Систематика ботаники. 120. ординация фитоценозов, её принципы
 Скачать 113.57 Kb. Скачать 113.57 Kb.
|

|
120. ординация фитоценозов, её принципы. Ординация растительности – упорядочение фитоценозов вдоль некоторых осей, определяющих характер варьирования растительности. Методы ординации делятся на прямые (ординация ведется по реальным факторам среды – экологическим, пространственным, временным) и непрямые одномерные (ординация вдоль одного фактора или одной оси) и многомерные (вдоль нескольких факторов или осей). В качестве примера прямой одномерной ординации можно рассмотреть эдаффитоценотические ряды, разработанные В.Н. Сукачевым Фитоценотический ряд показывает, как ассоциации или группы ассоциаций последовательно располагаются в зависимости от изменения какого-либо фактора. Проанализируем эдафо-фитоценотический ряд еловых лесов. ряд А характеризуется увеличением сухости и уменьшением богатства почвы, ряд В –увеличением увлажнения и ухудшением почвенной аэрации, ряд С – увеличением почвенного богатства и аэрации, D –увеличением увлажнения проточной водой, Е – переходный ряд от застойного увлажнения к проточному. В качестве примера прямой двухмерной ординации рассмотрим разработанную для лесов Полесья эдафическую сетку П.С. Погребняка, который развивал взгляды Е.В. Алексеева. В основе лесотипологической системы Алексеева – Погребняка лежат два фактора: трофность (плодородие почв) и влажность. По трофности выделяют четыре ступени: боры, субори, сложные субори и дубравы и др. По влажности выделяют шесть ступеней: ксерофильные (очень сухие), мезо-ксерофильные (сухие), мезофильные (свежие), мезо-гигрофильные (влажные), гигрофильные (сырые) и ультрагигрофильные (болота). Для того чтобы понять, что собой представляют непрямые методы ординации, мы рассмотрим т.н. висконсинскую сравнительную ординацию, которая была разработана Дж. Кертисом. Эти описания сообществ определяют противоположные, концевые точки первой оси. Все остальные описания ранжируются между ними, а их координаты на первой оси определяются по теореме Пифагора. Пара концевых точек второй оси выбирается из следующих соображений: оба описания должны находиться в средней части первой оси, сходство между ними должно быть минимально, их координаты относительно первой пары точек – близки. В дальнейшем все описания ранжируются вдоль второй оси по тем же правилам. Ординация не может заменить классификацию, т.к. ее результаты не позволяют осуществлять различные формы инвентаризации растительности. Но ординация может облегчить процесс классификации. 119. правила наименования синтаксонов. Подходы к классификации растительности
Основными единицами этой классификации являются формация – совокупность сообществ с одним доминантом и ассоциация, которая выделяется по 2–3 доминантам. Самая крупная синтаксономическая единица – тип растительности, который выделяется по эколого-морфологическому принципу. Тип растительности делится на классы формаций, которые выделяются по близости жизненных форм эдификаторов. Классы формаций далее разделяются на группы формаций. Формация – основная синтаксономическая единица «среднего» ранга. Формации далее разделяются на группы ассоциаций. Они выделяются по общности эдификаторов древесного и основного напочвенного ярусов. Наименьшей единицей классификации является ассоциация, к которой относятся фитоценозы с однородным видовым составом, одинаковой структурой, приуроченные к сходным условиям местообитания.
