ФИЗИОЛОГИЯ. Основные физиологические свойства живых тканей. А возбудимость
 Скачать 440.61 Kb. Скачать 440.61 Kb.
|

А)Возбудимость- способность живых клеток воспринимать изменения внешней среды и отвечать на эти изменения (раздражения) реакцией возбуждения. Б)Рефрактерность- невозбудимость во время возбуждения В) Лабильность- максимальный ритм раздражений, который живая ткань может воспроизвести за еденицу времени. Г)Сократимость- Способность развивать силу и напряжение, изменяя конфигурацию, при переходе в состояние возбуждения. Д) проводимость- Способность клеток органов и тканей проводить волну возбуждения.
Все мембраны по своей организации и составу обнаруживают ряд общих свойств. Они:
Основу мембраны составляет липидный бислой – двойной слой молекул липидов, которые обладают свойством амфифильности (содержат как гидрофильные, так и гидрофобные функциональные группы). В липидном бислое гидрофобные участки молекул взаимодействуют между собой, а гидрофильные участки обращены в окружающую водную среду.
Мембранным потенциалом покоя (МПП) или потенциалом покоя (ПП) называют разность потенци алов покоящейся клетки между внутренней и наружной сторонами мембраны. Основные положения мембранной теории происхождения МПП сводятся к следующему. В состоянии покоя клеточная мембрана хорошо проницаема для ионов К+ (в ряде клеток и для СГ), менее проницаема для Na+ и практически непроницаема для внутриклеточ ных белков и других органических ионов. Ионы К+ диффундируют из клетки по концентрационному градиенту, а непроникающие анионы остаются в цитоплазме, обеспечивая появление разности по тенциалов через мембрану. Возникающая разность потенциалов препятствует выходу К+ из клет ки и при некотором ее значении наступает равновесие между выходом К+ по концентрационному градиенту и входом этих катионов по воз никшему электрическому градиенту. Мембранный потенциал, при ко тором достигается это равновесие, называется равновесным потенци алом.
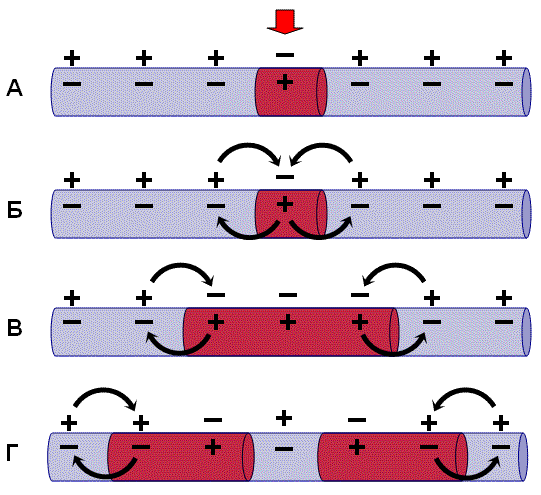
Потенциал действия – это сдвиг мембранного потенциала, возникающий в ткани при действии порогового и сверхпорогового раздражителя, что сопровождается перезарядкой клеточной мембраны. При действии порогового или сверхпорогового раздражителя изменяется проницаемость клеточной мембраны для ионов в различной степени. Для ионов Na она повышается и градиент развивается медленно. В результате движение ионов Na происходит внутрь клетки, ионы К двигаются из клетки, что приводит к перезарядке клеточной мембраны. Наружная поверхность мембраны несет отрицательный заряд, внутренняя – положительный. Компоненты потенциала действия: 1) локальный ответ; 2) высоковольтный пиковый потенциал (спайк); 3) следовые колебания. Ионы Na путем простой диффузии поступают в клетку без затрат энергии. Достигнув пороговой силы, мембранный потенциал снижается до критического уровня деполяризации (примерно 50 мВ). Критический уровень деполяризации – это то количество милливольт, на которое должен снизиться мембранный потенциал, чтобы возник лавинообразный ход ионов Na в клетку. Высоковольтный пиковый потенциал (спайк). Пик потенциала действия является постоянным компонентом потенциала действия. Он состоит из двух фаз: 1) восходящей части – фазы деполяризации; 2) нисходящей части – фазы реполяризации. Быстрое и большое количество поступление ионов Na в клетку приводит к изменению потенциала на клеточной мембране. Чем больше ионов Na войдет в клетку, тем в большей степени деполяризуется мембрана, тем больше откроется активационных ворот. Возникновение заряда с противоположным знаком называется инверсией потенциала мембраны. Движение ионов Na внутрь клетки продолжается до момента электрохимического равновесия по иону Na Амплитуда потенциала действия не зависит от силы раздражителя, она зависит от концентрации ионов Na и от степени проницаемости мембраны к ионам Na. Нисходящая фаза (фаза реполяризации) возвращает заряд мембраны к исходному знаку. При достижении электрохимического равновесия по ионам Na происходит инактивация активационных ворот, снижается проницаемость к ионам Na и возрастает проницаемость к ионам K. Полного восстановления мембранного потенциала не происходит. Потенциал действия (ПД)-быстрое колебание МП-самораспространяющийс процесс,связанный с изменениями ионной проводимости мембраны,вызванным функционированием ионных каналов. ПД распространяется без затухания,т е практически без уменьшения амплитудыМежду моментом нанесения раздражения и первым проявлением ПД имеется задержка-латентный период(ЛП). ЛП соответствует времени ,когда ПД движется по мембране нервной клеткиот места раздражения до отводящего электрода. Под действием раздражающего стимула происходит нарастающая деполяризация мембраны-локальный ответ.При достижении критического уровня деполяризации,сост в средн-55 мВ,начинается фаза деполяризации.В эту фазу уровень МП падает до 0 и даже преобретает положительное значение (овершут),а затем возвращается к исходному уровню фаза реполяризации.