|
Пособие по биологии. Пособие по биологии для поступающих в вузы Авторы Пименов А. В., Гончаров О. В
Онтогенез, или индивидуальное развитие — совокупность взаимосвязанных событий, закономерно совершающихся в процессе осуществления организмом жизненного цикла от момента образования зиготы до смерти. Изучение вопросов, связанных с индивидуальным развитием организмов, занимается эмбриология, основоположником которой считается академик Российской Академии К.М.Бэр. Основы учения об индивидуальном развитии организмов были изложены в его труде "История развития животных", опубликованном в 1828 г.
Индивидуальное развитие заключается в реализации организмом наследственной информации, полученной им от родителей.
Представители каждого вида организмов проходят определенные стадии развития от зиготы одного поколения до зиготы следующего. Такую последовательность стадий развития называют жизненным циклом. Жизненные циклы отличаются большим разнообразием, нередко связаны с чередованием поколений, различных типов размножения, с разными вариантами редукционного деления у растений, одноклеточных и многоклеточных животных, полиморфизмом особей.
Онтогенез — это непрерывный процесс развития особи, но для удобств изучения его делят на определенные периоды и стадии (у многоклеточных животных, размножающихся половым способом):
эмбриональный — от образования зиготы до рождения или же выхода из яйцевых оболочек, который состоит из ряда стадий:
одноклеточная (зигота);
дробление;
гаструляция;
гисто- и органогенез;
постэмбриональный — от выхода из яйцевых оболочек или рождения до смерти организма.
Гаметогенез Гаметогенез — это процесс развития половых клеток — гамет (рис. 308). Предшественники гамет (гаметоциты) образуются на ранних стадиях развития зародыша за пределами половых желез, а затем мигрируют в них. На стадии гаметоцитов клетки, как правило, неотличимы. Различия появляются лишь после их проникновения в половые железы. Гаметоциты диплоидны.
Этапы гаметогенеза Процесс образования сперматозоидов называется сперматогенезом, а образование яйцеклеток — оогенезом. В половых железах различают три разных участка (или зоны):
зона размножения;
зона роста;
зона созревания половых клеток.
Сперматогенез и оогенез включают 3 одинаковые фазы:
фаза размножения;
фаза роста;
фаза созревания (деления).
В сперматогенезе имеется еще одна фаза — фаза формирования.
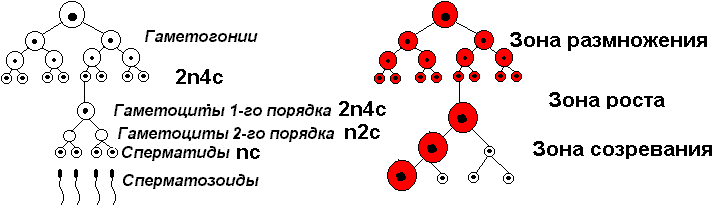
Рис. 308. Основные этапы гаметогенеза.
Фаза размножения Диплоидные клетки многократно делятся митозом. Количество клеток в гонадах растет. Их называют оогонии и сперматогонии. Набор хромосом 2n.
Фаза роста Сущность этой фазы — рост сперматогоний и оогоний, кроме того, в эту фазу происходит репликация ДНК, каждая хромосома становится двухроматидной (2n 4с). Образовавшиеся клетки называются ооциты 1-го порядка и сперматоциты 1-го порядка.
Фаза созревания Сущность фазы — мейоз. В первое мейотическое деление вступают гаметоциты 1-го порядка. В результате первого мейотического деления образуются гаметоциты 2-го порядка (набор хромосом n 2с), которые вступают во второе мейотическое деление, и образуются клетки с гаплоидным набором хромосом (n c). Оогенез на этом этапе практически заканчивается, а сперматогенез включает еще одну фазу, во время которой сперматозоиды приобретают свою специфическую структуру.
В процессе гаметогенеза из диплоидных клеток образуются гаплоидные — гаметы. Происходит это благодаря мейозу. Таким образом, мейоз — основной этап формирования половых клеток.