Основной таксономической категорией, или основной единицей растительности, в системе Браун-Бланке является ассоциация. Ассоциация – единица крупная, широкого объема, подразделяемая в свою очередь на более мелкие единицы. Она выделяется на основании анализа всего флористического состава и отличается свойственными ей характерными видами. Вышестоящие таксоны, относящиеся к союзам, порядкам, классам, имеют свои характерные виды. Чем выше ранг, тем большим числом характерных видов обладает данный таксон. Класс – это единица с выраженной собственной физиономией, флористическая комбинация при установлении класса стоит на втором месте. Порядок – крупный вариант класса, устанавливаемый на основе флористических критериев. Союз чаще всего – географический или экологический вариант порядка. Нижестоящие таксоны, относящиеся к субассоциациям,вариантам ассоциаций и фациям, не имеют своих характерных видов и отличаются лишь дифференциальными видами. 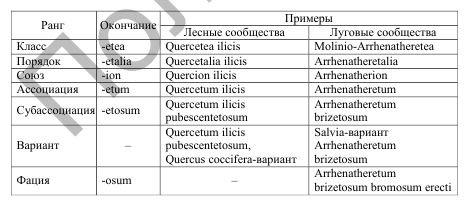 118. основные принципы эколого-флористической классификации. принцип эколого-флористической классификации основан на дедуктивно-индуктивном подходе, при котором главную роль играет процесс установления синтаксонов «снизу». Это происходит путем группирования сообществ по сходству флористического состава, который в значительной степени отражает экологические условия и стадию сукцессии. При этом упор делается на том, что различные виды по-разному связаны с различными условиями произрастания, т.е. с различными фитоценозами, формирующимися в данных условиях. Браун-Бланке различает пять уровней верности: Верные виды (степень верности 5) – виды, исключительно или почти исключительно связанные с определенным типом сообществ. Прочные виды (степень верности 4) – виды, встречающиеся и в других сообществах, но более редко и в меньшем числе, или с пониженной жизненностью. Благосклонные виды (степень верности 3) – виды, которые встречаются более или менее обильно во многих фитоценозах, но оптимальное их развитие связано с одним определенным типом сообщества. Неопределенные виды (степень верности 2) – виды без явно выраженной связи с каким-либо определенным типом сообществ. Чуждые виды (степень верности 1) – редкие и большей частью случайные включения из других растительных сообществ или же реликты ранее бывших растительных сообществ. Система Браун-Бланке предполагает, что каждое растительное сообщество экологически специфично, и данная специфичность проявляется во флористическом наборе, уникальном для каждого конкретного типа местообитания. Виды, характерные для конкретного синтаксона, называются диагностическими. Они, в зависимости от уровня верности, разделяются на три группы: 1. Характерные виды встречаются только в одном синтаксоне или встречаются в этом синтаксоне чаще, чем в других, благодаря своей экологической специализации. 2. Дифференцирующие виды диагностируют синтаксон границей своего ареала и входят в состав нескольких синтаксонов. 3. Константные виды встречаются с высоким постоянством и часто с высоким обилием, но проходят через этот синтаксон «транзитом». ?117. основные синтаксоны доминантной классификации и критерии их выделения. Основными единицами этой классификации по Браун-Бланке являются формация – совокупность сообществ с одним доминантом и ассоциация, которая выделяется по 2–3 доминантам. Самая крупная синтаксономическая единица – тип растительности, который выделяется по эколого-морфологическому принципу. Тип растительности делится на классы формаций, которые выделяются по близости жизненных форм эдификаторов. Классы формаций далее разделяются на группы формаций. Формация – основная синтаксономическая единица «среднего» ранга. Формации далее разделяются на группы ассоциаций. Они выделяются по общности эдификаторов древесного и основного напочвенного ярусов. Наименьшей единицей классификации является ассоциация, к которой относятся фитоценозы с однородным видовым составом, одинаковой структурой, приуроченные к сходным условиям местообитания. 116. понятие о растительной ассоциации как основной таксономической единице растительности. Основная единица в фитоценологии – растительная ассоциация. Ассоциация – это типологическая единица, тип фитоценоза. Каждый фитоценоз, относящийся к той или иной ассоциации, наряду с общими, свойственными всем сходным фитоценозам признакам, может обладать и некоторыми случайными, индивидуальными, которые при характеристике ассоциации отбрасываются. На Брюссельском ботаническом конгрессе в 1910 г. было предложено считать ассоциацию основной единицей растительного покрова. В.Н. Сукачев, в 1938 г. писал: Растительная ассоциация, или тип фитоценоза, объединяет фитоценозы, характеризующиеся однородным составом, строением и в основном одинаковым сложением составляющих их синузий, и имеющие одинаковый характер взаимоотношений как между растениями, так и между ними и средой». Т.