Фазы деполяризации,овершута и реполяризации образуют спайк (пик)ПД,длительность которого(1-2 милисек).После спайка наблюдается замедление скорости спада потенциала-фаза следовой деполяризации.После достижения исходного уровня покоя нередко наблюдается фаза следовой гиперполяризации.Они могут длиться десятки,сотни миллисек.В основе изменений МП,происходящий в течении ПД,лежат ионные механизмы.Локальный потенциал.раздражение клетки приводит к открытию части Na+каналов и появлению локального (нераспространяющегося потенциала)Фаза деполяризации.При достижении критического уровня деполяризации мембраны начинается лавинообразный процесс открытия большлго количества Na+-каналов.В фазу деполяризац происходит массивный вход в клетку ионов Na+по концетрационному и электрохимическому градиенту.Овершут.Деполяризация мембраны приводит к реверсии МП(МП станов положит).В эту фазу Na+-ток начинает стремительно спадать,что связано с инактивацией потенциало-зависимых Na+-каналов(время открытого состояния-доли сек) и исчезновением электрохимического градиента Na+.Фаза реполяризации.Инактивация Na+-каналов открытие потенциалозависимых К+-каналов способствуют развитию реполяризации.Выход К+ наружу способствует полноценному завершению фазы реполяризац.Следовые потенциалы связаны с длительными изменениями кинетических свойств К+-каналов.Восстановление исходного уровня Мп приводит Na- и К-каналы в состояние покоя.В ходе развития ПД происходят изменения возбудимости мембраны нервной клетки.Абсолютно рефрактерный период. Во время фазы деполяризации и большей части фазы реполяризации ПД клетка находится в абсолютно рефрактерном периоде, в течении которого даже сверхпороговое раздражение не способно вызвать пД.Это связано с инактивацией большинства Na-кан.Относительно рефрактерный период.В конце фазы реполяризации,а так же во время следовой гипераполяризации клетка способна генерировать ПД только в ответ на сверхпороговые раздражители.Это связано со значительным реполяризующим действием выходящих Ка токовЛабильность-максимально возможная частота генерации ПД для данного типа возбудимой клетки.Лабильность большинства нейронов составляет приблиз-но 400 ПД/сСоотношение фаз возбудимости с фазами ПДУровень возбудимости клетки зависит от фазы ПД. В фазу локального ответа возбудимость возрастает. Это фазу возбудимости называют латентным дополнением.В фазу реполяризации ПД, когда открываются все натриевые каналы и ионы натрия лавинообразно устремляются в клетку, никакой даже сверхсильный раздражитель не может стимулировать этот процесс. Поэтому фазе деполяризации соответствует фаза полной невозбудимости или абсолютной рефрактерности.В фазе реполяризации все большая часть натриевых каналов закрывается. Однако они могут вновь открываться при действии сверхпорогового раздражителя. Т.е. возбудимость начинает вновь повышаться. Этому соответствует фаза относительной невозбудимости или относительной рефрактерности.Во время следовой деполяризации МП находится у критического уровня, поэтому даже допороговые стимулы могут вызвать возбуждение клетки. Следовательно в этот момент ее возбудимость повышена. Эта фаза называется фазой экзальтации или супернормальной возбудимости.В момент следовой гиперполяризации МП выше исходного уровня, т.е. дальше КУД и ее возбудимость снижена. Она находится в фазе субнормальной возбудимости. Следует отметить, что явление аккомодации также связано с изменением проводимости ионных каналов. Если деполяризующий ток нарастает медленно, то это приводит к частичной инактивации натриевых, и активации калиевых каналов. Поэтому развития ПД не происходит.Возбуждение может быть 2-х видов: местное (локальный ответ);распространяющееся (импульсное). Местное возбуждение- наиболее древний вид (низшие формы организмов и низковозбудимые ткани - например, соединительная ткань). Местное возбуждение возникает и в высокоорганизованных тканях под действием подпорогового раздражителя или как компонент потенциала действия. При местном возбуждении нет видимой ответной реакции. Особенности местного возбуждения: нет латентного (скрытого) периода - возникает сразу же при действии раздражителя; нет порога раздражения; местное возбуждение градуально - изменение заряда клеточной мембраны пропорционально силе подпорогового раздражителя; нет рефрактерного периода, наоборот характерно небольшое повышение возбудимости; распространяется с декрементом (затуханием). Импульсное (распространяющееся) возбуждение - присуще высокоорганизменным тканям, возникает под действием порогового и сверхпорогового раздражителей.Особенности импульсного возбуждения: имеет латентный период - между моментом нанесения раздражения и видимой ответной реакцией проходит некоторое время; имеет порог раздражения; не градуально - изменение заряда клеточной мембраны не зависит от силы раздражителя; наличие рефрактерного периода; импульсное возбуждение не затухает. Вывод:в организме животного и человека наблюдается местное и импульсное возбуждение. Возникновение того или иного вида возбуждения зависит от степени развития ткани и силы раздражителя.
