Пассивные свойства мембраны и теория кабеля. Пассивные свойства мембраны и теория кабеля Возбуждение как активная реакция клетки на раздражитель
 Скачать 0.99 Mb. Скачать 0.99 Mb.
|

|
Пассивные свойства мембраны и теория кабеля Возбуждение как активная реакция клетки на раздражитель Реакция клетки на внешнее воздействие (раздражение) отличается от реакции небиологических систем следующими особенностями: • энергией для реакции клетки служит не энергия раздражителя, а энергия, образующаяся в результате метаболизма в самой биологической системе; • сила и форма реакции клетки не определяется силой и формой внешнего воздействия (если сила раздражителя выше пороговой). В некоторых специализированных клетках реакция на раздражитель проявляется особенно интенсивно. Такую интенсивную реакцию называют возбуждением. Возбуждение – активная реакция специализированных (возбудимых) клеток на внешнее воздействие, проявляющаяся в том, что клетка начинает выполнять присущие ей специфические функции. Возбудимая клетка может находиться в двух дискретных состояниях: • состоянии покоя (готовность к реагированию на внешнее воздействие, совершение внутренней работы); • состоянии возбуждения (активное выполнение специфических функций, совершение внешней работы). В организме существует 3 типа возбудимых клеток: • нервные клетки (возбуждение проявляется генерацией электрического импульса); • мышечные клетки (возбуждение проявляется сокращением); • секреторные клетки (возбуждение проявляется выбросом в межклеточное пространство биологически активных веществ). Возбудимость – способность клетки переходить из состояния покоя в состояние возбуждения при действии раздражителя. Разные клетки имеют различную возбудимость. Возбудимость одной и той же клетки меняется в зависимости от ее функционального состояния. При действии слабых (подпороговых) импульсов электрического тока в клетке развивается электротонический потенциал. Электротонический потенциал (ЭП) – сдвиг мембранного потенциала клетки, вызываемый действием постоянного электрического тока. ЭП есть пассивная реакция клетки на электрический раздражитель; состояние ионных каналов и транспорт ионов при этом не изменяется. ЭП не проявляется физиологической реакцией клетки. Поэтому ЭП не является возбуждением. При действии более сильного подпорогового тока возникает более пролонгированный сдвиг мембранного потенциала (МП) – локальный ответ. Локальный ответ (ЛО) – активная реакция клетки на электрический раздражитель, однако состояние ионных каналов и транспорт ионов при этом изменяется незначительно. ЛО не проявляется заметной физиологической реакцией клетки. ЛО называют местным возбуждением, так как это возбуждение не распространяется по мембранам возбудимых клеток. При действии порогового и сверхпорогового тока в клетке развивается потенциал действия (ПД). ПД характеризуется тем, что значение МП клетки очень быстро уменьшается до 0 (деполяризация), а затем мембранный потенциал приобретает положительное значение (+20…+30 мВ), т. е. внутренняя сторона мембраны заряжается положительно относительно наружной. Затем значение МП быстро возвращается к исходному уровню. Сильная деполяризация клеточной мембраны во время ПД приводит к развитию физиологических проявлений возбуждения (сокращение, секреция и др.). ПД называют распространяющимся возбуждением, поскольку, возникнув в одном участке мембраны, он быстро распространяется во все стороны. Механизм развития ПД практически одинаков для всех возбудимых клеток. Механизм сопряжения электрических и физиологических проявлений возбуждения различен для разных типов возбудимых клеток (сопряжение возбуждения и сокращения, сопряжение возбуждения и секреции). Емкостные свойства мембраны Мембрана – это пленка диэлектрика, изолятора, по обеим сторонам которой находится электрический заряд, создающий внутри нее электрическое поле. Мембрана тонкая, поэтому величина поля в ней очень велика, и на единицу площади мембраны приходится значительная энергия -порядка десятка кДж /моль (на перенос одновалентных ионов). Высокая