Пол это совокупность морфологических, физиологических, биохимических, поведенческих и других признаков организма, обеспечивающих воспроизводство потомства и передачу наследственной информации следующим поколениям.
 Скачать 282.73 Kb. Скачать 282.73 Kb.
|

|
1.Пол, его определение Пол – это совокупность морфологических, физиологических, биохимических, поведенческих и других признаков организма, обеспечивающих воспроизводство потомства и передачу наследственной информации следующим поколениям. Каждый вид животных и двудомных растений характеризуется примерно равным количеством особей мужского и женского пола. Таким образом, соотношение полов в популяции близко к 1:1. Существуют различные типы определения пола и механизмы определения пола Определе́ние по́ла, или детермина́ция по́ла — биологический процесс, в ходе которого развиваются половые характеристики организма. Большинство организмов имеют два пола. Иногда встречаются также гермафродиты, сочетающие признаки обоих полов. Некоторые виды имеют лишь один пол и представляют собой самок, размножающихся без оплодотворения путём партеногенеза, в ходе которого на свет появляются также исключительно самки. 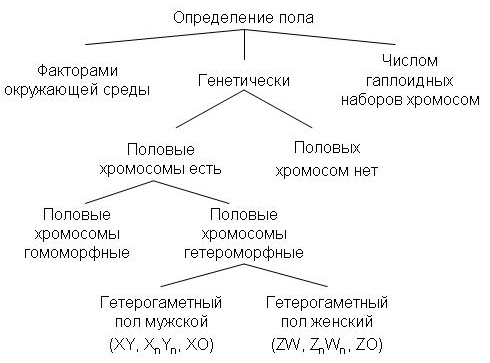 Варианты механизмов определения пола[1] Половое размножение и проявление полового диморфизма широко распространено в различныхтаксономических группах. Для механизмов полоопределения характерно большое разнообразие, что свидетельствует о неоднократности и независимости возникновения пола в различных таксонах[2]. Во многих случаях пол определяется генетически. Генетическая детерминация пола — наиболее распространённый способ определения пола у животных и растений, пол при этом может определяться серией аллелей одного или нескольких аутосомных генов, или детерминация пола может происходить при помощи половых хромосом с пол-определяющими генами (см. Хромосомное определение пола)[2]. При хромосомном определении пола набор половых хромосом у самцов и самок, как правило, разный из-за их гетероморфности, и пол определяется комбинациями половых хромосом: ХY, ZW, X0, Z0. В других случаях пол определяется факторами окружающей среды. Например, у всех крокодилов, некоторых ящериц,черепах, рыб и гаттерии пол зависит от температуры, при которой развивалась особь. У муравьёв, пчёл, оси некоторых других насекомых существует ещё один механизм: пол зависит от числа хромосомных наборов. Гаплоидные самцы развиваются из неоплодотворённых яиц, а диплоидные самки — из оплодотворённых[1]. Некоторые виды не имеют константного пола и могут менять его под действием внешних стимулов. Детали некоторых механизмов определения пола ещё не полностью ясны. Следует отличать детерминацию пола от дифференцировки пола. После детерминации пола по какому-либо из упомянутых выше механизмов, запускается половая дифференцировка. Это запуск, как правило, осуществляется главным геном — половым локусом, вслед за ним по каскадному механизму в процесс включаются остальные гены. Классификация механизмов полоопределения Половая принадлежность организма может определяться на разных этапах относительно момента оплодотворения, в зависимости от этого выделяют 3 типа определения пола:
Прогамное определение полаКак писалось выше, прогамное определение пола происходит до оплодотворения, в процессе формирования яйцеклеток. Это имеет место, например, уколовраток. Они образуют яйцеклетки двух сортов: крупные, с двумя наборами хромосом (диплоидные) и большим объёмом цитоплазмы и мелкие, с одним набором хромосом — гаплоидные. Из гаплоидных неоплодотворённых яиц развиваются гаплоидные самцы, продуцирующие гаплоидные гаметы. Если теперь гаплоидный самец оплодотворит гаплоидное яйцо, то разовьётся самка. Из крупных диплоидных яиц также развиваются самки, но в этом случае они появляются не в результате оплодотворения, а партеногенетически, то есть без оплодотворения. Таким образом, пол особи, развивающейся из диплоидного яйца, определяется ещё на стадии формирования яйцеклетки (на этом этапе закладывается её диплоидность), а пол особи, развивающейся из мелкого яйца, зависит от того, будет оно оплодотворено или нет[4]. Хромосомное определение пола[править | править вики-текст] Y-хромосома человека У растений и животных наиболее распространён хромосомный механизм определения пола. В зависимости от того, какой пол является гетерогаметным, выделяют следующие типы хромосомного определения пола:
У особей гомогаметного пола ядра всех соматических клеток содержат диплоидный набор аутосом и две одинаковыеполовые хромосомы, которые обозначаются как XX (ZZ). Организмы такого пола продуцируют гаметы только одного класса — содержащие по одной X (Z) хромосоме. У особей гетерогаметного пола в каждой соматической клетке, помимо диплоидного набора аутосом, содержатся либо две разнокачественные половые хромосомы, обозначаемые как Х и Y (Z и W), либо только одна — X (Z) (тогда количество хромосом получается нечётным). Соответственно у особей такого пола образуются два класса гамет: либо несущие X/Z-хромосомы и Y/W-хромосомы, либо несущие X/Z-хромосомы и не несущие никаких половых хромосом[5]. У многих видов животных и растений гомогаметен женский пол, а гетерогаметен мужской. К ним относятся млекопитающие[6], некоторые насекомые[7], некоторые рыбы[8] и некоторые растения[9] и др. Гомогаметный мужской пол и гетерогаметный женский имеют птицы, бабочки и некоторые рептилии. XY-определение пола[править | править вики-текст]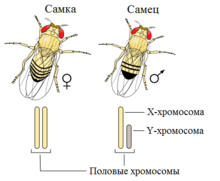 XY-определение пола удрозофилы XY-определение пола является наиболее распространённым; такая система определения пола работает у человека, также как и у подавляющего большинства млекопитающих. В этой системе женские особи имеют две одинаковые половые хромосомы XX, а у мужских имеется две различные половые хромосомы X и Y. Пара половых хромосом XY не похожи друг на друга по форме, размеру и генному составу, что отличает их от пар аутосомных гомологов. Половые хромосомы X и Y носят также название «гоносомы». У некоторых видов, в том числе и человека, на Y-хромосоме имеется ген SRY, определяющий мужское начало. У других, например, плодовой мушки (Drosophila melanogaster) пол зависит от соотношения числа Х-хромосом (Х) и наборов аутосом (А). Если оно равно 1, из неё развивается самка, если 0,5 — самец. При промежуточном соотношении (0,67) развиваются интерсексы — нечто среднее между самцами и самками. При соотношении > 1 развиваются метасамки (сверхсамки), при соотношении < 0,5 — метасамцы (сверхсамцы). И сверхсамцы, и сверхсамки слабы и рано погибают. При этом Y-хромосома роли в определении пола не играет, но она необходима для образования спермы[24]. X0-определение пола[править | править вики-текст] Кариотип горной слепушонки(Ellobius lutescens) 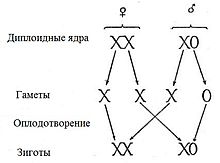 Наследование половых хромосомпри X0-определении пола Эта система представляет собой вариант XY-системы. Самки имеют две копии половой хромосомы (XX), а самцы — только одну (X0). 0 означает отсутствие второй половой хромосомы. В этом случае, как правило, пол определяется количеством генов, экспрессируемых на обеих половых хромосомах. Эта система встречается у некоторых насекомых, в том числе кузнечиков и сверчков из отряда прямокрылые (Orthoptera), а также тараканов (Blattodea). У небольшого числа млекопитающих также отсутствует Y-хромосома. К ним относятся мышевидные грызуны Tokudaia osimensis иTokudaia tokunoshimensis, а также обыкновенная бурозубка (Sorex araneus) из землеройковых (Soricidae). Горная слепушонка (Ellobius lutescens) также имеет форму X0-определения пола, в которой у обоих полов отсутствует вторая половая хромосома[27]. В этом случае механизм определения пола ещё не до конца понятен[35]. ZW-определение пола[править | править вики-текст] Комодский варан (Varanus komodoensis), появившийся на свет в результате партеногенеза Основная статья:ZW-определение пола ZW-определение пола встречается у птиц, рептилий, некоторых насекомых (бабочек) и других организмов. ZW-система обратна XY-системе: самки имеют две различные половые хромосомы (ZW), а самцы — одинаковые (ZZ). У курицы ген DMRT1 является ключевым полообразующим геном[38]. У птиц гены FET1 иASW, имеющиеся в W-хромосоме самок, сходны с геном SRY Y-хромосомы[25]. Однако не у всех