Чем больше сила раздражения, тем больше, до известных пределов, величина ответных реакций. 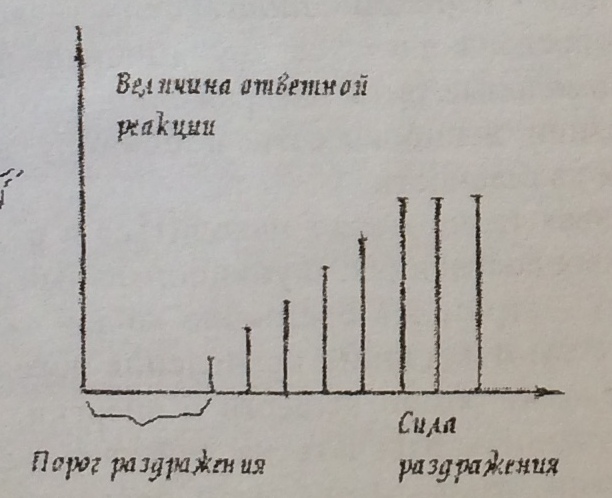
Чем больше сила раздражителя, тем меньше по времени может действовать данный раздражитель для получения ответной реакции. 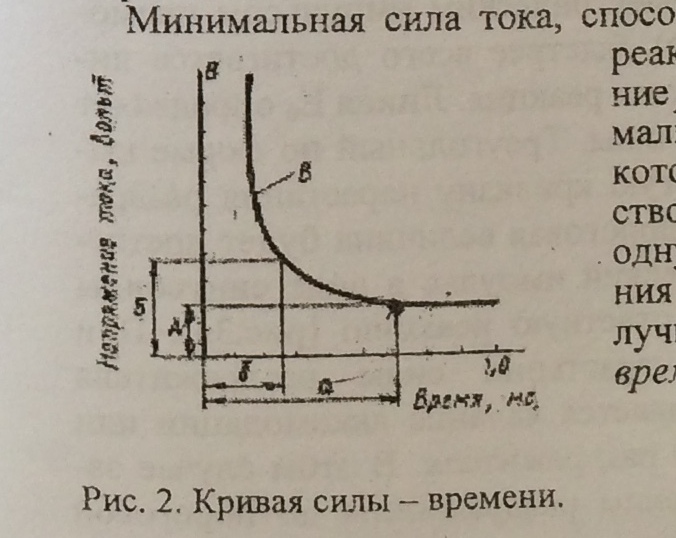
Чем быстрее нарастает сила раздражителя до пороговой величины, тем быстрее появляется ответная реакция. 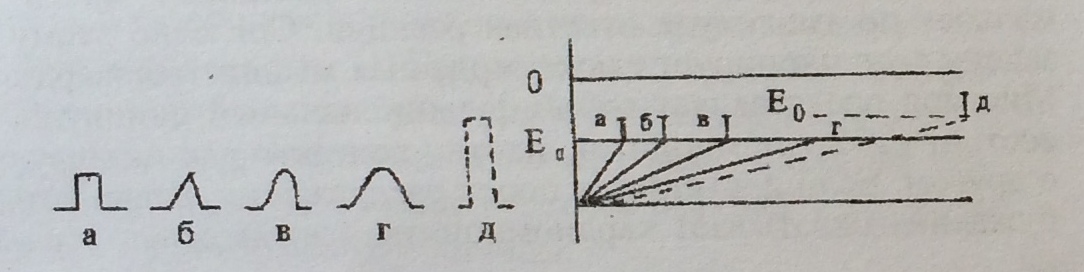
Нейрон состоит из тела диаметром от 3 до 100 мкм, содержащего ядро и другие органеллы, и отростков. Выделяют два вида отростков. (Аксон — длинный отросток, приспособленный для проведения возбуждения от тела нейрона. Дендриты — как правило, короткие и сильно разветвлённые отростки, служащие главным местом образования влияющих на нейрон возбуждающих и тормозных синапсов (разные нейроны имеют различное соотношение длины аксона и дендритов). Нейрон может иметь несколько дендритов, но только один аксон. Один нейрон может иметь связи с до 20 тысяч других нейронов. Физиологические свойства Нейроны восприимчивы к раздражению, то есть способны воспринимать раздражитель и отвечать на него генерацией потенциала действия (ПД). Обычно раздражителем для нейрона служит нейромедиатор, выделяемый другими нейронами в синаптические щели. Одни синапсы вызывают деполяризацию нейрона, другие - гиперполяризацию; первые являются возбуждающими, вторые - тормозящими. Обычно для возбуждения нейрона необходимо раздражение от нескольких возбуждающих синапсов. Классификация По положению в рефлекторной дуге различают афферентные нейроны (сенсорные нейроны), эфферентные нейроны (двигательные нейроны) и вставочные нейроны. На основании числа и расположения отростков нейроны делятся на униполярные нейроны, псевдоуниполярные нейроны, биполярные нейроны и мультиполярные нейроны. Все особенности распространения возбуждения в ЦНС объясняются ее нейронным строением: наличием химических синапсов, многократным ветвлением аксонов нейронов, наличием замкнутых нейронных путей. Этими особенностями являются следующие. 1. Иррадиация (дивергенция) возбуждения в ЦНС. Она объясняется ветвлением аксонов нейронов, их способностью устанавливать многочисленные связи с другими нейронами, наличием вставочных нейронов, аксоны которых также ветвятся (рис. 4.4, а). Иррадиацию возбуждения можно наблюдать в опыте на спинальной лягушке, когда слабое раздражение вызывает сгибание одной конечности, а сильное энергичные движения всех конечностей и даже туловища. Дивергенция расширяет сферу действия каждого нейрона. Один нейрон, посылая импульсы в кору большого мозга, может участвовать в возбуждении до 5000 нейронов. 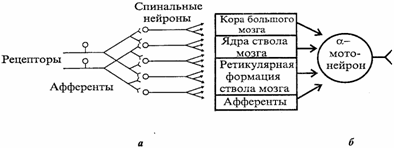 Рис. 4.4. Дивергенция афферентных дорсальных корешков на спинальные нейроны, аксоны которых, в свою очередь, ветвятся, образуя многочисленные коллатерали (в), и конвергенция эфферентных путей от различных отделов ЦНС на α-мотонейрон спинного мозга (6) 1. Конвергенция возбуждения (принцип общего конечного пути)схождение возбуждения различного происхождения по нескольким путям к одному и тому же нейрону или нейронному пулу(принцип шеррингтоновской воронки).Конвергенция возбуждения объясняется наличием многих аксонных коллатералей,вставочных нейронов, а также тем, что афферентных путей в несколько раз больше,чем эфферентных нейронов. На одном нейроне ЦНС может располагаться до 10 000синапсов. Явление конвергенции возбуждения в ЦНС имеет широкое распространение.Примером может служить конвергенция возбуждения на спинальном мотонейроне. Так,к одному и тому же спинальному мотонейрону подходят первичные афферентные волокна (рис. 4.4, б), а также различные нисходящие пути многих вышележащих центров ствола мозга и других отделов ЦНС. Явление конвергенции весьма важно:оно обеспечивает, например, участие одного мотонейрона в нескольких различных реакциях. Мотонейрон, иннервирующий мышцы глотки, участвует в рефлексах глотания, кашля, сосания, чиханья и дыхания, образуя общий конечный путь для многочисленных рефлекторных дуг. На рис. 4.4, я показаны два афферентных волокна, каждое из которых отдает коллатерали к 4 нейронам таким образом, что 3нейрона из общего их числа, равного 5, образуют связи с обоими афферентными волокнами. На каждом из этих 3 нейронов конвергируют два афферентных волокна. На один мотонейрон может конвергировать множество коллатералей аксонов, до 10 000-20 000, поэтому генерация ПД в каждый момент зависит от общей суммы возбуждающих и тормозящих синаптических влияний. ПД возникают лишь в том случае, если преобладают возбуждающие влияния.Конвергенция может облегчать процесс возникновения возбуждения на общих нейронах в результате пространственной суммации подпороговых ВПСП либо блокировать его вследствие преобладания тормозных влияний (см. раздел 4.8). 3. Циркуляция возбуждения по замкнутым нейронным цепям. Она может продолжаться минуты и даже часы (рис. 4.5). 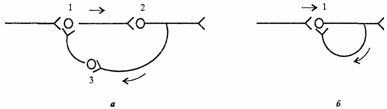 Рис. 4.5. Циркуляция возбуждения в замкнутых нейронных цепях по Лоренто де-Но (а) и по И.С.Беритову (б). 1,2,3возбуждающие нейроны Циркуляция возбуждения одна из причин явления последействия, которое будет рассмотрено далее (см. раздел 4.7). Считают, что циркуляция возбуждения в замкнутых нейронных цепях наиболее вероятный механизм феномена кратковременной памяти (см. раздел 6.6). Циркуляция возбуждения возможна в цепи нейронов и в пределах одного нейрона в результате контактов разветвлений его аксона с собственными дендритами и телом. 