напряженность поля в мембране может изменить конформацию белковых молекул, внутри мембраны, и вызвать различные внутриклеточные эффекты, через каскад сигнальных реакций. Энергия, запасенная в поле, принимает участие в трансформации внутриклеточной энергии, в переходе ее в химическую форму. Изменения электрических характеристик клеточных мембран играют решающую роль в работе мозга, в генерации и распространении нервного импульса, в мышечных сокращениях. Соли в водном растворе диссоциируют на ионы, обладающие положительными и отрицательными электрическими зарядами. Мембрана – плоский конденсатор, емкостные свойства определяются фосфолипидным бислоем, который обеспечивает эффективное разделение и накопление зарядов и электростатическое взаимодействие катионов и анионов посредством электрического поля. М 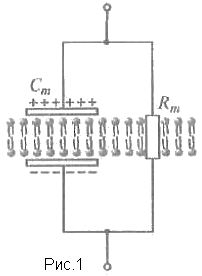 ембрана клетки не только проводит ионные токи, но и накапливает заряд на своей внешней или внутренней поверхности. С точки зрения теории электричества, разделение зарядов на мембране означает, что мембрана обладает свойствами конденсатора. В общем виде конденсатор состоит из двух проводящих пластин, отделенных друг от друга изолирующим материалом. ембрана клетки не только проводит ионные токи, но и накапливает заряд на своей внешней или внутренней поверхности. С точки зрения теории электричества, разделение зарядов на мембране означает, что мембрана обладает свойствами конденсатора. В общем виде конденсатор состоит из двух проводящих пластин, отделенных друг от друга изолирующим материалом. В случае нервной клетки проводниками являются два слоя жидкости, находящиеся по обе стороны мембраны, а сама мембрана играет роль изолирующей прослойки (рис.1.). При зарядке конденсатора от батареи на одной из пластинок накапливается положительный заряд, в то время как на второй пластинке создается равный по величине запас отрицательного заряда. Емкость конденсатора (С) определяется количеством заряда (Q), запасаемым на один вольт потенциала (U), приложенного к пластинам конденсатора: С = Q/U. С измеряется в кулонах, деленных на вольт, т. е. в фарадах (Ф). Чем ближе друг к другу расположены пластины, тем более эффективно конденсатор способен разделять и накапливать заряд. Поскольку толщина мембраны клетки всего 5 нм, она способна накапливать достаточно большой заряд. Обычно емкость мембраны нервных клеток составляет 1 мкФ/см2. Преобразовав выражение, получаем Q = CU. При потенциале покоя —80 мВ, количество избыточного отрицательного заряда на внутренней стороне мембраны составит (1 • 10–6) х (80 •10–3) = 8• 10–8 кулонов, деленных на см2, что соответствует 5 • 1011 одновалентных ионов (0,8 пмоль) на квадратный сантиметр мембраны. Величину тока, протекающего внутрь конденсатора или из него, можно подсчитать на основе соотношения заряда и напряжения, учитывая, что ток (I, в амперах) есть скорость изменения заряда во времени, т.е. 1 ампер = (1 кулон)/(1 с). Поскольку Q = C/U, получим: 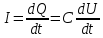 Скорость изменения заряда на конденсаторе прямо пропорциональна величине тока. Если ток постоянен, то потенциал будет меняться с постоянной скоростью: 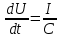 Когда эти два элемента, резистор и конденсатор, соединены параллельно, то весь ток пойдет сначала на зарядку конденсатора со скоростью I/С. Однако, как только на конденсаторе накопится какой-то заряд, ток потечет и через резистор. По мере нарастания тока, все большая его часть будет проходить через сопротивление, потому что скорость зарядки конденсатора будет постепенно снижаться. В конце концов весь ток будет течь через резистор, создавая на нем потенциал U = IR, a конденсатор будет полностью заряжен. По завершении скачка тока заряд из конденсатора постепенно рассеется на резисторе, а напряжение вернется к нулю. Уравнение Ходжкина-Хаксли Возбуждение мембраны описывается уравнениями Ходжкина-Хаксли. В упрощенном виде одно из уравнений Ходжкина-Хаксли имеет вид: 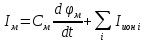 , ,где 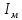 - ток через мембрану, - ток через мембрану, 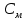 - ёмкость мембраны, - ёмкость мембраны,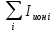 - сумма ионных токов через мембрану. - сумма ионных токов через мембрану. Электрический ток через мембрану складывается из ионных токов: ионов калия - 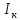 , натрия - , натрия - 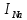 и других ионов (в том числе и других ионов (в том числе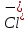 ), так называемого тока утечки ), так называемого тока утечки 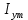 , а также ёмкостного тока , а также ёмкостного тока 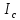 . Ёмкостный ток обусловлен перезарядкой конденсатора – мембраны, перетеканием зарядов с одной ее поверхности на другую. Его величина определяется количеством заряда, перетекающего с одной обкладки на другую за единицу времени . Ёмкостный ток обусловлен перезарядкой конденсатора – мембраны, перетеканием зарядов с одной ее поверхности на другую. Его величина определяется количеством заряда, перетекающего с одной обкладки на другую за единицу времени 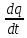 , а поскольку заряд конденсатора , а поскольку заряд конденсатора 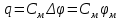 , то ёмкостной ток : , то ёмкостной ток :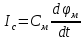 Полный мембранный ток: 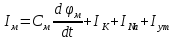 (1) (1) 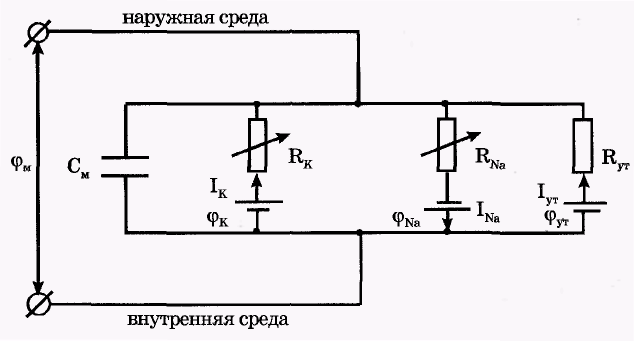 Рис. 2. Эквивалентная электрическая схема элемента возбудимой мембраны Каждый ионный ток определяется разностью мембранного   и равновесного нернстовского потенциала, создаваемого диффузией ионов данного типа и равновесного нернстовского потенциала, создаваемого диффузией ионов данного типа  : : 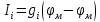 (2), (2),где 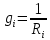 - проводимость (величина, обратная сопротивлению элемента мембраны для ионов данного типа). На эквивалентной электрической схеме элемента мембраны равновесные потенциалы Нернста моделируются источниками напряжений с электродвижущими силами: - проводимость (величина, обратная сопротивлению элемента мембраны для ионов данного типа). На эквивалентной электрической схеме элемента мембраны равновесные потенциалы Нернста моделируются источниками напряжений с электродвижущими силами: 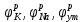 , а проводимости элемента мембраны для разных ионов моделируются резисторами , а проводимости элемента мембраны для разных ионов моделируются резисторами 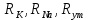 . .Воспользовавшись (2), запишем (1) в виде: 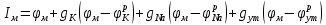 (3). (3).Согласно теории Ходжкина-Хаксли, возбуждение элемента мембраны связано с изменениями проводимости мембраны для ионов 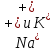 . Так как возбуждение вызывается повышением мембранного потенциала, проводимости ( . Так как возбуждение вызывается повышением мембранного потенциала, проводимости (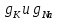 ) мембраны зависят от мембранного потенциала. В разные фазы развития импульса возбуждения ионные проводимости ) мембраны зависят от мембранного потенциала. В разные фазы развития импульса возбуждения ионные проводимости 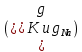 разные, то есть они зависят еще и от времени. разные, то есть они зависят еще и от времени. Кабельные свойства нервных волокон Пропуская электрический ток через введенный в клетку электрод, можно деполяризовать мембрану аксона в каком-то одном участке. Если сила тока мала, тогда деполяризация будет подпороговой, натриевые каналы останутся закрытыми и потенциал действия не возникнет. Установится равновесное состояние, при котором ток, протекающий через микроэлектрод внутрь клетки, сбалансирован током, текущим наружу через мембрану. Часть тока будет выходить вблизи электрода, а часть, прежде чем выйти из клетки, пройдет некоторое расстояние внутри аксона в том или другом направлении. Сила