организмов пол зависит от наличия W-хромосомы. Например, умолей и бабочек самки имеют кариотип ZW, но встречаются также самки Z0 и ZZW[36]. Кроме того, хотя у самок млекопитающих инактивируется одна из Х-хромосом, у самцов бабочек этого не наблюдается, и они образуют вдвое больше нормального количества ферментов, так как имеют две Z-хромосомы[36]. Поскольку ZW-определение пола широко варьирует, до сих пор неизвестно, как именно большинство видов определяют свой пол. Несмотря на сходства между XY и ZW, эти хромосомы возникли раздельно. В случае куриц их Z-хромосома наиболее похожа на человеческую 9-ю хромосому[39]. Куриная Z-хромосома также предположительно является родственной Х-хромосомам утконоса[40]. Когда ZW-виды, например, комодский варан (Varanus komodoensis), размножаются партеногенетически, рождаются только самцы. Это происходит из-за того, что гаплоидные яйца удваивают свои хромосомы, в результате чего получается ZZ или WW. Из ZZ развиваются самцы, в то время как WW нежизнеспособны и не развиваются далее яйца[41]. Вероятно, исходным для бабочек механизмом определения пола был механизм Z0 самка/ZZ самец[42]. Затем, путем хромосомных перестроек, возникла система определения пола WZ самка/ZZ самец, характерная для 98 % видов бабочек. У тутового шелкопряда (система WZ/ZZ) обнаружен отвечающий за развитие женского пола ген Fem в W-хромосоме. Z0-определение пола[При Z0-системе определения пола мужские особи имеют кариотип ZZ, а женские — Z0. Иными словами, у видов с системой Z0/ZZ определение пола зависит от соотношения числа пар половых хромосом и аутосом. Такое определение пола встречается у некоторых молей[43][44]. Гаплодиплоидность[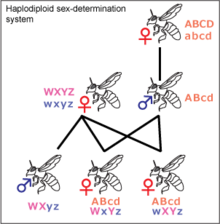 Гаплодиплоидное определение пола Сущность гаплоидиплоидности заключается в том, что генотипы самцов и самок различаются на геномном, а нехромосомном, уровне: гаплоидный организм развивается в самца, а диплоидный — в самку[45]. Гаплодиплоидность встречается у насекомых отряда Перепончатокрылые (Hymenoptera), например, муравьёв и пчёл. Неоплодотворённые яйца развиваются в гаплоидных самцов[45]. Диплоидные особи, развивающиеся из оплодотворённых яиц, как правило, являются самками[45], но могут быть и стерильными самцами. У самцов не может быть отцов и сыновей. Еслипчелиная матка спаривается с одним трутнем, её дочери имеют ¾ общих генов, а не ½, как в XY- и ZW-системах. Предполагается, что это имеет важное значение для развития эусоциальности, поскольку это увеличивает роль кин-отбора, но такая точка зрения оспаривается[46]. Большинство самок перепончатокрылых могут выбирать пол своих потомков, сохраняя полученную сперму в сперматеке и выпуская или не выпуская её в яйцевод. Это позволяет им создавать больше рабочих особей в зависимости от состояния колонии[47]. Эпигамное определение полаТемпературозависимое определение пола У аллигаторов пол потомства определяется температурой гнезда. Помимо генетических, существует много других механизмов определения пола. У некоторых рептилий пол определяется температурой, в которой развивалось яйцо в течение температурочувствительного периода. Это явление носит название температурозависимого определения пола (англ. temperature-dependent sex determination (TSD)). К видам с температурозвисимым определением пола относятся все крокодилы, большинство черепах, некоторые виды ящериц[48], а также гаттерии[49]. Обычно при низких температурах (ниже 27 ºС) из яиц вылупливаются особи одного пола, при высоких (выше 30 ºС) — другого, и только в небольшом промежуточном интервале — особи обоих полов. Так, у черепах при низких температурах появляются только самцы, у ящериц — только самки. Существуют отклонения от этой схемы. У каймановой черепахи при температурах от 20 до 30 ºС преобладают самцы, за пределами этого интервала — самки[48]. Эти температурные пороги обозначаются как порог I и порог II соответственно. Температуры, необходимые для развития каждого пола, называются женскими и мужскими благоприятствующими температурами (англ. the female and the male promoting temperature)[50] . 2.