4. Одностороннее распространение возбуждения в нейронных цепях, рефлекторных дугах. Распространение возбуждения от аксона одного нейрона к телу или дендритам другого нейрона, но не обратно объясняется свойствами химических синапсов, которые проводят возбуждение только в одном направлении(см. раздел 4.3.3). 5. Замедленное распространение возбуждения в ЦНС по сравнению с его распространением по нервному волокну объясняется наличием на путях распространения возбуждения множества химических синапсов. Время проведения возбуждения через синапс затрачивается на выделение медиатора в синаптическую щель, распространение его до постсинаптической мембраны,возникновение ВПСП и, наконец, ПД. Суммарная задержка передачи возбуждения в синапсе достигает примерно 2 мс. Чем больше синапсов в нейрональной цепочке,тем меньше общая скорость распространения по ней возбуждения. По латентному времени рефлекса, точнее по центральному времени рефлекса, можно ориентировочно рассчитать число нейронов той или иной рефлекторной дуги. 6. Распространение возбуждения в ЦНС легко блокируется определенными фармакологическими препаратами, что находит широкое применение в клинической практике. В физиологических условиях ограничения распространения возбуждения по ЦНС связаны с включением нейрофизиологических механизмов торможения нейронов. Рассмотренные особенности распространения возбуждения дают возможность подойти к пониманию свойств нервных центров.
Нервные волокна представляют собой от ростки нейронов, с помощью которых осу ществляется связь между нейронами, а также нейронов с исполнительными клетками. В состав нервного волокна входят осевой ци линдр (нервный отросток) и глиальная обо лочка. По взаимоотношению осевых цилинд ров с глиальными клетками выделяют два типа нервных волокон: безмиелиновые и миелиновые. Оболочку безмиелиновых волокон образуют шванновские клетки (леммоциты). При этом осевые цилиндры прогибают кле точную оболочку леммоцитов и погружаются в них. Клеточная мембрана обычно полнос тью окружает каждый осевой цилиндр и смы кается над ним, образуя сдвоенную мембрану (мезаксон Наиболее распространена классификация по Дж.Эрлангеру и Х.Гассеру (1937), в которой волокна разделяют на три типа: А, В и С (табл.3). Волокна типа А и В являются миелиновыми, типа С — безмиелиновыми. Во локна А делят на 4 подгруппы: α, β, γ, δ. В пе риферической нервной системе к волокнам Аα относятся афферентные волокна от механорецепторов кожи, мышечных и сухожиль ных рецепторов, а также эфферентные во локна к скелетным мышцам. К Аβ принадле жат афферентные волокна от кожных рецеп торов прикосновения и давления, от части мышечных и висцеральных рецепторов. Аγ представляют собой эфферентные волокна, через которые регулируется активность мы шечных рецепторов. К Аδ относят афферент ные волокна от части тактильных, темпера турных и болевых, а также суставных рецеп торов. К волокнам типа В принадлежат преганглионарные волокна вегетативной нерв ной системы. К волокнам типа С относят постганглионарные волокна вегетативной нерв ной системы, афферентные волокна от неко торых болевых (вторичная боль), тепловых и висцеральных рецепторов. Таблица 3. Типы волокон в нервах млекопи тающих (по Эрлангеру—Гассеру)
Из данных, представленных в табл.3, видно, что средний диаметр каждого типа во локна снижается от типа А до С (каждый примерно в 2 раза по отношению к предыду щему). Соответственно этому снижается и скорость проведения возбуждения. Низкая скорость проведения нервного импульса в волокнах типа С связана с особенностями проведения возбуждения в безмиелиновых волокнах. Лабильность также уменьшается от волокон Аα до С и находится в обратной за висимости от продолжительности фазы абсо лютной рефрактерности. Возбудимость тоже уменьшается от волокон Аα (наибольшая возбудимость) к волокнам С (наименьшая возбудимость). Например, пороговая сила электрического тока у волокон С в 30—50 раз больше, чем у волокон Аα. Исследование факторов, блокирующих нервную проводи мость, показало, что к давлению наиболее чувствительны волокна А, к кислородному голоданию (гипоксии) — волокна В, к мест ным анестетикам — волокна С. Нервные волокна имеют две основные функции — проведение возбуждения и транспорт веществ, обеспечивающих трофи ческую функцию
Нервная система человека делится на центральную (головной и спинной мозг) и периферическую. Центральная нервная система обеспечивает индивидуальное приспособление, поведение организма в конкретных условиях среды обитания, регулирует деятельность каждого органа, обеспечивает интеграцию и объединение органов в единую систему, согласует интенсивность функционирования систем организма, обеспечивает реагирование организма как единого целого на раздражители из внешней и внутренней среды организма. Общей функцией ЦНС является ее трофическое влияние на клетки. В органах, лишенных связей с ЦНС, развиваются дистрофические, воспалительные и атрофические процессы, приводящие к снижению и прекращению функциональной активности органов. Рефлекс – закономерная реакция организма на изменение внешней и внутренней среды, осуществляемая при участии нервной системы в ответ на раздражение рецепторов. В процессе рефлекторной реакции воспроизводится, изменяется интенсивность или прекращается деятельность тканей, органов или организма в целом. При помощи рефлекса устанавливается адекватное соотношение активности органов в пределах системы, систем в пределах организма, организма в его взаимоотношениях с окружающей средой. Рефлекторный ответ осуществляется за минимальное время и с максимальной безошибочностью. Морфологическим субстратом рефлекса является рефлекторная дуга. Ее звенья: 1. Афферентное (рецепторы и афферентный нейрон). 