тока, проходящего по какому-либо из этих путей, будет зависеть от их сопротивления. Так как цитоплазма аксона оказывает некоторое сопротивление току, сила тока будет наибольшей вблизи микроэлектрода н наименьшей вдали от него. При таком распределении тока сдвиг мембранного потенциала уменьшается экспоненциально с увеличением расстояния от источника возмущения. Такого рода пассивное распространение электрического сигнала вдоль нервного волокна-без какого-либо усиления, обусловленного открытием потенциал-зависимых каналов, аналогично распространению сигнала по телеграфному кабелю, лежащему на дне моря. По мере прохождения тока по осевому проводнику (цитоплазме) происходит некоторая утечка его через слой изоляции (мембрану) в окружающую среду, так что сигнал постепенно затухает. Поэтому электрические свойства, от которых зависит пассивное распространение сигналов, часто называют кабельными свойствами аксона. Рассмотрим распространение возбуждения на примере передачи нервного импульса по аксону (рис. 3). Если один участок аксона находится в возбужденном состоянии, а соседний находится в покое, тогда между возбужденным и невозбужденным участками волокна течет электрический ток, поскольку внутренняя поверхность первого из них обладает положительным потенциалом относительно второго и между ними существует разность потенциалов. Токи, возникающие в живых тканях вследствие возбуждения, называются локальными (рис. 3, 4), так как распространяются на незначительное расстояние от возбужденного участка. 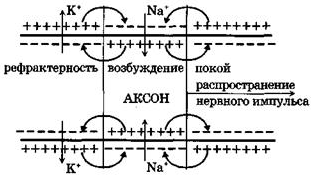 Рис. 3. Распространение нервного импульса по аксону. Их ослабление обусловлено затратами энергии на заряд мембраны и на преодоление сопротивления цитоплазмы волокна. Локальный ток служит раздражителем для покоящихся участков, непосредственно прилежащих к месту деполяризации. В них развивается возбуждение, а значит и новая деполяризация. Она приводит к установлению разности потенциалов между вновь деполяризованным и покоящимся (последующим) участком волокна, вследствие чего возникает локальный ток в следующем микроконтуре. Следовательно, распространение возбуждения представляет собой многократно повторяющийся и охватывающий соседние участки возбудимой мембраны процесс ретрансляции ПД и движения локальных токов от деполяризованных к покоящимся пунктам волокна. 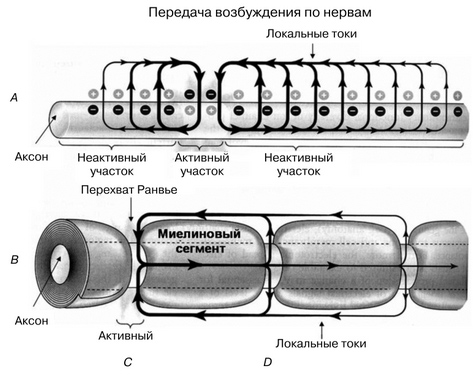 Рис. 4. Локальные токи Распространение потенциала действия (импульса) вдоль нервного волокна определяется кабельными свойствами этого волокна. При локальной деполяризации мембраны и возникновении потенциала действия ток, проходящий через открытые натриевые каналы, пассивно распространяется и деполяризует соседние участки мембраны, где в свою очередь возникает потенциал действия. Во многих аксонах позвоночных высокая скорость и эффективность проведения импульсов достигается благодаря изоляции поверхности аксона миелиновой оболочкой, изменяющей кабельные свойства аксона и оставляющей открытыми лишь небольшие участки возбудимой мембраны. В нервной системе позвоночных нервные волокна большего диаметра обычно миелинизированы. На периферии миелин образуют шванновские клетки, а в ЦНС — олигодендроциты. Эти клетки плотно облегают нейроны, обвиваясь вокруг них. Мембраны так плотно соприкасаются друг с другом, что цитоплазма выдавливается из зазора между ними, в результате чего образуются тесно упакованные спиральные мембранные «обертки». Количество оберток (ламелей) варьирует от 10-20 до 160). Наличие 160 ламелей означает, что между плазматической мембраной аксона и внеклеточной жидкостью последовательно располагается 320 мембран. Таким образом, эффективное сопротивление мембраны возрастает в 320 раз, и во столько же раз снижается мембранная емкость. Миелин составляет обычно от 20 до 40 % общего диаметра волокна. 