ряд ассоциированных с ними белков – так называемые МАР-белки (microtubule associated proteins). Эти белки связываются либо с «+»-концами, либо с «-»-концами микротрубочек, блокируя, таким образом, процессы сборки или разборки соответственно. Полимеризация происходит при гидролизе ГТФ до ГДФ. Система микротрубочек (рис. 78), как и система микрофиламентов, выполняет в клетке скелетную (опорную) и двигательную функции. Опорная функция заключается в том, что микротрубочки стабилизируют форму клетки, образуя ее радиальный скелет – астросферу. Двигательная функция состоит в образовании комплексов микротрубочек со специальными сократительными белками, способными приводить в движение клеточные компоненты. Основные двигательные белки, ассоциированные с системой микротрубочек, – это динеин и кинезин. Они состоят из двух тяжѐлых белковых цепей и нескольких лѐгких. Двигательные белки могут связываться с одной стороны с мембранами органоидов или другими внутриклеточными структурами, а с другой стороны – с микротрубочкой. Изменяя свою конформацию при расщеплении АТФ, белок «шагает» по микротрубочке, перенося за собой связанный органоид (рис. 79). Кинезин перемещается по микротрубочке в направлении к «+»-концу, динеин – к «-»-концу. Таким образом, в клетке по системе микротрубочек, как по рельсам, осуществляется массовый перенос различных внутриклеточных компонентов – органоидов, вакуолей, хромосом и др. В клетке существует множество специфических модификаций двигательных белков, участвующих в переносе строго определѐнных типов структур. Движение жгутиков и ресничек в эукариотической клетки также основано на функционировании системы микротрубочек при ее взаимодействии с двигательными белками, а именно, с динеином. Ресничка (или жгутик) представляет собой вырост плазматической мембраны, центральный скелет которого – аксонема – построен из микротрубочек, собранных в дуплеты. Схема расположения микротрубочек в аксонеме показана на рис. 80 В центре этого образования расположен центральный дуплет, а вокруг, по периметру реснички, располагаются ещѐ 9 дуплетов микротрубочек. Эти дуплеты