2. Центральное (вставочные нейроны и синапсы). 3. Эфферентное (эффекторный нейрон и эффектор). Простейшая (моносинаптическая) рефлекторная дуга имеет два нейрона: афферентный и эфферентный и один синапс. Рефлекторные дуги большинства рефлексов полисинаптические. Область тела, раздражение которой вызывает определенный рефлекс, называется рецептивным полем рефлекса (рефлексогенной зоной). Нервный центр – совокупность нейронов, необходимых для осуществления определенного рефлекса или регуляции той или иной функции. Время от начала раздражения рецептора до появления ответной реакции называется латентным периодом рефлекса. Более медленное проведение возбуждения по рефлекторной дуге, чем по нерву, связано с явлением синаптической задержки, необходимой для: 1) выделения медиатора нервным окончанием в ответ на импульс; 2) диффузии медиатора через синаптическую щель к постсинаптической мембране; 3) возникновения возбуждающего постсинаптического потенциала. Вместе с формированием пикового потенциала на мембране нейрона это время составляет 1,5 – 2,0 мс. Время, необходимое для проведения возбуждения по центральной части рефлекторной дуги (с аффекторных нейронов на эффекторные), зависит от количества вставочных нейронов и называется центральным временем рефлекса. Классификация рефлексов По биологическому значению: пищевые, половые, оборонительные, локомоторные, позно-тонические, ориентировочные. В зависимости от расположения рецепторов: экстрарецептивные, интеррецептивные и проприорецептивные. В зависимости от того, какие отделы мозга необходимы для осуществления рефлекса: спинальные, бульбарные, мезенцефальные, кортикальные. В зависимости от отдела нервной системы, который реализует ответ: соматические или вегетативные. По характеру ответной реакции: моторные, секреторные, сосудодвигательные. Моторные рефлексы по длительности ответной реакции разделяются на фазические и тонические. По приспособительному значению рефлексы делятся на безусловные и условные.
Синапс — специализированный контакт между нервными клетками или нервными клетками и другими возбудимыми образованиями, обеспечивающий передачу возбуждения с сохранением его информационной значимости. С помощью синапсов осуществляется взаимодействие разнородных по функциям тканей организма, например нервной и мышечной, нервной и секреторной. ^ Структура синапса. Пресинаптическое окончание аксона нейрона при подходе к иннервируемой клетке теряет миелиновую оболочку, что несколько снижает скорость распространения волны возбуждения. Небольшое утолщение на конце волокна, называемое синоптической бляшкой, содержит синаптические пузырьки размером 20—60 нм с медиатором — веществом, способствующим передаче возбуждения в синапсе. ^ Синаптическая щель — пространство между пресинаптическим окончанием и участком мембраны эффекторной клетки является непосредственным продолжением межклеточного пространства. ^ Постсинаптическая мембрана — участок эффекторной клетки, контактирующий с пресинаптической мембраной через синаптическую щель. Классификация синапсов. В соответствии с морфологическим принципом синапсы подразделяют на: • аксо-аксональные (между двумя аксонами); • аксодендритические (между аксоном одного нейрона и дендритом другого); • аксосоматические (между аксоном одного нейрона и телом другого); • дендродендритические (между дендритами двух или нескольких нейронов); • нервно-мышечные (между аксоном мотонейрона и исчерченным мышечным волокном); • аксоэпителиальные (между секреторным нервным волокном и грану-лоцитом); • межнейронные (общее название синапсов между какими-либо элементами двух нейронов). ^ Все синапсы делят на центральные (в головном и спинном мозге) и периферические (нервно-мышечные, аксоэпителиальные и синапсы вегетативных ганглиев). В соответствии с нейрохимическим принципомсинапсы классифицируют по виду химического вещества — медиатора, с помощью которого происходит возбуждение и торможение эффекторной клетки. ^ По способу передачи возбуждениясинапсы подразделяют на три группы. Первую составляют синапсы с химической природой передачи посредством медиаторов (например, нервно-мышечные); вторую — синапсы с передачей электрического сигнала непосредственно с пре- на постсинаптическую мембрану. Третья группа представлена «смешанными» синапсами, сочетающими элементы как химической, так и электрической передачи. По конечному физиологическому эффекту, а также по изменению потенциала постсинаптической мембраны, различают возбуждающие и тормозные синапсы. ^ Механизм проведения возбуждения в синапсах. Передача возбуждения в химическом синапсе — сложный физиологический процесс, протекающий в несколько стадий. Он включает синтез и секрецию медиатора; взаимодействие медиатора с рецепторами постсинаптической мембраны; инактивирование медиатора. В целом синапс осуществляет последовательную трансформацию электрического сигнала, поступающего по нервному волокну, в энергию химических превращений на уровне синаптической щели и постсинаптической мембраны, которая затем снова трансформируется в энергию распространяющегося возбуждения в эффекторной клетке. Свойства синапсов.