е. ассоциация при такой трактовке должна включать фитоценозы, сходные по своей структуре, флористическому составу, доминантам в каждом ярусе, сезонной и разногодичной динамике и по условиям существования. Ассоциация – единица крупная, широкого объема, подразделяемая в свою очередь на более мелкие единицы. Она выделяется на основании анализа всего флористического состава и отличается свойственными ей характерными видами. ?115.подходы и принципы классификации. 1. Физиогномический подход. Главный тип сообщества планеты, выделяемый по физиогномическим признакам – биом – совокупность сообществ и среды их обитания в пределах одной природной зоны. Биомы выделяются на основании господства одной жизненной формы растений. По данному признаку существует несколько основных физиогномических типов сообществ: 1) леса (доминант – деревья); 2) злаковники (доминанты – травы, как правило, злаки и осоки); 3) редколесья (невысокие деревья, обычно образующие несомкнутый покров с хорошо развитым нижним ярусом); 4) кустарники (преобладают кустарники при общем проективном покрытии растений, как правило, не выше 50 %); 5) пустыни (покрытие растений очень низкое, обычно не выше 10 %). 2.Доминантный и доминантно-детерминантный подход. В основу положен эколого-физиогномический принцип, причем более всего применяется схема, основывающаяся при выделении единиц всех рангов по жизненной форме доминантных видов господствующей синузии. Основными единицами этой классификации являются формация – совокупность сообществ с одним доминантом и ассоциация, которая выделяется по 2–3 доминантам. Самая крупная синтаксономическая единица – тип растительности - классы формаций - группы формаций - Формация – группы ассоциаций - ассоциация. 3. Паллиативный доминантно-детерминантный метод учитывает, с одной стороны, характер доминантности различных видов, а с другой стороны – довольно широко использовал такое понятие, как виды-детерминанты. Используя как доминантный, так и детерминантный критерий, Б.М. Миркин предложил следующую иерархию таксономических единиц эколого-морфологической классификации растительности:
4. Эколого-флористический подход (система Браун-Бланке) основан на дедуктивно-индуктивном подходе, при котором главную роль играет процесс установления синтаксонов «снизу». Класс – Порядок – Союз – ассоциация - субассоциация - вариант ассоциаций – фация. 114.принципы и методы классификации растительности. В фитоценологии, изучающей растительный покров с точки зрения слагающих его растительных сообществ, классификация (синтаксономия) в очень сильной степени отличается от классификации отдельных организмов (таксономии). Типы сообществ, которые выделяются в процессе классификации (синтаксоны), – это абстрактные классы, в каждом из которых сгруппировано определенное количество сообществ на основе присущих им свойств. Сообщества можно классифицировать по целому ряду признаков:
При методических подходах к классификации растительности различают два основных способа классификации: 1. Дедуктивный способ – классификация осуществляется «сверху», т.е. путем деления имеющегося множества объектов на все более мелкие классы. 2. Индуктивный способ – классификационная схема – начинает строиться с низших таксономических единиц. В этом случае исследователь заранее не знает тех признаков, которые он положит в основу выделения тех или иных таксонов. ?113. Суть взглядов Ф. Клементса сводилась к следующему: в пределах одной климатической зоны все фитоценозы в ходе сукцессии должны конвергировать к одному климаксовому сообществу. Климаксовые устойчивые фитоценозы Ф. Клементс противопоставлял серийным, т.е. сукцессионным. Смена серийных фитоценозов приводит к климаксу. 112.аллогенные сукцессии: гейтогенез и гологенез. Аллогенные сукцессии разделяются на гологенез и гейтогенез. Гологенез – сукцессии, являющиеся следствием процессов развития ландшафтов, обусловленных как природными, так и антропогенными причинами (например, формирование речной поймы, строительство водохранилищ и т.д.). Гейтогенез – сукцессии, обусловленные внешними факторами, которые не связаны с общими тенденциями развития ландшафта и имеют главным образом антропогенную природу. Аллогенные сукцессии обусловлены влиянием внешнего фактора, который действует на фитоценоз. При этом чаще всего, по мере ухудшения условий среды, происходит смена видов в направлении их более высокой патиентности и приспособленности к фактору, вызвавшему сукцессию. 111. Автогенные сукцессии. Модели автогенных сукцессий. Сукцессии со сменой модели. Автогенные сукцессии очень разнообразны, они могут быть первичными и вторичными, прогрессивными и регрессивными, быстрыми и медленными. Как правило, они необратимы и протекают постоянно. Несмотря на спонтанность этих сукцессий, их может вызвать также человек, повреждающий растительность. При этом на месте нарушения начинаются восстановительные автогенные сукцессии. Все разнообразие автогенных сукцессионных процессов можно свести к четырем принципиальным моделям сукцессии, которые были предложены Дж. Коннелом и П. Слэйтьером и Д. Боткиным.
