Статья Р.Б. Семенова и Л.С. Лепшокова. Отавность высокогорных растений и микориза
 Скачать 237.12 Kb. Скачать 237.12 Kb.
|

|
ОТАВНОСТЬ ВЫСОКОГОРНЫХ РАСТЕНИЙ И МИКОРИЗА Р.Б. Семенова, Л.С. Лепшокова ФГБОУ ВПО «Карачаево-Черкесский государственный университет имени У.Д. Алиева», г. Карачаевск Характер трофического воздействия животных и растений связан со спецификой воздействия животных на растительность, которая определяется, с одной стороны, отчуждением фитофагами живой растительной массы, и, с другой, реакцией растений на это отчуждение. От того, какую часть растительной массы изымают животные и как растительность реагирует на отчуждение этой массы, зависит результат взаимодействия животных и растений. Для растительности, сформировавшейся в условиях постоянного пастбищного воздействия животных, отчуждение части растительной продукции не только не наносит ущерба ее продуктивности, но даже необходимо для поддержания ее высокого уровня. И, наоборот, растительные сообщества, в состав которых входят не приспособленные к воздействию пастбищных животных растения, не выносят даже незначительного отчуждения массы. Не вызывает сомнения, что различные виды растений значительно различаются по своей способности выносить отчуждение растительной массы. Что касается фитоценозов в целом, формируемых такими растениями, то очевидно, что и они допускают без ущерба для своей продуктивности тот же уровень отчуждения. И, наконец, устойчивость растений к изъятию в значительной степени определяется видовым составом растительности, формирующимся в зависимости от режима использования. Из сказанного следует, что уровень «выносливости» растительного покрова к отчуждению меняется вместе с изменением видового состава растений и зависит от пастбищной нагрузки растительноядных животных (Абатуров, 1980). Влияние доступности почвенных ресурсов на отавность растений. На отавность растении значимое влияние оказывает доступность почвенных ресурсов. Симбиоз с микоризообразующими грибами является важной формой консортивных связей растений, обитающих в условиях почв, бедных элементами минерального питания (Селиванов, 1976, 1981; Chapin, 1980; Moser, Haselwandter, 1983; Harley, 1984). При изучении потока углерода в ризосферу и почву под Poa pratensis после дефолиации в Yellowstone национальном парке было показано, что дефолиация увеличивает поступление органических соединений в ризосферу, что ведет к усилению микробиологической активности и увеличению содержания минерального азота (скорость минерализации азота увеличилась вдвое), активно поглощаемого корнями растений. Ранее было показано, что дефолиация усиливает корневые выделения у нескольких злаков. После дефолиации значимо возрастало общее (микроорганизмы + корни) дыхание почвы (Hamilton, Frank, 2001). Изучение взаимоотношении микоризации, фитофаги и растений микрокосмах высокотравной прерии показало, что микориза увеличивала отавность растений после дефолиации и некоторые растения с высокой степенью микоризной инфекции показывали сверхкомпенсационный рост в присутствии микоризных грибов. Надземные фитофаги существенно увеличивали колонизацию корней микоризными грибами (Meyer, 2000) [Kula et al., 2005]. Высокогорные почвы обычно содержат мало доступных для растений соединений азота и