Уровни дифференциации пола в развитии. Пол - понятие комплексное, состоящее из нескольких взаимосвязанных звеньев репродуктивной системы: генетической структуры половой клетки (генетический пол), морфоструктуры гонад (гонадный пол), баланса половых гормонов (гормональный пол), строения половых органов и вторичных половых признаков (соматический пол), психосоциального и психосексуального самоопределения (психический пол), определенной роли в семье и обществе (социальный пол). В конечном итоге пол - это биологически и социально обусловленная роль субъекта в семье и обществе. Формирование пола человека проходит в онтогенезе несколько этапов. I ЭТАП Пол будущего организма предопределяется в момент оплодотворения и зависит от сочетания в зиготе половых хромосом: XX набор соответствует женскому, XY - мужскому полу. С Y-хромосомой связана активность гена-активатора HY-генов, определяющих развитие первичной гонады в мужском направлении. Они запускают синтез HY-антигена и белков-рецепторов к нему, гены которых локализованы в других хромосомах. Другая система генов Y-xpoмосомы обеспечивает развитие придатка яичка, семенных пузырьков, семявыносящего протока, предстательной железы, наружных гениталий в мужском направлении, а также инволюцию мюллеровых производных. В половых клетках первичной гонады (и при XY, и при XX хромосомных наборах) есть рецепторы к HY-антигену, в то время как в соматических клетках, они имеются только при XY-наборе [в структуру соматических рецепторов к HY-антигену входит особый вид р-микроглобулина, тогда как рецепторы половых клеток к HY-антигену (и XY, и XX) не связаны с этим специфическим белком]. Вероятно, этим и объясняется бипотенциальность первичной гонады. II ЭТАП Между 7-й и 10-й неделей внутриутробного развития происходит формирование половых желез в соответствии с набором половых хромосом. III ЭТАП Между 10-й и 12-й неделей эмбриогенеза образуются внутренние гениталии. Функционально полноценные тестикулы в этот период выделяют особый пептидный гормон [Josso, 1984], вызывающий рассасывание мюллеровских производных. При отсутствии тестикулов или при их патологии с нарушением продукции антимиллерова гормона развиваются внутренние женские половые органы (матка, трубы, влагалище) даже у эмбриона с генетическим мужским полом (46, XY). IV ЭТАП Между 12-й и 20-й неделей эмбриогенеза идет формирование наружных гениталий. Определяющую роль в мужском развитии на этом этапе играют андрогены (независимо от их источника) - тестикулярные, надпочечниковые, поступающие из материнского организма (при наличии у матери андрогенпродуцирующих опухолей или в связи с приемом андрогенных препаратов). При отсутствии андрогенов и при нарушении рецепторной чувствительности к ним наружные гениталии формируются по женскому (<нейтральному>) типу даже при наличии 46,XY кариотипа и нормальной функции эмбриональных тестикулов. Возможно и развитие промежуточных вариантов (неполной маскулинизации). V ЭТАП Опускание тестикулов в мошонку. Происходит между 20-й и 30-й неделей эмбриогенеза. Механизм, вызывающий или нарушающий продвижение тестикулов, до конца не ясен. Однако несомненно, что и тестостерон, и гонадотропины причастны к этому процессу. VI ЭТАП Половой дифференцировки происходит уже в пубертатном возрасте, когда окончательно формируются связи в системе гипоталамус - гипофиз - гонады, активируется гормональная и генеративная функция гонад и закрепляется социально-половое самосознание, определяющее роль субъекта в семье и обществе. http://www.medrostov.ru Развитие признаков пола, как и любых других признаков организма, определяется генотипом и факторами внешней среды. Так как организмы генетически бисексуальны, процесс дифференциации пола оказывается сложным. Бисексуальная основа организма в принципе позволяет изменять направление его развития, т. е. переопределять пол в онтогенезе. Вслед за определением пола следует дифференциация, т. е. развитие половых различий: формирование воспроизводительной системы, а также физиологического и биохимического механизмов, обеспечивающих скрещивание. В ходе дифференциации пола идет развитие одного из слоев гонады и подавление другого. У мужского пола быстрее развивается медуллярная