Сперматогенез Во время периода размножения диплоидные сперматогенные клетки делятся митотически, в результате чего образуется множество более мелких клеток, называемых сперматогониями. Часть образовавшихся сперматогониев может подвергаться повторным митотическим делениям, в результате чего образуются такие же клетки сперматогонии. Другая часть — прекращает делиться и увеличивается в размерах, вступая в следующий период сперматогенеза — период роста. Увеличившиеся в размерах сперматогонии называются сперматоцитами 1-го порядка. Период созревания начинается тогда, когда сперматоцит 1-го порядка подвергается первому мейотическому делению, в результате чего образуются два сперматоцита 2-го порядка. Затем эти вновь образовавшиеся клетки делятся (второе мейотическое деление), и в результате образуются гаплоидные сперматиды. Таким образом, из одного сперматоцита 1-го порядка возникают четыре гаплоидных сперматиды. Период формирования сперматозоидов характеризуется тем, что первично шаровидные сперматиды подвергаются ряду сложных преобразований, в результате которых образуются сперматозоиды. Процесс превращения сперматид в сперматозоиды называется спермиогенезом. В нем участвуют все элементы ядра и цитоплазмы. Ядро сперматид уплотняется вследствие гиперспирализации хромосом, которые становятся генетически инертными. Аппарат Гольджи перемещается к одному из полюсов ядра и образует акросому. Центриоли занимают место у противоположного полюса ядра. Одна из них принимает участие в образовании жгутика. У основания жгутика в виде спирального чехла концентрируются митохондрии. Почти вся цитоплазма сперматиды отторгается.
Оогенез Все периоды развития яйцеклеток осуществляются у животных в яичниках. В отличие от образования сперматозоидов, которое происходит только после достижения половой зрелости (в частности, у позвоночных животных), процесс образования яйцеклеток начинается еще у зародыша. Период размножения полностью осуществляется на зародышевой стадии развития и заканчивается к моменту рождения (у млекопитающих и человека). Он характеризуется тем, что в результате простых мейотических делений первичных половых клеток (оогенных клеток) образуются оогонии, которые снова подвергаются митотическому делению. Дочерние клетки, возникшие в результате деления оогоний, называются ооцитами 1-го порядка. Их возникновение указывает на переход оогенеза в следующую фазу — период роста.
Ооциты увеличиваются в размерах и вступают в профазу I. Увеличение размеров ооцитов связано с тем, что в цитоплазме происходит накопление ряда питательных веществ (белков, жиров, углеводов) и пигментов — образуется желток. Затем ооциты 1-го порядка вступают в период созревания. В результате первого мейотического деления возникают две дочерние клетки. Одна из них, относительно мелкая, называемая первым полярным тельцем, не является функциональной, а другая, более крупная (ооцит 2-го порядка), подвергается дальнейшим преобразованиям.
Второе деление мейоза осуществляется до стадии метафазы II и продолжится только после того, как ооцит 2-го порядка вступит во взаимодействие со сперматозоидом, и произойдет оплодотворение. Таким образом, из яичника выходит, строго говоря, не яйцеклетка, а ооцит 2-го порядка. Лишь после оплодотворения он делится, в результате чего возникает яйцеклетка (или яйцо) и второе полярное тельце. Однако традиционно для удобства яйцеклеткой называют ооцит 2-го порядка, готовый к взаимодействию со сперматозоидом. Таким образом, в результате оогенеза образуется одна нормальная яйцеклетка и три полярных тельца.
Гаметы Гаметы — это половые клетки, при слиянии которых образуется зигота, дающая начало новому организму. Они представляют собой высокоспециализированные клетки, участвующие в осуществлении процессов, связанных с половым размножением. Гаметы имеют ряд особенностей, отличающих их от соматических клеток:
хромосомный набор соматических клеток (у большинства организмов) — диплоидный (2n 2с), а гамет — гаплоидный (n с);
гаметы не делятся;
гаметы, особенно яйцеклетки, более крупные, чем соматические клетки;
яйцеклетка содержит много питательных веществ, сперматозоид — мало (практически отсутствуют);
гаметы имеют измененное ядерно-цитоплазматическое соотношение по сравнению с соматическими клетками (в яйцеклетке ядро занимает значительно больший объем, чем цитоплазма, в сперматозоиде — наоборот, причем ядро имеет такие же размеры, что и яйцеклетка).
Активная роль в оплодотворении принадлежит сперматозоиду. Как правило, он имеет малые размеры и подвижен (у животных). Яйцеклетка не только приносит в зиготу свой набор хромосом, но и обеспечивает ранние стадии развития зародыша. Поэтому она имеет крупные размеры и, как правило, содержит большой запас питательных веществ.