фосфора, поэтому микосимбиотрофизм альпийских растений имеет большое значение как приспособление к получению элементов минерального питания. Был поставлен 13-недельный лабораторный эксперимент для проверки зависимости реакции растений на дефолиацию от видового состава сообщества и присутствия грибов АМ. Тестовые растения – Phleumpratense и Trifoliumrepens и их смешанные посевы. Видовой состав растений (монокультуры и смесь) влиял на микоризацию корней, она была максимальна в монокультуре клевера, средняя в смеси и минимальная – в монокультуре тимофеевки. Дефолиация не повлияла на микоризацию клевера, но увеличила микоризацию тимофеевки и смешанной культуры. Дефолиация увеличила содержание азота в побегах тимифеевки и в смешанном посеве, но не у клевера в монокультуре. Содержание углерода в побегах после дефолиации снизилось на 2,7%. Потребители бактерий (нематоды) были на 45% более обильны на вариантах с дефолиацией, чем без нее. Авторы делают вывод, что дефолиация негативно сказывается на продукции растений, но позитивно на нематодах, остальные реакции были видоспецифичными (Hokka et al., 2004). Методика исследования Отавность растений Для изучения отавности был проведен трехлетний полевой эксперимент в альпийских сообществах для 42 видов альпийских растений. Опыт для каждого вида проводили по схеме 1 контроль и 2 варианта (одно- и двукратное) срезание за сезон в разные сроки. На контрольных площадках в первые два года производили только подсчет побегов растений, а их срезание было выполнено только на третий год. В первом варианте растения срезали один раз в начале июля, во втором варианте срезали два раза: в начале июля и в начале августа. Варианты опыта отмечали римскими цифрами (I, II), а сроки срезания – арабскими (1, 2). Во всех случаях оценивали массу срезаемых побегов после высушивания в термостате в течение 8 часов при 90-105° С. Срезание проводили у поверхности почвы ножницами. Перед срезанием учитывали численность вегетативных и генеративных побегов. Для сравнительной интегральной оценки устойчивости растений к дефолиации мы использовали балльные характеристики. Для общей биомассы и средней массы отдельного побега на третий год эксперимента видам присваивались следующие баллы: при величинах восстановления 10% и менее от исходного состояния – 0 баллов, 11-25% - 1 балл, 26-74% - 2 балла, 75% и более – 3 балла. Поскольку значения восстановления численности побегов отличались от таковых для биомассы, для численности вегетативных побегов использовалась следующая шкала: 25% и менее от начальной численности побегов на третий год эксперимента – 0 баллов, 26-74% - 1 балл, 75-100% - 2 балла, более 100% - 3 балла. Таким образом, каждому виду присваивали по 3 оценки для каждого параметра (общая биомасса, численность побегов, масса побега) и варианта (I-1, II-1, II-2).(см. Текеев и др., 2011). Данные по восстановлению биомассы, численности побегов и массы отдельных побегов для всех исследованных нами растений представлены в отдельных публикациях (Текеев, 2005, Аджиева, Онипченко, 2006). Изучение микориз Полевые сборы большинства корней растений проведены в августе 1984 г. А.С. Байкаловой. Сбор и дальнейшую обработку проводили по методике И.