соединены друг с другом так называемыми динеиновыми ручками (рис. 80). В момент движения реснички молекулы динеина меняют свою конформацию таким образом, что ручки вытягиваются, захватывают микротрубочку из соседнего дуплета и «бегут» по ней. В результате происходит «скольжение» дуплетов микротрубочек друг относительно друга, и ресничка изгибается. Следует добавить, что в основании реснички лежит базальное тельце, организация которого принципиально сходна с организацией самой реснички, только вместо дуплетов микротрубочек базальное тельце содержит их триплеты, а центральный дуплет совсем отсутствует. Таким образом, ресничка как бы является продолжением базального тельца, но с небольшими изменениями в организации системы микротрубочек.   2.Для каких цитоскелетных структур характерны процессы самосборки и разборки: Что известно об этих механизмах? Микрофиламенты: Самосборка (полимеризация) начинается при достаточной концентрации молекул актина. При этом актиновая фибрилла обладает полярностью в отношении процесса полимеризации – в ней можно выделить «+»-конец и «-»-конец. На «+»-конце полимеризация преобладает над деполимеризацией, а на «-» конце, наоборот, деполимеризация преобладает над полимеризацией (рис. 68). Таким образом, актиновый филамент растет со стороны «+»-конца, а со стороны «-»-конца убывает. Микротрубочки, как и актиновые микрофиламенты, представляют собой самособирающиеся, динамичные, полярные структуры. У микротрубочки имеется быстро растущий «+»-конец и медленно растущий или разбирающийся «-»-конец. В покоящейся клетке все микротрубочки подвергаются полной сборке-разборке каждые 15-20 минут, а в процессе деления клетки среднее время жизни микротрубочки – 30 секунд. В составе микротрубочек, помимо тубулина, обнаруживается ряд ассоциированных с ними белков – так называемые МАР-белки (microtubule associated proteins). Эти белки связываются либо с «+»-концами, либо с «-»-концами микротрубочек, блокируя, таким образом, процессы сборки или разборки соответственно. 3.