ткань, которая подавляет деятельность кортикального слоя, в результате гонады превращаются в семенники. У женского пола ускоряется развитие кортикального слоя, в силу чего подавляется формирование медуллярного слоя и гонады превращаются в яичники. В соответствии с этими преобразованиями дифференцируются и половые пути, которые тоже закладываются одинаковыми у особей обоих полов. Процесс дифференциации пола у многих животных обусловлен гормонами, которые выделяются не только эндокринными железами, но кортикальным и медуллярным слоями полового зачатка, а в последующем — и половыми железами. Эти вещества отличаются по характеру своего действия и по времени их выработки. У особей мужского пола преобладает «мужское вещество» — медулларин, у женского — кортикальный гормон. Дальнейшая дифференциация пола, особенно развитие вторичных половых признаков, также идет под влиянием различных гормонов. Уровень гормональной секреции контролируется генами, точнее, их балансом. Преобладание генов, определяющих мужской пол, в общем балансе приводит к повышению активности мужских гормонов и к дифференциации мужского пола, обратное соотношение генов — к развитию женского пола. Смена активности гормональной секреции то одного, то другого пола в онтогенезе приводит к развитию интерсексуальных форм. Уровни дифференцировки пола в развитии: 1. Генотипическая (хромосомный) 2. Определение пола на уровне гонад 3. Фенотипическое – различие в строении внутренних и внешних развитии признака. 4. Психологический 5. Социальный (по паспорту) 3.Генотипическое определение пола Пол организмов, размножающихся половым способом, определяется парой особах половых хромосом, тогда как остальные хромосомы называют аутосомами. Пол, особи которого имеют одинаковые половые хромосомы, называется гомогаметным. У особей гомогаметного пола при образовании гамет все они будут одинаковыми по половым хромосомам. Пол, у особей которого половые хромосомы разные, называется гетеро- гаметным. Соответственно и половые клетки этих особей будут двух типов, причемобразовываться они будут в равном соотношении. У половых хромосом нет принципиальных отличий от аутосом: они, как и аутосомы, несут гены, которые контролируют развитие тех или иных признаков. Половые хромосомы принято обозначать буквами. У видов с гетерогаметным мужским полом одинаковые по размеру и форме половые хромосомы обозначают буквой X, а хромосому, имеющуюся только у самцов, - буквой Y. Такой тип наследования пола встречается у многих червей, моллюсков и членистоногих, а из позвоночных - у млекопитающих и человека. В хромосомном комплексе самок этих видов две X-хромосомы, а у самцов - X-хромосома и Y-хромосома. У видов с гетерогаметным женским полом хромосому, имеющуюся как у самок, так и у самців обозначают буквой Z, а имеющуюся только у самок - буквой W. Самки этих видов имеют хромосомный комплекс с половыми хромосомами ZW, а самцы - с половыми хромосомами ZZ. Такой тип наследования пола имеет место, в частности, среди растений - у клубники, среди насекомых - у бабочек, а среди позвоночных - у рыб, пресмыкающихся и птиц. Медоносная плела относится к видам, у которых гомогаметным яв- ляется женский пол, но пчелиная матка откладывает два типа яиц - оплодотворенные и неоплодотворенные. Из оплодотворенных яйцеклеток (генотип XX) развиваются самки (матка и рабочие пчелы), а из неоплодотворенных (одна X- хромосома, генотип X0) - самцы (трутни). В связи с этим организмы, образовавшиеся при обыкновенном половом размножении, - диплоидные, а самцы, развивающиеся в результате партеногенеза, - гаплоидные. В процессе индивидуального развития в клетках тела трутней восстанавливается диплоидный хромосомный набор, тогда как в половых клетках гаплоидное число хромосом сохраняется и при образовании сперматозоидов не уменьшается. Аналогичное явление встречается и у некоторых других насекомых. Х- хромосома намного длиннее, чем Y-хромосома, и содержит некоторые гены, которых нет в Y-хромосоме, в том числе и гены, кодирующие не связанные с проявлением пола белки. Однако продукты таких генов не образуются в женском организме в двойном количестве по сравнению с мужским организмом, потому что одна из X-хромосом в каждой клетке женского организма неактивна, причем инактивация происходит на ранних етапах эмбриогенеза. Будет ли в данной отдельной клетке инактивироваться материнская или отцовская X-хромосома - дело случая, но далее во всех поколениях клеток, происходящих от данной клетки, неактивной будет одна и та же хромосома, поэтому женский організм является мозаичным по признакам, определяемым X-хромосомой. Механизм инактивации X-хромосомы неизвестен. 