В большинстве же случаев разные периоды сукцессионного процесса протекают в соответствии с разными моделями. При этом сукцессия, как правило, начинается с модели благоприятствования или нейтральности и заканчивается моделью толерантности. Это связано с тем, что на самых ранних стадиях автогенных сукцессий конкуренция между растениями чаще всего ослаблена. Но в дальнейшем происходит обострение конкурентных взаимоотношений между видами. 110.сукцессии растительности. Их классификация. Сукцессии – постепенные направленные, как правило, необратимые изменения растительности, которые могут быть вызваны как внутренними, так и внешними причинами, при которых изменяется видовой состав или количественное соотношение видов, т.е. структура фитоценозов. Сукцессии можно классифицировать по достаточно большому числу критериев, например: по масштабу времени:
по обратимости:
по степени постоянства процесса:
по происхождению:
по характеру изменения структуры и видового состава:
по антропогенности:
по причинам:
109.разногодичная изменчивость фитоценозов. Флуктуации, их основные признаки. Причины. Типы флуктуации в соответствии с причинами их возникновения и по степени выраженности. Возрастные изменения фитоценозов. Параметры любого растительного сообщества не остаются неизменными, а колеблются год от года. Их значения в разные годы оказываются то большими, то меньшими по отношению к среднему значению за определенный, но не слишком большой период, т.е. колеблются около средней характеристики. Изменяются сроки прохождения фенологических фаз, обилие видов и их соотношение, фитомасса, иногда происходит изменение и во флористическом составе. Причина, определяющая эти изменения, заключается в колебаниях погодных условий – количества солнечной энергии, поступающей к растениям, температурного режима, количества осадков. Меняющиеся условия существования дают при этом преимущества то одним, то другим видам, что приводит в изменении системы взаимодействий между растениями. Термин «флуктуация» впервые применил в 1939 г. Х. Глизон. Различают следующие типы флуктуаций в соответствии с причинами их возникновения:
Возрастная изменчивость фитоценозов, по В.Н. Сукачеву – это «изменение фитоценозов в связи с онтогенезом эдификаторов». Разногодичные изменения при увеличении амплитуды колебаний условий существования могут перейти в смены одного сообщества другим. Смены, вызванные возобновлением того или иного вида, при гибели большей части генеративных особей, также способны привести к смене фитоценозов. 108.сезонная изменчивость фитоценозов, её причины. Этот тип изменчивости фитоценозов связан с изменением условий произрастания растений в течение года и с присутствием в составе фитоценозов видов растений, различающихся по ритму сезонной вегетации. Изменение условий произрастания в течение года может быть обусловлено:
107.горизонтальная структура фитоценозов. мозаичность фитоценозов, ее причины и степень выраженности в разных типах фитоценозов. Комплексность растительного покрова. Для объяснения природы горизонтальной структуры сообщества очень удобна несложная модель вариации растительности, разработанная В.И. Василевичем. В основе этой модели лежит закон разложения дисперсии, на котором основан метод дисперсионного анализа: σ2 = σа2 + σb2 + σz2, где σа2 – это доля вариации растительности, которая порождена изменениями условий среды. В этом случае чередуются разные сообщества и их сочетание называется комплексом. Комплексность порождает β-разнообразие растительности. σb2 – это доля дисперсии, которая вызвана непосредственно эффектами взаимодействия растений (конкуренция и др). В результате действия этих внутренних факторов, если они достаточно сильны, формируются закономерно повторяющиеся пятна растительности, которые называют микрогруппировками, а их сочетание называется мозаичностью. При этом повышается α-разнообразие. σz2 – это т.н. «шум», т.е. случайные изменения растительности, связанные с малоуловимыми вариациями условий среды, случайностями при разбросе семян и т.д. Мозаичность может быть плавной, с нерезкими границами между микрогруппировками, и контурной, с относительно резкими границами. В настоящее время различают несколько типов мозаичности по ее происхождению. Фитогенная мозаичность обусловлена конкуренцией, изменением фитосреды или спецификой жизненных форм растений (способность к вегетативному размножению и образованию клонов). Эдафотопическая мозаичность связана с неоднородностями почвы как среды обитания (неровности микрорельефа, различный дренаж, неоднородность почв и подстилки, их мощности, гумусированности, гранулометрического состава и т.д.). Зоогенная мозаичность вызывается влиянием животных, как прямым, так и косвенным, – выеданием, вытаптыванием, откладыванием экскрементов, деятельностью землероющих животных и т.д. Антропогенная мозаичность связана с деятельностью человека – вытаптыванием вследствие рекреационной нагрузки, выпасом сельскохозяйственных животных, выкашиванием травяных и вырубкой лесных растительных сообществ, ресурсными заготовками и т.д. Экзогенная мозаичность обусловлена внешними абиотическими факторами среды – влиянием ветра, воды и т.