Микрофиламенты как скелетные и двигательные структуры. При каких условиях возникает способность микрофиламентов к двигательной реакции? Объясните механохимические основы мышечного движения. Один из механизмов движения псевдоподий амебоидной клетки заключается в том, что в месте выпячивания псевдоподий сеть микрофиламентов разбирается, цитоплазма разжижается и начинает натекать на плазматическую мембрану, после чего этот «натѐк» стабилизируется восстановлением кортикальной сети микрофиламентов. Надстраивающаяся сеть микрофиламентов может также прогибать плазматическую мембрану, образуя еѐ выпячивания – тонкие филоподии и более широкие ламеллоподии. Клетка крепится к субстрату при помощи специальных клеточных контактов – фокальных контактов. Прикрепляясь к субстрату, клетка генерирует механическое натяжение, – если поместить клетку на достаточно легко деформируемый субстрат, последний будет прогибаться с образованием складок. Механическое натяжение создается пучками актиновых филаментов – стресс-фибриллами, заякоренными с одной стороны на фокальные контакты, а с другой – на соседние стресс-фибриллы и, таким образом, пронизывающими весь объем клетки (рис. 70 и 71). 4.Дать сравнительную характеристику организации акто-миозиновых комплексов в немышечных (фибробласт, амеба) и специализированных мышечных клетках (поперечно-полосатое мышечное волокно). Показать иммуноцитохимическую локализацию альфа-актинина, актина и миозина в этих структурах. Разберѐм строение актин-миозинового комплекса в поперечно-полосатых (скелетных) мышцах. Скелетные мышцы позвоночных животных состоят не из отдельных клеток, а из мышечных волокон, каждое из которых представляет собой симпласт, образовавшийся за счѐт слияния многих мышечных клеток. Объединенная цитоплазма мышечного волокна содержит большое количество сократимых нитей – миофибрилл. В миофибриллах, в свою очередь, имеются тонкие нити – актиновые микрофиламенты, и толстые нити – миозиновые филаменты. Каждый миозиновый филамент состоит из двух разнонаправленных пучков миозиновых молекул, переплетенных своими хвостиками (рис. 72, б). Миозин имеет свойство обратимо связываться с помощью своих головок с актиновыми нитями и подтягивать