Организация яйцеклеток животных Яйцеклетка человека была открыта в 1821 году К.М.Бэром. Окончательное созревание яйцеклетки происходит уже после оплодотворения, поэтому фактически зрелой яйцеклетки не существует.
Размер яйцеклеток колеблется в широких пределах — от нескольких десятков микрометров до нескольких сантиметров (яйцеклетка человека — около 100 мкм, яйцо страуса, имеющее длину со скорлупой порядка 155 мм — тоже яйцеклетка). Форма ее обычно округлая или слегка сплюснутая. Принципиальных различий в строении яйцеклетки и соматических клеток не существует: они имеют ядро, цитоплазму с органоидами и оболочку (рис. 309). Вместе с тем, яйцеклетка имеет ряд особенностей, отличающих ее от соматических клеток. К ним относятся:
наличие ряда оболочек, располагающихся поверх плазматической мембраны;
наличие в ее цитоплазме более или менее большого количества запасных питательных веществ.
Оболочки У большинства животных яйцеклетки имеют дополнительные оболочки, располагающиеся поверх цитоплазматической мембраны. В зависимости от происхождения различают:
П
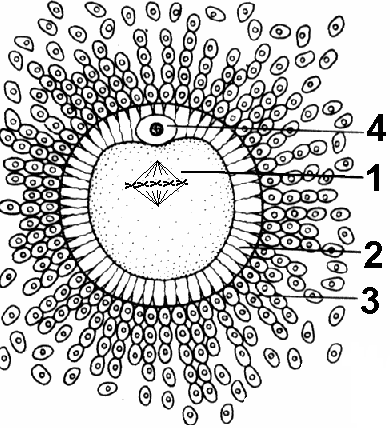
Рис. 309. Яйцеклетка млекопитающих:
1 — пронуклеус на стадии метафазы 2; 2 — блестящая оболочка; 3 — лучистая оболочка; 4 — первое полярное тельце.
ервичные оболочки, возникающие в результате выделения ооцитом и, возможно, фолликулярными клетками веществ, образующих слой, контактирующий с наружной цитоплазматической мембраной яйцеклетки. Выполняют защитную функцию. У некоторых — обеспечивают видовую специфичность проникновения сперматозоида, то есть не позволяет сперматозоидам других видов проникать в яйцеклетку. У млекопитающих эта оболочка называется блестящей.
Вторичные оболочки, образованные выделениями фолликулярных клеток яичника. Имеются не у всех яиц. Вторичная оболочка яиц многих насекомых, например, содержит канал — микропиле, через который сперматозоид проникает в яйцеклетку.
Третичные оболочки, образующиеся за счет деятельности специальных желез яйцеводов. Например, у птиц происходит образование белковой, подскорлуповой пергаментной, скорлуповой и надскорлуповой оболочек.
Вторичные и третичные оболочки, как правило, образуются у яйцеклеток животных, зародыши которых развиваются во внешней среде. Их строение соответствует условиям среды.
Поскольку у млекопитающих наблюдается внутриутробное развитие, их яйцеклетки имеют только первичную оболочку, поверх которой располагается лучистый венец — слой фолликулярных клеток, доставляющих к яйцеклетке питательные вещества.
Питательные вещества яйцеклетки В яйцеклетках происходит накопление запаса питательных веществ, которые называют желтком. Он содержит белки, жиры, углеводы, РНК, минеральные вещества, причем основную его массу составляют липопротеиды и гликопротеиды. Желток содержится в цитоплазме обычно в виде желточных гранул. Количество питательных веществ, накапливаемых в яйцеклетке, зависит от условий, в которых происходит развитие зародыша. Так, если развитие яйцеклетки происходит вне организма матери и приводит к формированию крупных животных, то желток может составлять бо-
л

Рис 310. Типы яйцеклеток хордовых животных:
1 — алецитальная; 2 — изолецитальная; 3 — умеренно телолецитальная; 4 — резко телолецитальная.
ее 95% объема яйцеклетки. Яйцеклетки млекопитающих, развивающиеся внутри тела матери, содержат малое количество желтка — менее 5%, так как питательные вещества, необходимые для развития эмбрионы получают от матери.