д. Очень широко в природе распространена демутационная мозаичность, связанная с восстановлением растительности в нарушенных местах. В отличие от мозаичности, которая характеризует внутриценотическую горизонтальную неоднородность, комплексность – это неоднородность растительного покрова на надфитоценотическом уровне. Она проявляется в закономерном чередовании в пределах одного ландшафта отдельных фитоценозов. Комплексность растительного покрова определяется микро- или мезорельефом. 106.синузии. синузиальная структура фитоценоза. Синузия – это структурно-функциональная часть фитоценоза, которая обособлена в пространстве по вертикали или во времени и отличается, кроме этого, морфологической, флористической, экологической и фитоценотической обособленностью. Синузии образованы растениями одной или близких жизненных форм, поэтому входящие в их состав растения обладают, как правило, сходными потребностями и сходно трансформируют среду в благоприятную для себя и своих партнеров сторону. Полисинузиальность (т.е. существование в фитоценозе одновременно нескольких синузий) характерна для сложных ярусно-дифференцированных сообществ, сформированных различными жизненными формами (например, леса). В таких случаях синузия представляет собой не что иное, как ярус или его часть (например, сезонные синузии). Травяные сообщества, как правило, моносинузиальны. По Т. Липпмаа, фитоценозы представляют собой комбинации элементарных синузий («одноярусных ассоциаций»), а абстрактные типы фитоценозов (ассоциации) – комбинации типов синузий. По признаку автономности выделяют различные типы синузий – как тесно связанные с определенными типами фитоценозов, так и сравнительно самостоятельные, зависящие в основном от условий экотопа. 105.вертикальный колонтинуум. Причины формирования. В противоположность ярусным лесам умеренной зоны в природе достаточно часто встречаются фитоценозы, в которых ярусность абсолютно отсутствует или выражена очень слабо – такие фитоценозы формируют вертикальный континуум. Ярким примером этого могут быть тропические леса, масса и листовая поверхность в которых располагается непрерывно и скопления надземных органов (в том числе листьев) отсутствуют. В таких лесах от поверхности почвы и до верхней границы сообщество буквально «набито» листьями разных видов. Фитоценозами, в которых практически отсутствует ярусность, является также и большинство травяных сообществ. Отсутствие расчленения на ярусы в них связано с:
Ярусное распределение растений снижает конкуренцию между ними и обеспечивает устойчивость сообществ. 104.вертикальная структура фитоценоза. Ярусность. Подходы к выделению ярусов. Фитоценотические горизонты. Вертикальная структура фитоценозов характеризуется объемом среды, в котором размещены органы растений, и особенностям размещения в отдельных горизонтах среды их массы, объема, поверхности соприкосновения со средой. Объем используемой среды определяется высотой надземных органов растений и варьирует в широких пределах. используемый объем может быть однородным в пределах всего фитоценоза или изменяться в вертикальном направлении. На сегодняшний день общепризнанно, что ярусное расчленение фитоценозов является результатом коэволюции видов, которые приспособились к использованию света различной интенсивности. При этом ярусное расчленение фитоценоза представляет собой экологическое дополнение одних видов другими, т.е. ведет к более полному использованию ресурсов окружающей среды, в первую очередь света. Помимо количества и состава света, ярусы будут также различаться особенностями теплового режима, влажности воздуха, содержанием углекислого газа и т.д. Различные ярусы будут также различаться по высоте и мощности. Существуют два подхода к выделению ярусов. При первом подходе основанием для отнесения растений к определенным ярусам служат группы особей, достаточно однородные по размещению их органов ассимиляции в отдельных горизонтах среды. При этом различные группы особей одного и того же вида в зависимости от того, в каком горизонте сосредоточена основная масса их ассимилирующих органов, могут участвовать в формировании не одного, а нескольких ярусов. При втором подходе ярусы рассматриваются, как отграниченные друг от друга горизонты фитоценоза, в которых сосредоточена основная масса ассимилирующих органов растений. При этом неравномерность распределения фотосинтезирующих органов растений является основанием для выделения ярусов. При таком подходе ярусы представляет собой фитоценотические горизонты – структурные части биогеоценотических горизонтов. Каждый фитоценотический горизонт характеризуется не только составом видов, но и составом органов этих растений. Кроме того, каждый горизонт будет характеризоваться свойственными ему средой и взаимоотношениями с консортами (фитофагами, паразитными грибами, эпифитами и др.). Так, в лесах можно выделить следующие фитоценотические горизонты: 1) кроновый 2) стволовой 3) травяной, травяно-кустарничковый или кустарничковый 4) припочвенный. 103.типы ценопопуляции по их онтогенетическому составу. Понятие о полночленности и неполночленности ценопопуляции. 1. Инвазионные ценопопуляции. В ценопопуляции подобного типа основную часть растений представляют собой виргинильные особи. Растения приживаются в фитоценозе, еще только обосновываются в нем, и потому не совершают своего полного цикла развития. 