их за счет изменения угла наклона головки, как было описано выше (рис. 73). Затем головка на короткий миг отцепляется от актина и делает «шажок»; при этом угол между головкой и хвостиком молекулы принимает исходную величину, но место прикрепления головки к актиновому филаменту меняется за счет совершившегося перемещения. Конечно, шаг миозиновой головки очень мал, и ей необходимо сделать тысячи «шажков», чтобы переместить актиновые нити на достаточную длину. 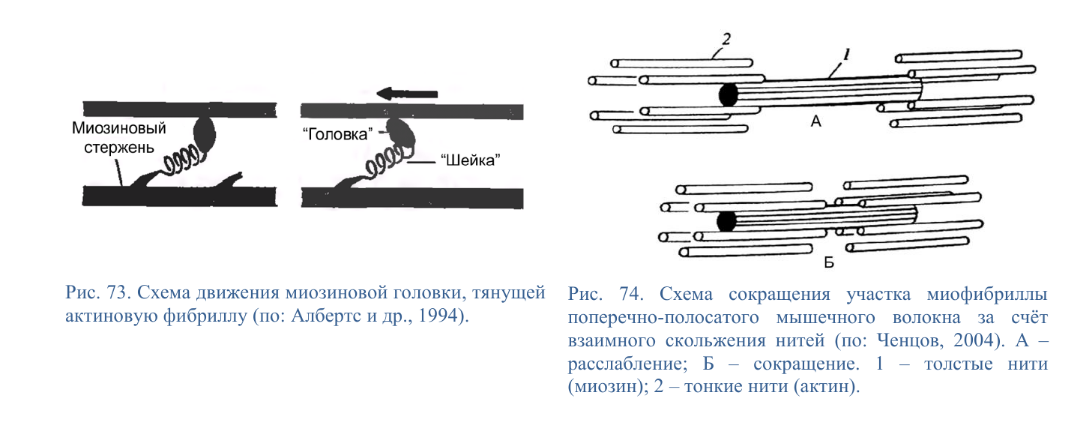 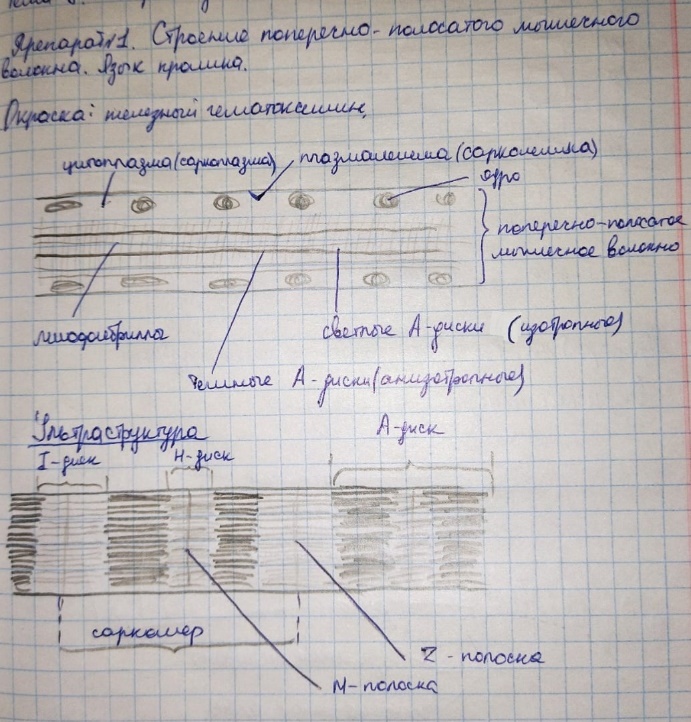 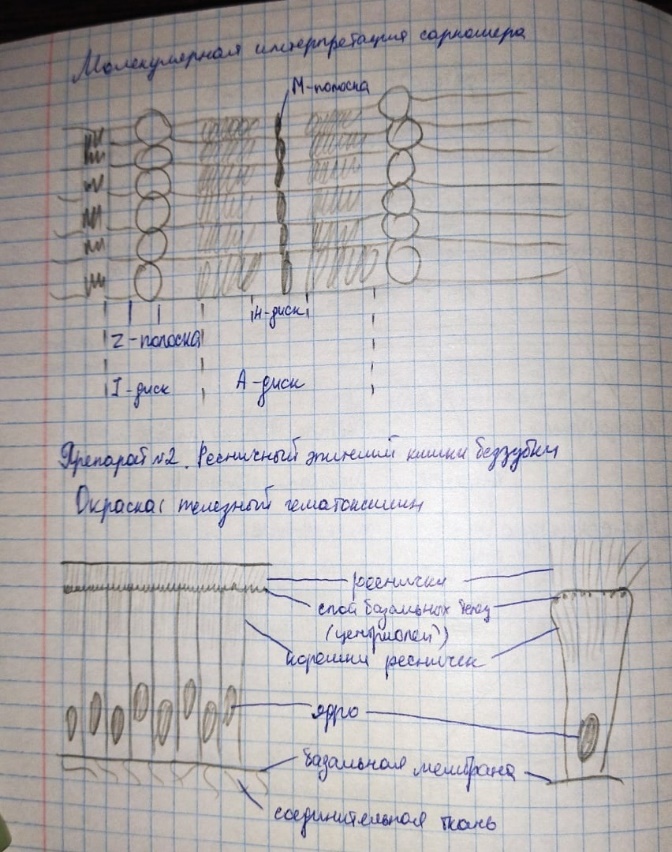 5.Микротрубочки как скелетные и двигательные структуры. Привести примеры использования микротрубочек в качестве цитоскелета. Система микротрубочек (рис. 78), как и система микрофиламентов, выполняет в клетке скелетную (опорную) и двигательную функции. Опорная функция заключается в том, что микротрубочки стабилизируют форму клетки, образуя ее радиальный скелет – астросферу. Двигательная функция состоит в образовании комплексов микротрубочек со специальными сократительными белками, способными приводить в движение клеточные компоненты. Основные двигательные белки, ассоциированные с системой микротрубочек, – это динеин и кинезин. Они состоят из двух тяжѐлых белковых цепей и нескольких лѐгких. Двигательные белки могут связываться с одной стороны с мембранами органоидов или другими внутриклеточными структурами, а с другой стороны – с микротрубочкой. Изменяя свою конформацию при расщеплении АТФ, белок «шагает» по микротрубочке, перенося за собой связанный органоид (рис. 79). Кинезин перемещается по микротрубочке в направлении к «+»-концу, динеин – к «-»-концу. Таким образом, в клетке по системе микротрубочек, как по рельсам, осуществляется массовый перенос различных внутриклеточных компонентов – органоидов, вакуолей, хромосом и др. В клетке существует множество специфических модификаций двигательных белков, участвующих в переносе строго определѐнных типов структур. 6.Показать строение реснички и объяснить механохимические основы ресничного движения. 