Медиаторами, или нейротрансмиттерами, нейронов ЦНС являются различные биологически активные вещества. В зависимости от химической природы их можно разделить на 4 группы: 1) амины (ацетилхолин, норадреналин, дофамин, серотонин), 2) аминокислоты (глицин, глутаминовая, аспарагиновая, гамма-аминомасляная - ГАМК), 3) пуриновые и нуклеотиды (АТФ); 4) нейропептиды (вещество Р, вазопрессин, опоидни пептиды и др.). Раньше считали, что во всех окончаниях одного нейрона "выделяется один медиатор (по принципу Дейла). За последние годы выяснили, что во многих нейронах может содержаться 2 медиаторы или больше. По действию медиаторы можно разделить на ионотропных и метаболотропни. Ионотропных медиаторы после взаимодействия с циторецепторамы постсинаптической мембраны изменяют проницаемость ионных каналов. Метаболотропни медиаторы постсинаптическую действие проявляют путем активации специфических ферментов мембраны. Вследствие этого в мембране или (чаще) в цитоплазме клетки активируются так называемые вторичные посредники (вторичные мессенджеры), которые в свою очередь запускают каскады внутриклеточных процессов, тем самым влияя на функции клеток. К основным мессенджеров систем внутриклеточной сигнализации относят аденилатциклазной и полифосфоинозитидну. В основе первой лежит аденилатциклазной механизм. Центральным звеном второй системы является кальциймобилизуючий каскад полифосфоинозитидив, который контролируется фосфолипазой С. Физиологический эффект этих систем осуществляется путем активации специфических ферментов - протеинфосфокиназ, конечным итогом чего является широкий спектр воздействия на белковые субстраты, которые могут подвергаться фосфорилированию. Вследствие этого изменяется проницаемость мембран для ионов, синтезируются и выделяются медиаторы, регулируется синтез белков, осуществляется энергетический обмен и т.д.. Метаболотропним эффектом обладают большинство нейропептидов. Метаболические изменения, происходящие в клетке или на ее мембране под действием метаболотропних медиаторов, длительные, чем при действии ионотропных медиаторов. Они могут затрагивать даже геном клетки. По функциональным свойствам медиаторы ЦНС делятся на возбуждающие, тормозные и модулирующие. Возбуждающими медиаторами могут быть различные вещества, которые вызывают деполяризацию постсинаптической мембраны. Важнейшее значение имеют производные глутаминовой кислоты (глутамата), субстанция Р. Некоторые центральные нейроны имеют холинорецепторы, т.е. содержат на постсинаптической мембране рецепторы, которые реагируют с холинового соединениями, например, ацетилхолин в клетках Реншоу .. возбуждающими медиаторами могут быть также моноамины (норадреналин, дофамин , серотонин). € основания считать, что тип медиатора, который образуется в синапсе, обусловлен не только свойствами окончания, но и общим направлением биохимических процессов во всем нейроне. Природа тормозного медиатора до конца не установлена. Полагают, что в синапсах различных нервных структур эту функцию могут выполнять аминокислоты - глицин и ГАМК.
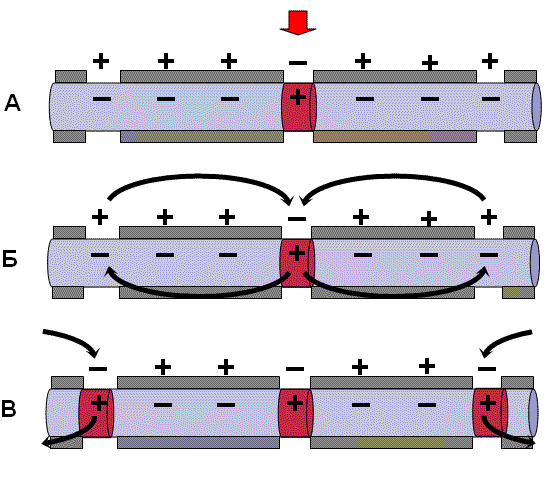
Кроме синапсов с химической передачей возбуждения преимущественно в центральной нервной системе (ЦНС) встречаются синапсы с электрической передачей. Возбуждающим электрическим синапсам свойственны очень узкая синаптическая щель и очень низкое удельное сопротивление сближенных пре- и постсинаптических мембран, что обеспечивает эффективное прохождение локальных электрических токов. Низкое сопротивление, как правило, связано с наличием поперечных каналов, пересекающих обе мембраны, т. е. идущих из клетки в клетку (щелевой контакт). Каналы образуются белковыми молекулами (полумолекулами) каждой из контактирующих мембран, которые соединяются комплементарно. Эта структура легко проходима для электрического тока. Механизм передачи возбуждения в электрическом синапсеМеханизм проведения возбуждения аналогичен механизму проведения возбуждения в нервном волокне. Во время развития ПД происходит реверсия заряда пресинаптической мембраны. Электрический ток, возникающий между пресинаптической и постсинаптической мембраной, раздражает постсинаптическую мембрану и вызывает генерацию в ней ПД (рис. 2).