А. Селиванова (1981). Корни растения выкапывали, осторожно очищали от почвы и фиксировали в 4% растворе формалина. Число изученных особей каждого вида было не менее пяти. Фиксировали не всю корневую систему, а наиболее тонкие корни (до 2 мм толщины). Камеральная обработка заключалась в мацерации и микроскопическом исследовании окрашенных корней. До окрашивания корни были подвергнуты мацерации в 20% растворе КОН без нагревания (этот метод предложен В.Г. Логиновой, личное сообщение). Пробы сразу после сбора отмывали от фиксатора и заливали щелочью на 2-3 месяца. Этого времени мацерации достаточно для мягких бесцветных корешков. Темноокращенные корни мацерировали дольше. Наиболее жесткие и темные корни подвергали горячей мацерации в течение часа в том же растворе щелочи. Далее корни отмывали и окрашивали по способу Л.Н. Курсанова в видоизменении Н.В. Лобанова и Л.Г. Крюгер. Красителем служил раствор Кобеля (0.1 г анилиновой сини, 50 г молочной кислоты, 100 г воды). Отмытые от красителя корни заливали на 30 мин молочной кислотой для дифференциации окраски. Обработанные таким образом корни хранили в глицерине. Для приготовления препаратов брали корни толщиной не более 0,5 мм, так как в более толстых корнях гриб обычно не встречается. Приготовленные препараты просматривали под микроскопом при увеличении Х120. Количество эндофита в поле зрения определялось по пятибалльной шкале, предложенной И.А. Селивановым (1981) и основанной на относительной оценке числа клеток мезодермы, занятых микоризообразующим грибом. Всего просматривалось 130-150 полей зрения в каждой пробе. Исходя из полученных данных, рассчитывали интенсивность микоризной инфекции для каждого вида по формуле (Селиванов, 1981): С= S*100/(N*K) где S – сумма баллов по всем полям зрения, N – число просмотренных полей зрения, K – наивысший балл шкалы учета (5). Результаты исследования Значимая связь между отавностью и интенсивностью микоризной инфекции на наших данных не была обнаружена (рис. 1, 2). 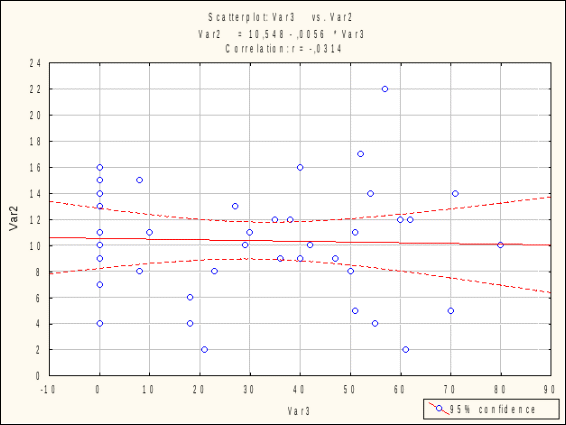 Рис. 1. Связь интенсивности микоризной инфекции (%, абсцисса) с отавностью альпийских растений (баллы, ордината) для видов, образующих микоризу. Анализируя полученные результаты, легко заметить разнообразие поведения видов. Представленные данные (табл.1) свидетельствуют о том, что как наиболее устойчивые (Geraniumgymnocaulon, Trifoliumpolyphyllum, Oxytropis kubanensis, Anemonespeciosa,Taraxacumstevenii), так и наименее устойчивые к дефолиации виды (Antennaria dioica, Festuca ovina, Festuca varia,Sibbaldiaprocumbens, Gnaphaliumsupinum, Nardus stricta, Hedysarum caucasicum) имеют высокие показатели микоризной инфекции. 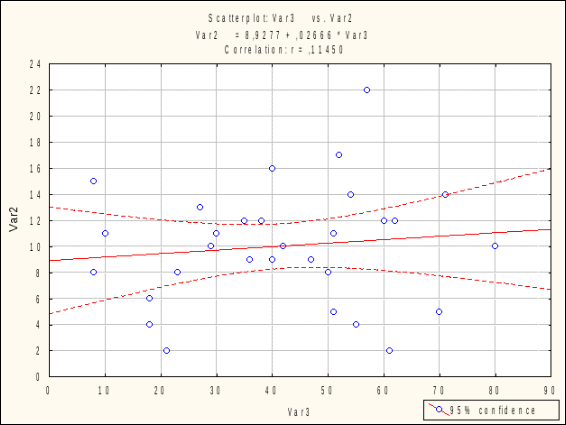 Рис. 2. Связь интенсивности микоризной инфекции (%, абсцисса) с отавностью альпийских растений (баллы, ордината) для видов, образующих микоризу В то же время один из наиболее устойчивых к дефолиации видов - Hyalopoapontica относится к слабомикосимбиотрофным. Среди микоризообразующих видов растений Galium verum относится как к слабомикоризным видам, так и к наименее устойчивым к дефолиации. Сравнение отавности и микоризы сообществ показало, что лучше восстанавливаются после дефолиации виды, растущие на более продуктивных гераниево-копеечниковых лугах. Слабее отрастают виды, приуроченные к альпийским лишайниковым пустошам. Виды, произрастающие на пестроовсяницевых лугах и альпийских коврах имеют примерно одинаковую низкую степень отавности (рис. 3). Таблица 1 Характеристика исследованных видов (сообщество: АЛП – альпийские лишайниковые пустоши, ПЛ - пестроовсяницевые луга, ГКЛ – гераниево-копеечниковые луга, АК – альпийские ковры)