4.Первичное, вторичное, третичное соотношение полов. Соотношение полов (СП) является важной характеристикой раздельнополой популяции. Обычно его выражают количеством самцов, приходящихся на 100 самок, долей мужских особей или в процентах. В зависимости от стадии онтогенеза различают первичное, вторичное и третичное соотношение полов. Первичное—это соотношение полов в зиготах после оплодотворения; вторичное—соотношение полов при рождении и, наконец, третичное—соотношение полов зрелых, способных размножаться особей популяции. В настоящее время доказано и общепринято, что у большинства видов животных и растений основным механизмом, определяющим пол, является хромосомный. Поскольку в ходе гаметогенеза гаметы, содержащие Х и Y-хромосомы, производятся в равном числе, считалось, что этот механизм обеспечивает примерно равные количества полов при зачатии. Согласно новой концепции третичное соотношение полов устанавливает отношение между тенденциями наследственности и изменчивости, т. е. определяет эволюционную пластичность вида. Поскольку на разных этапах эволюции, а также в различных условиях среды требуется разная эволюционная пластичность популяции, то для каждого из них существует свое определенное оптимальное значение третичного соотношения полов, не обязательно равное 1:1. Вторичное соотношение полов также является не константой, а переменной величиной, тесно связанной с условиями среды. В стабильных, оптимальных условиях, когда не требуется много мужских особей, вторичное соотношение полов должно понижаться. В изменчивых, экстремальных условиях, когда требуется много мужских особей, наоборот, оно должно повышаться 5.Теория генного баланса К.Бриджеса, ее суть и значение для биологии и медицины. Балансовая теория пола К. Бриджеса При изучении наследования пола у мухи дрозофилы было установлено, что самцы могут иметь разные наборы половых хромосом XY и ХО (последние имеют все признаки мужского пола, но стерильные, так как в Y-хромосоме содержатся гены, необходимые для нормального течения сперматогенеза). Из этого был сделан вывод, что Y-хромосома у мухи дрозофилы не имеет существенного значения для определения мужского пола. Затем были получены особи с разнообразными сочетаниями числа Х-хромосом и наборов аутосом (А) и изучен их пол: 2Х: 2А — нормальные самки; 1Х: 2А — нормальные самцы; ЗХ: 2А — сверхсамки; гипертрофированы признаки женского пола, бесплодны; 1Х: ЗА — сверхсамцы; гипертрофированы признаки мужского пола, бесплодны; 2X: ЗА — интерсексы; имеют признаки обоих полов, бесплодны. Пол в данном случае определяется не половыми хромосомами, а отношением (балансом) числа Х-хромосом и количества наборов аутосом. Если это отношение 1: 1— развиваются нормальные самки, если отношение 1: 2— развиваются нормальные самцы. Чем больше в кариотипе Х-хромосом, тем более выражены признаки женского пола; чем больше наборов аутосом, тем резче проявляются признаки мужского пола. При отношении 1: 1,5 (2Х: ЗА) развиваются признаки обоих полов. При нарушении течения митоза могут образовываться особи —гинандроморфы. Содержание половых хромосом в разных клетках таких особей разное (мозаичность). Например, у мухи дрозофилы в одних клетках содержатся две Х-хромосомы, а в других — ХО, в связи с чем разные части тела могут иметь соответствующие признаки пола. У человека могут быть разные случаи мозаицизма: ХХ/ХХХ, XY/XXY, ХО/ХХХ, ХО/XXY и др. Если процент мозаичных клеток велик, возможны морфо-физиологические проявления. 6.Особенности наследования сцепленных с полом признаков.Примеры. Признаки, зависимые от пола и ограниченные полом.