Нервный центр — это совокупность связанных между собой нейронов, совместно выполняющих определённую функцию путём преобразования входящего возбуждения в выходящее с изменёнными характеристиками. Наличие входов и выходов для возбуждения. В нервном центре можно различить приносящие (афферентные) входы и выносящие (эфферентные) выходы. Одно стороннее проведение возбуждения. Это свойство отдельного синапса и нервной цепи. В нервном центре может быть множество путей между входами и выходами. За счёт обратных связей возможно возвратное движение возбуждения. Но это происходит внутри нервного центра. А если рассматривать нервный центр целиком, то возбуждение приходит внего по приходящим путям, а выходит по эфферентным выходящим. Таким образом, можно говорить об одностороннем проведении возбуждения нервным центром. Задержка (замедление) проведения возбуждения. В нервных центрах возникает задержка в проведении возбуждения, так называемый латентный (скрытый) период. Задержка обусловлена синаптической передачей возбуждения. Чем больше синапсов участвует в проведении возбуждения, тем более длительной получается задержка. Суммация возбуждения. Если одновременно подавать возбуждение на несколько входов нервного центра, то на выходе можно получить более сильное возбуждение. Свойством суммации обладает и отдельный нейрон за счёт суммации локальных потенциалов. Трансформация (преобразование) входящего возбуждения в иное - выходящее. Нервный центр осуществляет изменение, перекодирование поступающих в него потоков импульсов. Трансформация возбуждения - это, пожалуй, самое главное свойство нервного центра. Наиболее известное свойство из этого ряда – трансформация ритма. Нервный центр получает на входе один ритм импульсации, а на выходе дает другой (более медленный или более частый). Последействие (облегчение). Это означает, что после возбуждения нервного центра он некоторое время ещё сохраняет повышенную возбудимость. Поэтому последующее возбуждение даёт более сильный эффект и получение эффекта от работы нервного центра облегчено. Утомляемость и низкая лабильность. Лабильность - это предельная частота импульсации, доступная данной нервной структуре. Нервные центры могут пропускать через себя потоки возбуждения с ограниченной частотой импульсации вследствие задержки передачи возбуждения, которая происходит в многочисленных синапсах. Повышенная утомляемость нервных центров объясняется высокой утомляемостью синапсов и ухудшением метаболизма (обменнных процессов) в нейронах после нагрузки. Тонус. Это означает, что даже без внешнего воздействия нервный центр сохраняет определённый уровень возбудимости и самостоятельно поддерживает у себя определённый уровень возбуждения. Чувствительность к кис лороду и к действию биологически активных веществ (нейротропных). Это создаёт предпосылки к хеморегуляции - химическому управлению деятельностью нервного центра. Например, усиление или ослабление кровоснабжения изменяет работу нервных центров. Возбудимость (воз буждение). Это способность нервных центров переходить в более возбуждённое состояние, например, при внешнем воздействии на них (стимуляции) или под влиянием других нервных центров. Торможение ("тормозимость"). Это способность нервных центров переходить в менее возбуждённое состояние, например, при внешнем воздействии на них или под влиянием других нервных центров. Иррадиация возбуждения. Это "растекание возбуждения" по нервному центру, распространение возбуждения на новые участки от места его первоначального появления. Конвергенция (схождение). Это объединение двух или нескольких входящих потоков возбуждения в один выходящий поток. Т.е. в нервный центр входит больше потоков возбуждения, че выходит из него. Дивергенция (расхождение). Это разделение входящего потока возбуждения на несколько выходящих потоков. За счёт дивергенции получается, что в нервный центр входит меньше потоков возбуждения, чем выходит из него.