Луговые фитоценозы (ГКЛ, ПЛ) имеют близкие значения общих микосимбиотических показателей. По всем показателям альпийские ковры и пустоши характеризуются меньшей степенью развития микосимбиотрофии. На коврах отмечена наименьшая абсолютная и относительная микоризная инфекция, а также самый низкий процент микоризообразующих видов (таб. 2). Таблица 2 Микосимбиотические показатели альпийских фитоценозов (сообщество: АЛП – альпийские лишайниковые пустоши, ПЛ - пестроовсяницевые луга, ГКЛ – гераниево-копеечниковые луга, АК – альпийские ковры)
Основной вывод нашей работы заключается в том, что как наиболее устойчивые, так и наименее устойчивые к дефолиации виды альпийских сообществ могут иметь высокие показатели микоризной инфекции. Низкую степень отавности имеют растения пестроовсяницевых лугов и альпийских ковров, а низкими показатели развития микориз обладают растения альпийских лишайниковых пустошей и альпийских ковров. Литература 1. Абатуров Б.Д. Особенности трофических взаимодействий типа «фитофаги-растения» в экосистемах пастбищ.- В кн.: Фитофаги в растительных сообществах. М.: Наука, 1980, с. 31-42. 2. Аджиева Р.Б., Онипченко В.Г. Реакция растений альпийских ковров и гераниево-копеечниковых лугов на разные режимы дефолиации // Бюлл. Московск. о-ва испытателей природы, отд. биологический, 2006, т. 111, вып.1, с. 24-36. 3. Байкалова А.С. Микосимбиотрофизм растений альпийских пустошей северо-западного Кавказа // Бюлл. МОИП. Отд. биол. 1985. Т. 90. №5. С. 87-90. 4. Байкалова А.С., Онипченко В.Г. Микосимбиотрофизм альпийских растений Тебердинского заповедника // Опыт исследования растительных сообществ в заповедниках / Тр. ЦНИЛ Главохоты РСФСР. – М., 1988. – С. 93-107. 5. Селиванов И.А. Микосимбиотрофизм как форма консортивных связей в растительном покрове Советского Союза: Дис…д-ра биол. наук. Пермь, 1976. 297с. В надзаг.: Пермский гос. пед. ин-т. 6. Селиванов И.А. Микосимбиотрофизм как форма консортивных связей в растительном покрове Советского Союза. М.: Наука, 1981. 231 с. 7. Селиванов И.А. Проблемы изучения консортивных отношений в природных и антропогенных экосистемах // Микориза и другие формы консортивных связей в природе. Пермь,1983. С. 3-12. 8. Текеев Д.К. Реакция растений альпийских лишайниковых пустошей и пестроовсяницевых лугов на разные режимы дефолиации с. 107-130. Павлов В.Н., Онипченко В.Г, Елумеева Т.Г. (ред.). Альпийские экосистемы: структура и механизмы функционирования / Труды Тебердинского гос. биосферного заповедника, вып. 30. М.: б.и., 2005, 207 с. 9. Chapin F.S. The mineral nutrition of wild plants // Ann. Rev. Ecol. and System. 1980. V. 11. P. 233-260. 10. Harley J.L. The mycorrhizal associations // Cell. Interact. – Berlin e. a., 1984. P. 148-186. 11. Haselwandter K. Mycorrhizal status of ericaceous plants in alpine and subalpine areas // New Phytolog. 1979. V. 83. P. 427-431. Руководитель: к.б.н., доцент кафедры биологии и химии Семенова Р.Б. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||