Характеристика, примеры. Гены могут находиться на половых хромосомах, в этом случае говорят, что они сцеплены с полом. Наследование, сцепленное с полом, имеет некоторые важные особенности. Дело в том, что У-хромосома несет гораздо меньше генов, чем Х-хромосома. Это обстоятельство приводит к тому, что для многих генов наХ-хромо-соме нет соответствующих аллелей на У-хромосоме. В результате если у мужчины наХ-хромосоме оказывается рецессивный аллель, то он проявится в фенотипе. Например, имеется наследственная форма гемофилии - болезни, связанной с нарушением нормальной свертываемости крови. При этих нарушениях у больного возникают длительные кровотечения даже при незначительном повреждении кровеносных сосудов. Существуют две формы гемофилии - А и В, я обе определяются рецессивными генами, локализованными в Х-хромосоме. На рис.2 показано наследование гемофилии. Схематически представлены половые хромосомы родителей. На Х-хромосомах обозначены аллели рецессивного аллеля гемофилии а. Нормальный аллель доминирует - обозначен как А. Если в брак со здоровым мужчиной вступает здоровая женщина, являющаяся носительницей гена гемофилии, то с одинаковой вероятностью (в 25% случаев) могут родиться здоровая дочь, здоровая дочь - носительница гена гемофилии, здоровый сын, больной сын. Таким образом, носительницами гена гемофилии являются женщины, а болеют ею мужчины. Однако если мужчина-гемофилик вступит в брак со здоровой женщиной, то все его сыновья будут обязательно здоровы (потому что они получат от отца 7-хромосому). Дочери будут тоже здоровыми, но обязательно будут носительницами гена гемофилии. Теоретически возможна гемофилия и у женщины, но такая вероятность очень невысока, так как для этого необходимо вступление в брак больного-гемофилика с женщиной - носительницей гена гемофилии (и даже в этом случае вероятность рождения больной девочки будет только 0,25). Из-за низкой частоты встречаемости гена гемофилии и того, что больные гемофилией часто умирают до брачного возраста, такие случаи практически не отмечаются. Итак, если рецессивный ген сцеплен с А-хромосомой, то он гораздо чаще проявляется в фенотипе у мужчин, чем у женщин. Среди других генов, сцепленных с полом, стоит упомянуть гены, связанные с цветовой слепотой. Признаки, зависимые от пола и ограниченные полом. Признаки, расщепление по которым при скрещивании связано с полом, называют сцепленнымис полом. Эти признаки обусловливаются генами, локализованными в половых хромосомах. Установлено, что наследование их зависит в основном от Х-хромосомы. Y-хромосома имеет небольшие размеры, состоит преимущественно из гетерохроматина и является генетически инертной, за исключением, возможно, некоторых генов, контролирующих воспроизводительную функцию и признаки пола. У самцов млекопитающих гены, локализованные в Х-хромосоме, не имеют доминантных или рецессивных партнеров (аллелей) на Y-хромо-соме. Рецессивные гены у них проявляют свое действие уже в одинарной дозе (гемизиготном состоянии) по типу доминантного. Признаки человека, наследование которых каким-то образом связано с полом, делятся на несколько категорий: Признаки, ограниченные полом. Их развитие обусловлено генами, расположенными в аутосомах обоих полов, но проявляются только у одного пола. Например, гены, определяющие ширину таза женщины, локализованы в аутосомах, унаследуются и от отца и от матери, но проявляются только у женщин. То же самое касается возраста полового дозревания девушек. Среди мужских признаков, ограниченных полом - количество и распределение волосяного покрова на теле Признаки, контролируемые полом или зависимые от пола. Развитие соматических признаков обусловено генами, расположенными в аутосомах, проявляются они у мужчин и женщин, но по-разному: Например, у мужчин раннее облысение— признак доминантен, проявляется как у доминантных гомозигот (АА), так и у гетерозигот (Аа). У женщин этот признак рецессивен, он проявляется только у рецессивных гомозигот (aа). Поэтому лысых мужчин значительно больше, чем женщин. Другим примером может быть подагра, у мужчин ее пенетрантность выше: 80% против 12% у женщин. То есть, чаще подагрой болеют мужчины. Экспрессивность признаков, контролируемых полом,обусловленаполовыми гормонами. Например, тип певческого голоса (бас, баритон, тенор, сопрано, меццо-сопрано и альт) контролируется половой конституцией. Начиная с периода полового созревания, признак находится под влиянием половых гормонов 8.