Нейроглия. В процессе развития тканей нервной системы из материала нервной трубки, а также нервного гребня происходит развитие глиобластов. Результатом глиобластической дифференцировки является образование нейроглиальных клеточных дифферонов. Они выполняют опорную, разграничительную, трофическую, секреторную, защитную и другие функции. Нейроглия создает постоянную, стабильную внутреннюю среду для нервной ткани, обеспечивая тканевый гомеостаз и нормальное функционирование нервных клеток. По строению и локализации клеток различают эпендимную глию, астроцитную глию и олигодендроглию. Нередко эти разновидности глии объединяют обобщенным понятием "макроглия". Эпендимная глия имеет эпителиоидное строение. Она выстилает центральный канал спинного мозга и мозговые желудочки. В качестве эпендимного эпителия эта разновидность нейроглии относится к нейроглиальному типу эпителиальных тканей. Выпячивания мягкой оболочки мозга в просвет его желудочков покрыты эпендимоцитами кубической формы. Они принимают участие в образовании спинномозговой жидкости. В стенке Ш-го желудочка мозга находятся специализированные клетки — танициты, обеспечивающие связь между содержимым желудочка и кровью за счет ультрафильтрации элементов спинномозговой жидкости. Астроцитная глия является опорной структурой (каркасом) спинного и головного мозга. В астроцитной глии различают два вида клеток: протоплазматические и волокнистые астроциты. Первые из них располагаются преимущественно в сером веществе мозга. Они имеют короткие и толстые, часто распластанные отростки. Вторые — находятся в белом веществе мозга. Волокнистые астроциты имеют многочисленные отростки, содержащие аргирофильные фибриллы. За счет этих фибрилл формируются глиальные остов и разграничительные мембраны в нервной системе, пограничные мембраны вокруг кровеносных сосудов и так называемые "ножки" астроцитных отростков на кровеносных сосудах. Олигодендроглия состоит из различно дифференцированных клеток — олигодендроцитов. Они плотно окружают тела нейронов и их отростки на всем протяжении до концевых разветвлений. Есть несколько видов олигодендроцитов. В органах центральной нервной системы олигодендроглия представлена мелкими отростчатыми клетками, называемыми глиоцитами. Вокруг тел чувствительных нейронов спинномозговых ганглиев находятся глиоциты ганглия (мантийные глиоциты). Отростки нервных клеток сопровождают нейролеммоциты, или шванновские клетки. Источник их развития в периферических нервах, по данным некоторых авторов, эктомезенхима нервного гребня. Функции олигодендроглиоцитов многообразны и чрезвычайно важны для нормальной деятельности нервных клеток. Они обеспечивают трофику нейронов. В единой метаболической системе "нейрон-глия" происходит взаимообмен некоторыми ферментами, белками и РНК. Олигодендроциты играют существенную роль в процессах возбуждения и торможения нейронов и проведения по их отросткам нервных импульсов. Так, нейролеммоциты совместно с отростками нейронов образуют миелиновые и безмиелиновые нервные волокона периферической нервной системы, выполняя при этом роль изоляторов, препятствующих рассеиванию импульсов. Олигодендроциты принимают участие в регуляции водно-солевого баланса в нервной системе. Они могут набухать, перераспределять ионы и т. д. Специализированные глиоциты нервных окончаний участвуют в процессах рецепции, а также в передаче нервного импульса на рабочие структуры. Помимо макроглии в нервной системе есть еще микроглия. Источником ее развития является мезенхима, а клетки микроглии представляют собой глиальные макрофаги и относятся к нейроглии лишь на основании гистотопографии. Клетки микроглии могут размножаться, проявлять фагоцитарную активность, синтезировать не свойственные организму антигены, что наблюдается при некоторых заболеваниях.
Торможение в центральной нервной системеИнтегративная и координационная деятельность центральных нервных образований осуществляется при обязательном участии тормозных процессов. Торможение в центральной нервной системе — активный процесс, проявляющийся внешне в подавлении или в ослаблении процесса возбуждения и характеризующийся определенной интен сивностью и длительностью. Торможение в норме неразрывно связано с возбуждением, яв ляется его производным, сопутствует возбудительному процессу, ограничивая и препятствуя чрезмерному распространению послед него. При этом торможение часто ограничивает возбуждение и вместе с ним формирует сложную мозаику активированных и за торможенных зон в центральных нервных структурах. Формирую щий эффект тормозного процесса развивается в пространстве и во времени. Торможение — врожденный процесс, постоянно совершен ствующийся в течение индивидуальной жизни организма. При значительной силе фактора, вызвавшего торможение, оно может распространяться на значительное пространство, вовлекая в тормозной процесс большие популяции нервных клеток. История развития учения о тормозных процессах в центральной нервной системе начинается с открытия И. М. Сеченовым эффекта центрального торможения (химическое раздражение зрительных бугров тормозит простые спинномозговые безусловные реакции). Вначале предположение о существовании специфических тормозных нейронов, обладающих способностью оказывать тормозные влияния на другие нейроны, с которыми имеются синаптические контакты, диктовалось логической необходимостью для объяснения сложных форм координационной деятельности центральных нервных образо ваний. Впоследствии это предположение нашло прямое эксперимен тальное подтверждение (Экклс, Реншоу), когда было показано су ществование специальных вставочных нейронов, имеющих синап тические контакты с двигательными нейронами. Активация этих вставочных нейронов закономерно приводила к торможению двига тельных нейронов. В зависимости от нейронного механизма, способа вызывания тормозного процесса в ЦНС различают несколько видов торможения: постсинаптическое, пресинаптическое, пессимальное. Постсиналтическое торможение — основной вид торможения, развивающийся в постсинаптической мембране аксосоматических и аксодендритических синапсов под влиянием активации тормозных нейронов, в концевых разветвлениях аксонных отростков которых освобождается и поступает в синаптическую щель тормозной меди атор. Тормозной эффект таких нейронов обусловливается специфи ческой природой медиатора — химического переносчика сигнала с одной клетки на другую. Наиболее распространенным тормозным медиатором является гамма-аминомасляная кислота (ГАМК). Хи мическое действие ГАМК вызывает в постсинаптической мембране эффект гиперполяризации в виде тормозных постсинаптических по тенциалов (ТПСП), пространственно-временная суммация которых повышает уровень мембранного потенциала (гиперполяризация), приводит к урежению или полному прекращению генерации рас пространяющихся ПД. Возвратным торможением называется угнетение (подавление) активности нейрона, вызываемое возвратной коллатералью аксона нервной клетки. Так, мотонейрон переднего рога спинного мозга прежде чем покинуть спинной мозг дает боковую (возвратную) ветвь, которая возвращается назад и заканчивается на тормозных нейронах (клетки Реншоу). Аксон последней заканчивается на мо тонейронах, оказывая на них тормозное действие. Пресинаптическое торможение развертывается в аксоаксональных синапсах, блокируя распространение возбуждения по аксону. Пресинаптическое торможение часто выявляется в структурах моз гового ствола, в спинном мозге Пессимальное торможение представляет собой вид торможения центральных нейронов. Оно наступает при высокой частоте раздра жения. В первый момент возникает высокая частота ответного воз буждения. Через некоторое время стимулируемый центральный ней рон, работая в таком режиме, переходит в состояние торможения. |