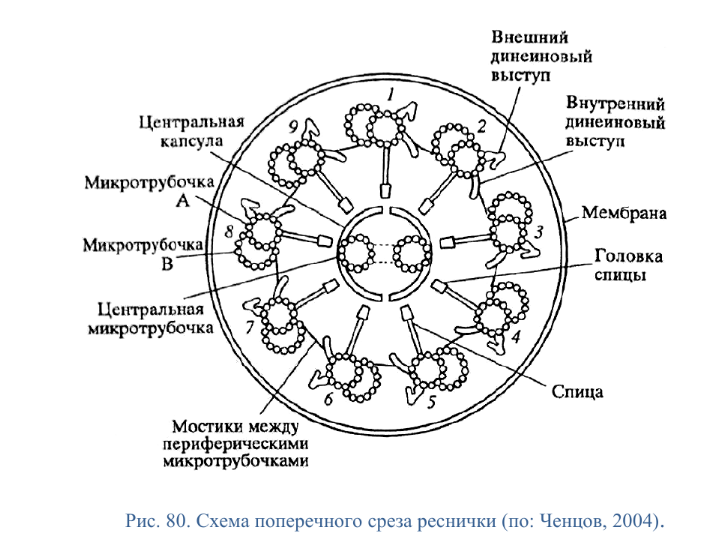 Движение жгутиков и ресничек в эукариотической клетки также основано на функционировании системы микротрубочек при ее взаимодействии с двигательными белками, а именно, с динеином. Ресничка (или жгутик) представляет собой вырост плазматической мембраны, центральный скелет которого – аксонема – построен из микротрубочек, собранных в дуплеты. Схема расположения микротрубочек в аксонеме показана на рис. 80 В центре этого образования расположен центральный дуплет, а вокруг, по периметру реснички, располагаются ещѐ 9 дуплетов микротрубочек. Эти дуплеты соединены друг с другом так называемыми динеиновыми ручками (рис. 80). В момент движения реснички молекулы динеина меняют свою конформацию таким образом, что ручки вытягиваются, захватывают микротрубочку из соседнего дуплета и «бегут» по ней. В результате происходит «скольжение» дуплетов микротрубочек друг относительно друга, и ресничка изгибается. Следует добавить, что в основании реснички лежит базальное тельце, организация которого принципиально сходна с организацией самой реснички, только вместо дуплетов микротрубочек базальное тельце содержит их триплеты, а центральный дуплет совсем отсутствует. Таким образом, ресничка как бы является продолжением базального тельца, но с небольшими изменениями в организации системы микротрубочек. 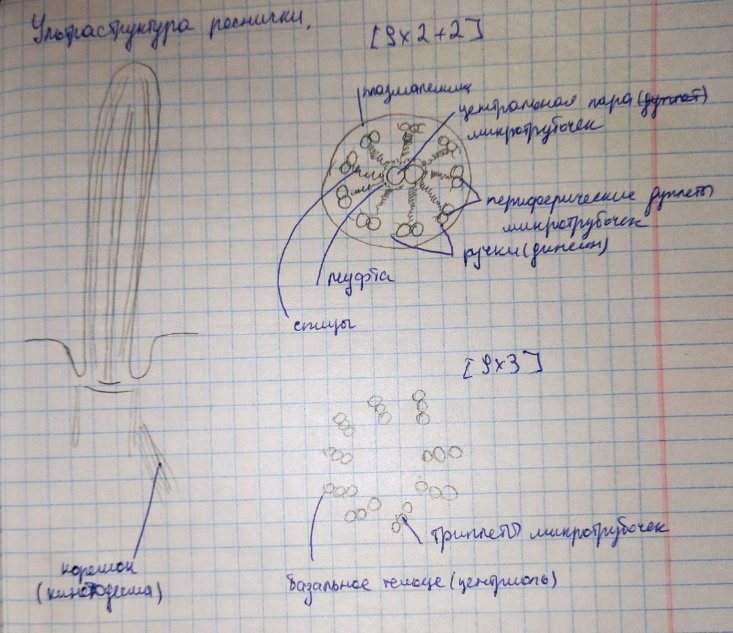 7.Что является центрами организации микротрубочек в клетке? Где они локализуются? Процесс полимеризации микротрубочек начинается в чётко ограниченных участках клетки, а именно – в центрах организации микротрубочек (ЦОМТ), ещё называемых клеточным центром. В основе ЦОМТ лежат две центриоли. Эта пара называется центросомой или диплосомой. Центриоли действительно чаще всего располагаются в геометрическом центре клетки, и от них во всех направлениях отходят микротрубочки, образующие радиальный скелет клетки и поддерживающие ее форму. В зонах ЦОМТ осуществляется закладка микротрубочек, обращѐнных к клеточному центру «-»-концами. На центриолях «-»-концы микротрубочек заблокированы специальными белками, предотвращающими их разборку. Соответственно, «+»-концы микротрубочек обращены в цитоплазму и растут по направлению к периферии клетки. 