2. Ценопопуляции нормального типа. В ценопопуляциях этого типа представлены особи всех возрастных групп, т.е. в данном случае растения проходят весь цикл своего развития. При этом процент генеративных особей будет очень сильно зависеть от степени благоприятности условий окружающей среды. 3. Ценопопуляции регрессивного типа. В ценопопуляциях этого типа генеративное возобновление растений прекратилось. При этом растения могут еще цвести и плодоносить, но появляющиеся проростки очень быстро гибнут, либо растения не образовывают жизнеспособных семян, или же цветут, но не плодоносят. Наконец, растения в ценопопуляции могут вообще утратить способность к цветению и лишь вегетировать. Иногда выделяется еще один специальный тип – ложноинвазионные ценопопуляции. Они образуются у тех видов растений, которые имеют в почве банк семян или вегетативных зачатков (корневищ, клубней, луковиц и т.д.). Обычно эти растения находятся в состоянии покоя и отсутствуют в составе травостоя. Но если на покоящиеся зачатки растений подействует какой-либо фактор, способный их вывести из покоящегося состояния, то происходит массовое развитие растений – вид может стать доминантом в сообществе. Ценопопуляцию, в которой представлены растения всех возрастных групп, называют полночленной, а при отсутствии определенных групп – неполночленной. Как правило, ценопопуляции инвазийного или регрессивного типа будут неполночленными, т.е. будут отсутствовать либо виргинильные особи (в регрессивных ценопопуляциях), либо генеративные и сенильные (в инвазийных ценопопуляциях). В то же время нормальные ценопопуляции могут быть как полночленными, так и неполночленными. При этом неполночленность ценопопуляции будет временной. 102.виталитет ценопопуляции. Методы его определения. Возрастная структура ценопопуляции. Виталитет (жизненность) ценопопуляции – это жизненность (степень процветания или угнетения) организма. Виталитет выражается и в интенсивности цветения, и образования семян – в т.н. репродуктивном усилии. Определение виталитета ценопопуляции может проводиться по-разному в зависимости от характера выбранного признака. Вначале выбирается признак, отражающий виталитет, и получается выборка из нескольких десятков результатов измерений растений по выбранному признаку. Затем полученную выборку ранжируют от минимума к максимуму и делят вариационный ряд на три равные части – крупные, промежуточные и мелкие особи (а, b и с). По соотношению участия этих групп растений в ценопопуляции оценивается ее виталитет. Особи растений, входящие в состав ценопопуляции, как правило, различаются по возрасту. устанавливается относительный возраст, т.е. тот, на который растение «выглядит». В этом случае говорят о выделении онтогенетических групп растений. У растений, размножающихся семенами, за основу выделения онтогенетических групп принято разграничение жизненного цикла на четыре периода. 1. Латентный (период первичного покоя) – период, когда растение находится в состоянии семян. Ценопопуляция при этом представлена банком семян, находящемся в почве. 2. Виргинильный (девственный) – период от прорастания семян до начала генеративного размножения особи. выделяются растения следующих возрастных состояний: проростки ювенильные растения; имматурные растения; виргинильные растения – растения, у которых появляются основные черты типичного взрослого растения (характерные листья, побеги и корни), но генеративные органы еще отсутствуют. 3. Генеративный – период, когда растение размножается семенами. В его пределах выделяются растения следующих возрастных состояний: молодые генеративные растения средние генеративные растения старые генеративные растения 4. Сенильный (старческий) – период, когда растение уже не способно к семенному размножению и обречено на отмирание. Растения, находящиеся в сенильном периоде, разделяются на: субсенильные сенильные растения отмирающие растения 101.ценопопуляции растений. Плотность ценопопуляции, методы ее определения. Варианты размещения особей по территории. Ценопопуляция, или локальная (фитоценотическая) популяция – это группа особей одного вида на определенной экологически однородной территории, то есть в пределах одного растительного сообщества. Ценопопуляция, как и любая другая биологическая система, имеет свой состав, структуру и определенные закономерности изменения во времени, т.е. определенную динамику. Ценопопуляции между собой могут различаться по плотности, массе, распределению особей в пространстве, соотношению полов (для двудомных растений), виталитету (жизненности), генетической и возрастной структуре. Плотность ценопопуляции – это количество особей, приходящихся на единицу площади. Определение плотности ценопопуляций легко осуществляется у растений, размножающихся только семенным путем. Но у многих видов т.н. клональных растений, помимо семенного, происходит активное вегетативное размножение. В таких случаях счетной единицей при определении плотности ценопопуляции служит не особь, а побег. У большинства растений плотность ценопопуляций регулируется соотношением двух факторов: зависимостью от смертности и пластичностью. Различают четыре основных способа распределения 1. Регулярное распределение – особи ценопопуляции расположены на примерно одинаковом расстоянии друг от друга. 