Нарушение расхождения хромосом при гаметогенезе. Анеуплоиди́я (греч. an + eu + ploos + eidos — отрицательная приставка + вполне + кратный + вид) — наследственное изменение, при котором число хромосом в клетках не кратно основному набору. Может выражаться, например, в наличии добавочной хромосомы (n + 1, 2n + 1 и т. п.) или в нехватке какой-либо хромосомы (n — 1, 2n — 1 и т. п.). Анеуплоидия может возникнуть, если в анафазе I мейоза гомологичные хромосомы одной или нескольких пар не разойдутся. В этом случае оба члена пары направляются к одному и тому же полюсу клетки, и тогда мейоз приводит к образованию гамет, содержащих на одну или несколько хромосом больше или меньше, чем в норме. Это явление известно под названием нерасхождение. Когда гамета с недостающей или лишней хромосомой сливается с нормальной гаплоидной гаметой, образуется зигота с нечетным числом хромосом: вместо каких-либо двух гомологов в такой зиготе их может быть три или только один. Зигота, в которой количество аутосом меньше нормального диплоидного, обычно не развивается, но зиготы с лишними хромосомами иногда способны к развитию. Однако из таких зигот в большинстве случаев развиваются особи с резко выраженными аномалиями. Формы анеуплоидии Моносомия Моносомия — это наличие всего одной из пары гомологичных хромосом. Примером моносомии у человека является синдром Тернера, выражающийся в наличии всего одной половой (X) хромосомы. Генотип такого человека X0, пол — женский. У таких женщин отсутствуют обычные вторичные половые признаки, характерен низкий рост и сближенные соски. Встречаемость среди населения Западной Европы составляет 0,03 %. Подробнее читайте в статье синдром Шерешевского-Тернера. В случае обширной делеции в какой-либо хромосоме иногда говорят о частичной моносомии, например синдром кошачьего крика. Трисомия Трисомия — это наличие трёх гомологичных хромосом вместо пары в норме. Наиболее часто встречающейся у человека является трисомия по 16-й хромосоме (более одного процента случаев беременности). Однако следствием этой трисомии является спонтанный выкидыш в первом триместре. Схематическое изображение кариотипа мужчины, страдающего синдромом Дауна. Нерасхождение хромосом G21 в одной из гамет привело к трисомии по этой хромосоме. Среди новорождённых наиболее распространена трисомия по 21-й хромосоме, или синдром Дауна (2n + 1 = 47). Эта аномалия, названая так по имени врача, впервые описавшего её в 1866 г., вызывается нерасхождением хромосом 21. К числу её симптомов относятся задержка умственного развития, пониженная сопротивляемость болезням, врождённые сердечные аномалии, короткое коренастое туловище и толстая шея, а также характерные складки кожи над внутренними углами глаз, что создаёт сходство с представителями монголоидной расы. Другие случаи нерасхождения аутосом: 1. Трисомия 18 (синдром Эдвардса) 2. Трисомия 13 (синдром Патау) 3. Трисомия 16 выкидыш 4. Трисомия 9 5. Трисомия 8 (синдром Варкани) Синдром Дауна и сходные хромосомальные аномалии чаще встречаются у детей, рождённых немолодыми женщинами. Точная причина этого неизвестна, но, по-видимому, она как-то связана с возрастом яйцеклеток матери. Случаи нерасхождения половых хромосом: 1. XXX (женщины внешне нормальны, плодовиты, но отмечается умственная отсталость) 2. XXY, Синдром Клайнфельтера (мужчины, обладающие некоторыми вторичными женскими половыми признаками; бесплодны; яичники развиты слабо, волос на лице мало, иногда развиваются молочные железы; обычно низкий уровень умственного развития) 3. XYY (мужчины высокого роста с различным уровнем умственного развития;) Тетрасомия и пентасомия Тетрасомия (4 гомологичные хромосомы вместо пары в диплоидном наборе) и пентасомия (5 вместо 2-х) встречаются чрезвычайно редко. Примерами тетрасомии и пентасомии у человека могут служить кариотипы XXXX, XXYY, XXXY, XYYY, XXXXX, XXXXY, XXXYY, XYYYY и XXYYY. |