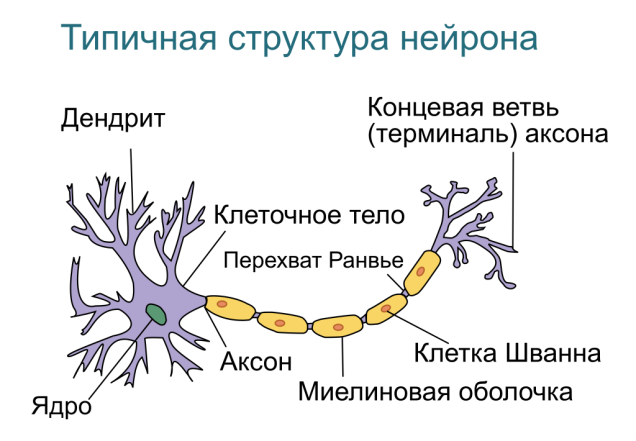 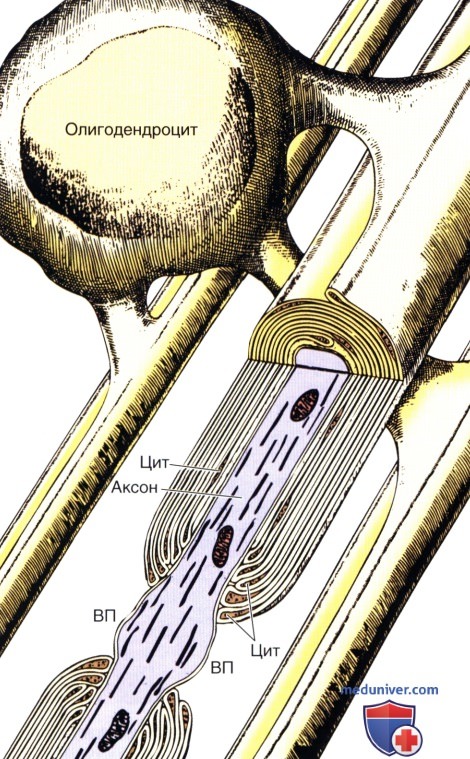 Рис. 5. Структура нервного волокна. М 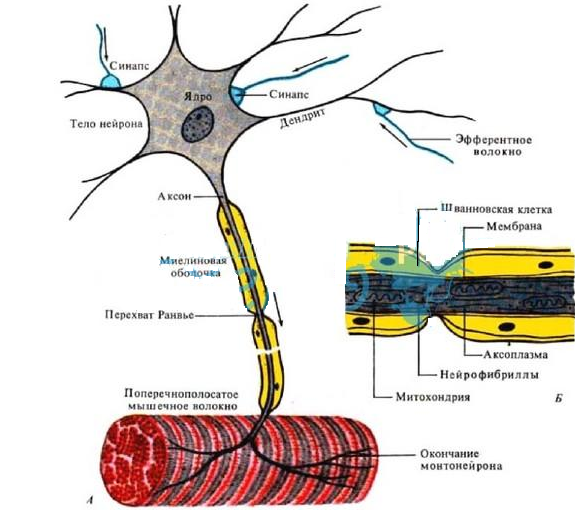 иелиновая оболочка не сплошная, а прерывается через равные промежутки так называемыми перехватами Ранвье, в которых собственная мембрана аксона не покрыта миелином. Расстояние между перехватами обычно в 100 раз превосходит внешний диаметр волокна и составляет от 0,2 до 2 мм. Эти участки шириной всего около 0,5 мкм являются центрами электрической активности. Почти все натриевые каналы аксона сосредоточены в перехватах, где плотность каналов достигает несколько тысяч на 1 мкм2, тогда как в участках, покрытых миелиновой оболочкой, их почти нет. Поэтому изолированные участки мембраны не способны возбуждаться, но обладают превосходными кабельными свойствами - низкой емкостью и высоким сопротивлением для утечки тока. Благодаря этому локальные токи, обусловленные ПД в области перехвата Ранвье, легко доходят (путем пассивного проведения - электротонически) до следующего перехвата, быстро деполяризуют мембрану и возбуждают очередной потенциал действия. Такое проведение, когда сигнал распространяется вдоль аксона, "перескакивая" с одного перехвата на другой, называется сальтаторным. Миелинизация дает два главных преимущества: обеспечивает быстрое распространение ПД и сберегает метаболическую энергию, так как активное возбуждение происходит лишь на небольших участках в перехватах Ранвье. иелиновая оболочка не сплошная, а прерывается через равные промежутки так называемыми перехватами Ранвье, в которых собственная мембрана аксона не покрыта миелином. Расстояние между перехватами обычно в 100 раз превосходит внешний диаметр волокна и составляет от 0,2 до 2 мм. Эти участки шириной всего около 0,5 мкм являются центрами электрической активности. Почти все натриевые каналы аксона сосредоточены в перехватах, где плотность каналов достигает несколько тысяч на 1 мкм2, тогда как в участках, покрытых миелиновой оболочкой, их почти нет. Поэтому изолированные участки мембраны не способны возбуждаться, но обладают превосходными кабельными свойствами - низкой емкостью и высоким сопротивлением для утечки тока. Благодаря этому локальные токи, обусловленные ПД в области перехвата Ранвье, легко доходят (путем пассивного проведения - электротонически) до следующего перехвата, быстро деполяризуют мембрану и возбуждают очередной потенциал действия. Такое проведение, когда сигнал распространяется вдоль аксона, "перескакивая" с одного перехвата на другой, называется сальтаторным. Миелинизация дает два главных преимущества: обеспечивает быстрое распространение ПД и сберегает метаболическую энергию, так как активное возбуждение происходит лишь на небольших участках в перехватах Ранвье. Скорость проведения импульса будет тем выше, чем больше расстояние которое локальный ток, направленный от возбужденного к невозбужденному участку волокна пройдет, сохраняя способность деполяризовать мембрану до порогового уровня. Это расстояние в свою очередь зависит от кабельных свойств аксона и будет тем больше, чем меньше электрическая емкость мембраны, больше ее поперечное сопротивление (меньше ток утечки) и меньше продольное сопротивление цитоплазмы. Сопротивление единицы длины аксона определяется площадью поперечного сечения аксона, и следовательно с увеличением диаметра (толщины) аксона скорость проведения импульсов возрастает. У кальмара и многих других беспозвоночных для быстрой передачи сигналов (до 100 м/с) в ходе эволюции появились аксоны огромного диаметра (гигантские аксоны - излюбленный объект исследования нейробиологов). Однако у позвоночных столь же высокая скорость проведения сигналов достигается гораздо более экономичным способом. Аксоны их нервных клеток заключены в липопротеидную, миелиновую оболочку, которая по происхождению является плазматической мембраной особых клеток глии, называемых шванновскими клетками. В процессе формирования нервной системы, швановские клетки многократно накручиваются на аксон (до 300 спиральных слоев), при этом цитоплазма почти полностью выдавливается из межмембранного пространства (рис. 13). Изолирующий слой, образуемый миелиновой оболочкой, резко уменьшает емкость мембраны и утечку через нее тока. Диффузия ионов через миелиновую оболочку практически невозможна. Кроме