2. Случайное распределение – расстояние между особями подчиняется закону случайного распределения, положение каждой особи не зависит от положения других особей. 3. Групповое распределение (пятнистое, контагиозное) – особи в пространстве распределены группами (куртинами). 4. Клинальное распределение выражается в постепенном и направленном изменении плотности особей на определенной территории. По-видимому, главная причина такого размещения состоит в постепенном изменении в пространстве условий произрастания (например, при движении по склону). Возможны разные варианты клинального распределения. Распределение случайно-клинальное, и контагиозно-клинальное. ?100.состав жизненных форм фитоценоза как показатель свойств экотопа, истории фитоценоза и форм взаимодействия между растениями. причины, определяющие степень экологической неоднородности фитоценоза. Жизненная форма растения – это его внешний облик, комплекс морфологических, анатомических и физиологических признаков, отражающий приспособленность вида к условиям среды. ?99. Степень флористического богатства и ее причины. Видовая насыщенность. Представление о минимальной площади выявления флористического состава и других признаков фитоценоза. Флористический состав характеризуется рядом показателей. Первый – это видовое богатство, т.е. общее количество видов, свойственное фитоценозу. Этот показатель может изменяться в пределах от 1 (монодоминантные одновидовые сообщества) до 1000 и более видов (некоторые тропические леса). В геоботанике гораздо чаще используется показатель видовой насыщенности – число видов, отнесенное к единице площади. Площадь выявления – величина учетной площадки, на которой выявляются все существенные черты фитоценоза (флористический состав фитоценоза, его структура и количественное соотношение видов; в лесных сообществах, кроме этого – запас древесины и кривая распределения деревьев по классам диаметра). Размер площади учета является очень важным фактором, влияющим на видовое богатство фитоценоза. Так, к примеру, в мелком масштабе наибольшим видовым богатством характеризуются альварные луга, встречающиеся в Эстонии и Швеции. Факторы, которые определяют видовое богатство фитоценоза, многочисленны и сложно взаимодействуют. При всем интересе к показателю степени видового богатства очевидно, что использование его в сравнительно-аналитических построениях во многих случаях некорректно. Так, к примеру, несравнимы в отношении видового богатства маленькое болотце и участок тропического леса. 98. Флористическая полночленность и неполночленность фитоценозов. Основные причины неполночленности фитоценозов. Под флористически неполночленными фитоценозами Раменский понимал сообщества, в состав которых входят не все виды растений, способные в них существовать. Им были выделены фитоценозы абсолютно полночленные, туземно полночленные, практически полночленные и явно неполночленные. Полночленность или неполночленность фитоценозов может быть точно установлена лишь при проведении опытов с подсевом семян видов, не входящих в их состав. Абсолютно полночленных фитоценозов в природе, вероятно, не существует. Можно различать первичную и вторичную, или антропически обусловленную неполночленность. Причиной этого может быть следующее: 1. Малый возраст; 2. Отсутствие видов, способных входить в состав фитоценоза в окружающей его растительности, и затруднения в приносе их диаспор из более отдаленных мест. 3. Отмирание особей видов, входивших ранее в состав компонентов фитоценоза и способных произрастать в данных условиях и в настоящее время, что обусловлено длительным отсутствием обсеменения соответствующих видов каким-либо эпизодическим сильным воздействием (длительный застой воды, образование ледяной корки и др.). 97. Факторы формирования флористического состава фитоценоза. Флористический состав луговых фитоценозов зависит от следующих факторов:
96. Взаимоотношения между растениями и их консортами. Специфичность видов по воздействию на среду: эдификаторы и ассектаторы. Как правило, в состав консорций входят бактерии, грибы, животные. Между растениями и непосредственно связанными с ними консортами возникают многочисленные типы взаимоотношений, из которых наиболее важными являются три типа взаимоотношений: 1) оказывающие отрицательное воздействие на растение, но выгодные для консортов (паразитизм и хищничество); 2) взаимовыгодные как для растений, так и для их консортов (мутуализм); 3) выгодные для консортов, но не оказывающие значительного воздействия на растение (комменсализм). В подавляющем большинстве случаев взаимоотношения между растениями и их консортами являются взаимоотношениями организмов различных трофических уровней. Говоря о взаимодействии видов растений друг с другом и с окружающей средой, нельзя забывать о том, что они не только изменяются под действием абиотических и биотических факторов, но и сами способны трансформировать окружающую их среду. При этом каждый вид растений специфичен по способности изменять среду. Специфичность видов в трансформации окружающей среды послужила основой для формирования системы фитоценотипов. Растения, по В.