8.Строение центриолей, их взаиморасположение в клеточном центре. Механизм воспроизведения центриолей. 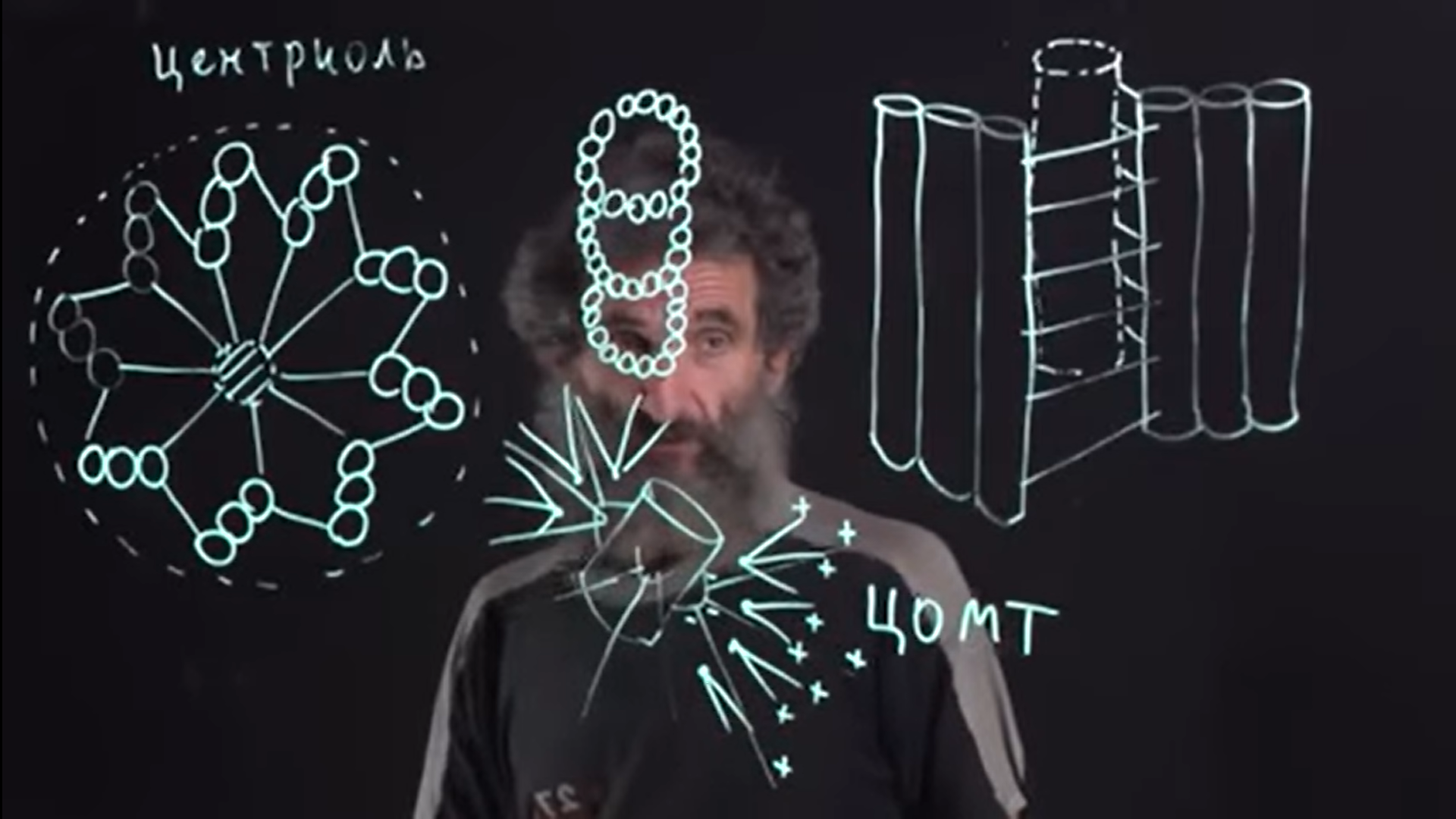 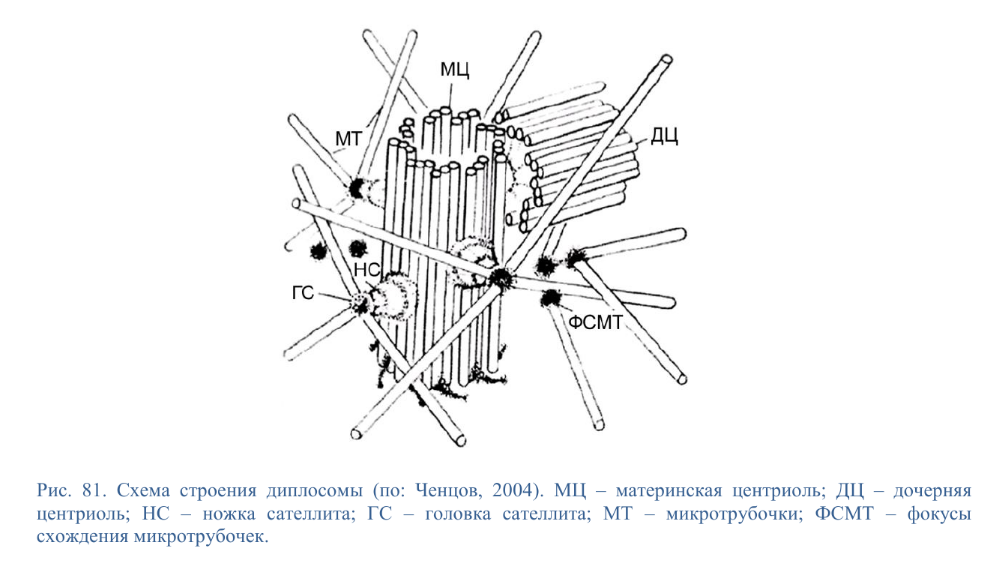 Диплосома состоит из двух центриолей – материнской и дочерней. Каждая центриоль представляет собой полый цилиндр, образованный девятью триплетами микротрубочек. При делении клетки центриоли расходятся к полюсам и передаются по одной штуке в дочерние клетки. Затем каждая из них достраивает новую дочернюю центриоль. В качестве ЦОМТ в клетке функционирует лишь материнская центриоль, на которой располагаются особые образования – сателлиты, от которых и отходят микротрубочки (рис. 81). Когда центриоли расходятся к полюсам делящейся клетки, бывшая дочерняя центриоль подвергается перестройке и приобретает сателлиты, – теперь она тоже может выступать в качестве клеточного центра. Микротрубочки, отрастающие теперь от обеих центриолей, направлены от полюсов к экватору и играют роль веретена деления. По микротрубочкам веретена с помощью особой формы динеина будут перемещаться хромосомы – от центра к полюсам клетки. 9.Какова роль центриолей и как объяснить их отсутствие в клетках высших растений? У центриолей в клетке имеется еще одна важная функция – именно они выступают в качестве базальных телец ресничек и жгутиков (см. выше). Следует заметить, что центриоли отсутствуют в клетках голосеменных и цветковых растений. Это связано с тем, что у их клеток жгутики или реснички не обнаруживаются ни на одной стадии жизненного цикла, в отличие от всех остальных высших растений, включая мхи и папоротники. ЦОМТ можно посадить на мембрану. Дополнительные вопросы: 10.Провести аналогию в механохимической организации и принципах работы актин- миозиновых и тубулин-динеиновых двигательных систем. 11.Какие опорные и двигательные структуры имеются у растительных клеток? Как они развиты по сравнению с животными клетками? 12.Как организован двигательный аппарат бактерий? Каковы принципиальные отличия бактериального жгутика от реснички эукариотной клетки?  |