того, в участках аксона, покрытых миелином отсутствуют потенциал-зависимые ионные каналы. Важно отметить, что локальные токи, достаточные для регенерации ПД могут протекать даже через 2-3 последовательно расположенных перехвата Ранвье. Долее частое, чем необходимо для нормального распространения возбуждения, расположение активных узлов служит повышению надежности нервных коммуникаций в организме. В безмиелиновых волокнах ретрансляция ПД должна происходить значительно чаще (см. рис. ). Там генераторы ПД расположены вдоль всей длины волокна в непосредственной близости друг от доруга. Этим обусловлена сравнительно низкая скорость проведения возбуждения по мембранам мышечных волокон. Передача возбуждения в миелизированом волокне происходит скачкообразно, т.е. сальтаторно. Сальтаторное проведение обеспечивает существенную экономию энергии. Так потребление кислорода при нем в 200 раз меньше, чем при непрерывном распространении нервных импульсов по безмиелиновому волокну.  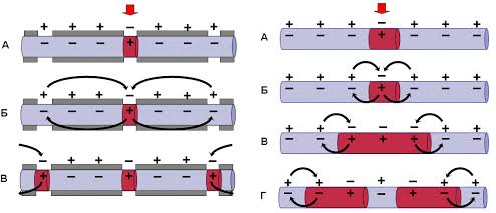 Передача электрических сигналов нервной клеткой основана на изменении мембранного потенциала в результате прохождения относительно небольшого числа ионов через мембранные каналы. Эти ионы перемещаются за счет энергии, большой запас которой создается благодаря работе Na-К насоса, поддерживающего более низкую концентрацию Nа и более высокую концентрацию К внутри клетки по сравнению с наружной средой. В покоящемся нейроне каналы избирательной утечки К делают мембрану более проницаемой для калия, чем для других ионов, и поэтому мембранный потенциал покоя близок к равновесному потенциалу К, составляющему примерно - 70 мВ. Внезапная деполяризация мембраны изменяет ее проницаемость, так как при этом открываются потенциал-зависимые натриевые каналы. Но, если деполяризованное состояние поддерживается, эти каналы вскоре инактивируются. Под влиянием мембранного электрического поля отдельные каналы совершают резкий переход от одной из возможных конформаций к другой. Потенциал действия инициируется тогда, когда под влиянием короткого деполяризующего стимула открывается часть потенциал-зависимых натриевых каналов, что делает мембрану более проницаемой для Nа и еще дальше смещает мембранный потенциал по направлению к равновесному натриевому потенциалу. В результате такой положительной обратной связи открывается еще больше натриевых каналов, и так продолжается до тех пор, пока не возникнет потенциал действия, подчиняющийся закону всё или ничего Потенциал действия быстро исчезает вследствие инактивации натриевых каналов, а во многих нейронах также и открытия потенциал-зависимых калиевых каналов. 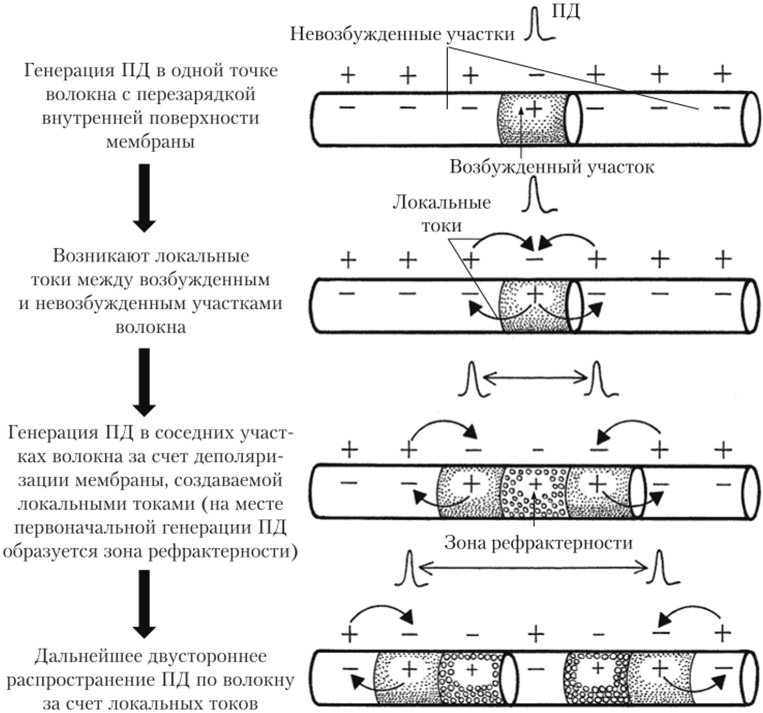 Кабельная теория дендритов (также Кабельная теория Ролла или Модель Ролла) — это множество предположений и результатов, которые относятся к распространению и взаимодействия электрических сигналов в дендритных деревьях. Кабельная теория применяется для описания проведения биоэлектрических потенциалов вдоль цилиндрической клетки. Кабельная теория исходит из того, что нервную, мышечную или иную клетку можно представить в виде отрезка кабеля, помещенного в проводящую среду и имеющего клеточную мембрану, которая играет роль изоляции. Кабельная модель цилиндрической клетки и основанная на этой модели теория расчёта соотношения величин тока и напряжения позволяют экспериментально определять электрические параметры клеточной мембраны и оценивать условия распространения подпороговых электрических импульсов. Понимание электротонических свойств нейрона возникло в XIX в. как результат объединения теории распространения тока в нервных клетках и мышцах с кабельной теорией, которая описывала распространение электрического тока на дальних расстояниях через кабели на дне океана. Поэтому «электротонические свойства» нейронов часто называют «кабельными свойствами». Основы кабельной теории заложил английский физик Уильям Томсон (Лорд Кельвин), который в 1855 представил Королевскому Обществу математический анализ затухания сигнала в трансатлантическом телефонном кабеле