Н. Сукачеву, подразделяются на: эдификаторы – созидатели, строители сообщества, т.е. обильные виды, формирующие внутреннюю среду сообщества и в значительной мере определяющие появление в нем других видов; ассектаторы – виды, мало влияющие на создание фитогенной среды сообщества. В роли эдификатора, помимо отдельных видов, могут выступать и группы, состоящие из растений разных видов, но одной жизненной формы. Виды, входящие в такую группу, называются соэдификаторами. 95. Конкуренция. Роль конкурентных взаимоотношений в формировании растительных сообществ. Аменсализм: одностороннее и взаимное отрицательное средообразование, аллелопатия. 1. Конкуренция – это соревнование организмов за потребление одного или нескольких ресурсов, находящихся в дефиците. Аменсализм – один вид, именуемый аменсалом, претерпевает угнетение роста и развития, а второй, именуемый ингибитором, таким испытаниям не подвержен: одностороннее отрицательное средообразование – пример влияния деревьев-доминантов на виды мохового и травяного ярусов. взаимное отрицательное средообразование – отношения сфагновых мхов и сосудистых растений на сфагновом болоте, когда сфагновые мхи довольно быстро растут, повышают уровень поверхности болота и постепенно погребают в своей толще многолетние живые органы цветковых растений, выступая как ингибиторы. Аллелопатия – это взаимное влияние растений через изменение среды в результате выделения в нее продуктов жизнедеятельности. Это могут быть летучие вещества или вещества, растворенные в воде, стекающей с листьев, либо омывающей корни, а также субстанции, образующиеся или высвобождающиеся в результате распада отмирающих частей растений. 94. Типы взаимоотношений растений друг с другом (классификации В. Н. Сукачева, В. С. Ипатова и Л. Н. Кириковой, Б. М. Миркина).
Ввиду многообразия и сложности взаимоотношений между растениями В.С. Ипатовым и Л.А. Кириковой (1999) была предложена двухуровневая система классификации: I. По субъектам:
II. По способам воздействия:
III.По участию среды:
IV.По роли среды в питании растений:
V. По последствиям для растений:
Б.М. Миркин с соавторами (Миркин и др., 2001) выделяют следующие типы взаимоотношения растений:
93. Взаимоотношения между организмами в фитоценозе. Фитогенное поле, его роль во взаимоотношениях растений. Консорции, их структура. Типы консорций. В результате жизнедеятельности растений, входящих в состав фитоценоза и связанных с ними гетеротрофных организмов, экотоп трансформируется в биотоп. При этом входящие в состав фитоценоза растения, с одной стороны, взаимодействуют друг с другом на уровне так называемых фитогенных полей, а с другой стороны – находятся в сложных многогранных отношениях с гетеротрофными компонентами биоценозов, формируя консорции. Фитогенное поле означает сферу влияния одного растения на окружающую его среду и на занимающие эту среду растения. Радиус фитогенного поля может меняться от нескольких сантиметров у трав до нескольких метров у крупных деревьев. Консорция – это такая совокупность особей различных видов, в центре которой находится особь автотрофного или гетеротрофного вида, компоненты которой связаны с центром трофически, топически, форически или фабрически; под влиянием которой формируется специфическая микросреда. Консорция является элементарной структурной единицей биоценозов, в которой различные организмы объединены друг с другом. Центральная особь консорции называется центральным ядром (детерминантом консорции), а связанные с ней организмы – консортами. консорцию можно рассматривать как структурную единицу биогеоценоза; она имеет свои пространственные и функциональные параметры. Выделяют следующие типы консорций: - индивидуальные (одного растения), - ценопопуляционные (популяции вида в растительном сообществе), - региональные, - видовые. 92. Роль эдафических (механический состав, физические и химические свойства почвы) и орографических (рельеф) факторов в формировании растительного покрова. Почвы и грунты. Эдафические условия определяются свойствами субстрата, на котором произрастают растения. Субстрат может быть лишь незначительно затронут процессами почвообразования (скалы, подвижные пески), или же в результате действия физического выветривания и живых организмов поверхностный слой горной породы может превратиться в почву. Почвы характеризуются рядом физико-химических характеристик, имеющих существенное значение для роста и развития растений, а также для формирования того или иного типа растительного сообщества: 1. Кислотность почвы. По отношению к кислотности почвы как к экологическому фактору выделяются следующие группы растений
2. Валовое содержание питательных веществ (богатство почвы). По отношению к богатству почвы как к экологическому фактору выделяются следующие группы растений:
3. Содержание солей в почве. Растения, приспособленные к высокому содержанию солей в почве, носят название галофитов. Они характеризуются высоким осмотическим давлением клеточного сока. Большинство галофитов либо имеет суккулентное, либо ксероморфное строение. Растения незасоленных почв называются гликофитами.
|