между Америкой и Англией, который тогда планировали прокладывать. Подводный кабель схож по своим свойствам с нервным волокном. Он имеет токопроводящие сердцевины, покрытые изолирующей оболочкой, и окружён морской водой. (Так же и нейроны находятся в соляном растворе, который содержит электрически заряженные частицы (ионы), которые генерируют электрический потенциал и двигаются в ответ на потенциалы). Но, поскольку изоляция кабеля не является совершенной, то существует конечное сопротивление утечки через изолятор. Главная количественная разница между кабелем и нейроном в том, что кабельные жилы сделаны из меди, что является намного лучшим проводником, чем соляной раствор нейрона, а также кабельное покрытие является значительно лучшим изолятором, чем мембрана клетки. Поэтому сигналы в кабеле, в отличие от нейрона, могут распространяться на дальние расстояния без существенного затухания. Кабельная теория впервые была математически применена к нервной системе в конце XIX в. для описания электрического тока через нервные волокна. В 1940-х гг. Алан Ходжкин и др. применили кабельную теорию к аксонам беспозвоночных (краба и кальмара), тем самым положив начало математическому описанию и моделированию нейронов. Созданные Роллом модели позволили построить единую математическую теорию дендритных функций, которая в сочетании с математическими моделями генерации синаптических потенциалов и потенциала действия обеспечила целостное теоретическое описание нейронной активности. Нервные и мышечные волокна, будучи генераторами электромагнитной энергии, вместе с тем обладают пассивными электрическими свойствами. Они характеризуют способность живых тканей поглощать энергию внешнего ЭМП, затрачиваемую на их поляризацию. Характеризуя потери электромагнитной энергии в тканях, говорят о декременте (затухании) в них электрического потенциала, которым обладало ЭМП до взаимодействия с ними. Закономерности затухания идентичны для потенциалов, приложенных извне и генерируемых самими живыми тканями. Степень декремента зависит от их сопротивления (R) и емкости (С). В электротехнике R и С называются пассивными свойствами электрических цепей. Пассивные электрические свойства клеток, в первую очередь нервных и мышечных волокон, определяющие декремент потенциала на них, в биофизике принято называть кабельными свойствами, поскольку живую клетку, особенно нервное или мышечное волокно, можно уподобить электрическому кабелю, помещенному в электропроводящую среду. КАБЕЛЬНЫЕ СВОЙСТВА БИОЛОГИЧЕСКИХ МЕМБРАН Допустим, в какой-то точке биомембраны скачком (мгновенно) изменился мембранный потенциал (МП) на величину 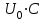 . С течением времени сдвиг потенциала в данной точке угасает. Декремент потенциала (т. е. этого сдвига МП- . С течением времени сдвиг потенциала в данной точке угасает. Декремент потенциала (т. е. этого сдвига МП-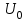 ) имеет экспоненциальный характер: ) имеет экспоненциальный характер: 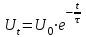 Где 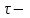 постоянная времени (время, в течение которого падает в е раз, т. е. постоянная времени (время, в течение которого падает в е раз, т. е. 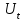 достигает примерно 37 % ). достигает примерно 37 % ).Поскольку 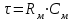 , декремент определяется сопротивлением ( , декремент определяется сопротивлением (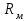 ) и емкостью биомембраны ( ) и емкостью биомембраны (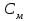 ). Например, в гигантском аксоне кальмара ). Например, в гигантском аксоне кальмара 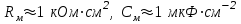 и, следовательно, среднее значение и, следовательно, среднее значение 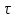 составляет около 1 мс (от 0,7 до 1,6 мс). У миелинизированного аксона лягушки короче (приблизительно 0,006 мс), а упоперечнополосатого мышечного волокна – длительнее (10,3 мс). Различия обусловлены разнообразием пассивных электрических свойств ( составляет около 1 мс (от 0,7 до 1,6 мс). У миелинизированного аксона лягушки короче (приблизительно 0,006 мс), а упоперечнополосатого мышечного волокна – длительнее (10,3 мс). Различия обусловлены разнообразием пассивных электрических свойств ( ) различных плазматических мембран. ) различных плазматических мембран. У 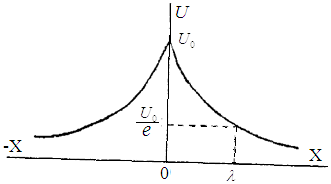 гасание потенциала происходит не только с течением времени в точке его возникновения, но также при распространении потенциала вдоль мембраны по мере удаления от этой точки. Такой декремент является функцией не времени, а расстояния (х): гасание потенциала происходит не только с течением времени в точке его возникновения, но также при распространении потенциала вдоль мембраны по мере удаления от этой точки. Такой декремент является функцией не времени, а расстояния (х):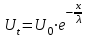 , где , где 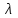 - постоянная длины (расстояние, на котором падает в е раз). - постоянная длины (расстояние, на котором падает в е раз). Рис.1. Декремент электрического потенциала вдоль мембраны: по оси абсцисс- расстояние (х) от точки с максимальным уровнем потенциала ( ); по оси ординат- уровень электрического потенциала (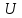 ); - постоянная длины ); - постоянная длины 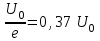 Декремент потенциала вдоль биомембраны изображен на рис.1. Угасание происходит в обе стороны от точки пикового значения сдвига мембранного потенциала. Распределение электрического потенциала на мембране, изображенного на рис.1. устанавливается моментально, так как скорость распространения ЭМП имеет порядок 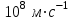 , и сохраняется только на мгновение. , и сохраняется только на мгновение.С течением времени потенциал падает во всех точках волокна в соответствии с уравнением для (понятно, что величина в этом уравнении для каждой точки разная и равна 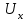 ). ). При длительно существующих сдвигах мембранного потенциала постоянная длины вычисляется по формуле: 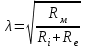 , где , где 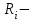 сопротивление цитоплазмы, сопротивление цитоплазмы,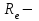 сопротивление межклеточной среды, - удельное сопротивление мембраны. При коротких импульсах (например, таких, как ПД) нужно учитывать также емкостные свойства биомембраны. Чем больше , тем слабее декремент потенциала вдоль мембраны. Так, в гигантском аксоне кальмара около 2,5 мм, а в портняжной мышце лягушки- 0,55-0,80 мм. Следовательно, на плазмолемме мышечного волокна электрический потенциал затухает быстрее. У большинства клеток составляет расстояние, равное приблизительно 20-40 поперечным размерам. сопротивление межклеточной среды, - удельное сопротивление мембраны. При коротких импульсах (например, таких, как ПД) нужно учитывать также емкостные свойства биомембраны. Чем больше , тем слабее декремент потенциала вдоль мембраны. Так, в гигантском аксоне кальмара около 2,5 мм, а в портняжной мышце лягушки- 0,55-0,80 мм. Следовательно, на плазмолемме мышечного волокна электрический потенциал затухает быстрее. У большинства клеток составляет расстояние, равное приблизительно 20-40 поперечным размерам.Декремент потенциала зависит также от крутизны переднего фронта этого потенциала, которая определяется по формуле: 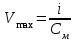 , где i- ионный ток через мембрану , где i- ионный ток через мембрану 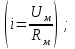 - емкость мембраны. Чем больше - емкость мембраны. Чем больше 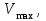 тем слабее декремент потенциала. тем слабее декремент потенциала.Таким образом, и являются основными параметрами кабельных свойств биомембран. Они количественно характеризуют декремент потенциала как во времени, так и в пространстве (вдоль координаты х). Постоянная длины волокна возрастает по мере понижения сопротивления цитоплазмы и уменьшения емкости клеточной мембраны. |