В зависимости от количества желтка, содержащегося в яйцеклетках, различают (рис. 310):
алецитальные яйца — яйца, не содержащие желтка или имеющие незначительное количество желточных включе-
ний (млекопитающие, плоские черви);
изолецитальные яйца — яйца с равномерно распределенным желтком (ланцетник, морской еж);
умеренно телолецитальные яйца — яйца с неравномерным распределением желтка (рыбы, земноводные);
резко телолецитальные яйца — яйца, в которых желток занимает большую часть, и лишь небольшой участок цитоплазмы на анимальном полюсе свободен от него (птицы).
В связи с накоплением питательных веществ, у яйцеклеток появляется полярность. Противоположные полюсы называются вегетативным и анимальным. Поляризация у разных животных выражена неодинаково и зависит от количества и распределения желтка.
Поляризация проявляется в том, что происходит изменение местоположения ядра в клетке (оно смещается в сторону анимального полюса), а также в особенностях распределения цитоплазматических включений (во многих яйцах количества желтка возрастает от анимального к вегетативному полюсу).
Организация сперматозоидов Сперматозоид открыт в 1617 году учеником Гука. Он обеспечивает встречу с яйцеклеткой, приносит в нее свою часть генетической информации, стимулирует развитие зиготы. Длина сперматозоида человека 50-60 мкм. Функции сперматозоида определяют и его строение. Сперматозоид млекопитающих имеет форму длинной нити (рис. 311).
Головка Самая крупная часть сперматозоида, образованная ядром, сильно уплотненным в результате гиперспирализации хромосом. Ядро окружено тонким слоем цитоплазмы. На переднем конце головки расположена акросома — часть цитоплазмы с видоизмененным аппаратом
Г
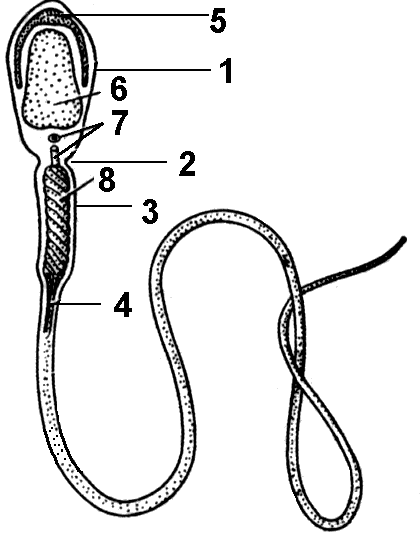
Рис. 311. Строение сперматозоида:
1 — головка; 2 — шейка; 3 — средняя часть; 4 — (жгутик); 5 — акросома; 6 — ядро; 7 — центриоли; 8 —митохондрии.
ольджи. Она вырабатывает фермент гиалуронидазу, который способствует растворению оболочек яйцеклетки и проникновению в нее сперматозоида.
Шейка
В месте перехода головки в среднюю часть образуется перехват — шейка сперматозоида, в которой расположены две центриоли.
Средняя часть
За шейкой располагается средняя часть сперматозоида, представляющая собой скопление митохондрий.
Хвост
Хвост имеет типичное для всех жгутиков эукариот строение и является органоидом движения сперматозоида. Энергию для движения поставляет гидролиз АТФ, происходящий в митохондриях средней части сперматозоида.
Оплодотворение Оплодотворение — совокупность процессов, приводящих к слиянию мужских и женских гамет (сингамия), объединению их ядер (кариогамия) и образованию зиготы, которая дает начало новому организму.
В процессе оплодотворения происходит:
активация яйцеклетки;
восстановление диплоидного набора хромосом;
определение пола будущего организма;
объединение наследственных свойств родительских организмов и возникновение у потомков новых комбинаций наследственных факторов.
Осеменение Как правило, оплодотворению предшествует осеменение. Под осеменением понимают сближение гамет.
Различают два типа осеменения:
наружное, при котором встреча сперматозоидов и яйцеклеток происходит во внешней (чаще всего водной) среде (рыбы);
внутреннее, при котором встреча сперматозоидов и яйцеклеток происходит в половых путях самки (пресмыкающиеся, птицы, млекопитающие).
Проникновение спермия в яйцо Собственно процесс оплодотворения начинается в момент контакта сперматозоида и яйцеклетки. Он начинается с так называемой акросомальной реакции. В момент контакта сперматозоида и яйцеклетки плазматическая мембрана акросомального выроста и прилежащая к ней часть мембраны акросомального пузырька растворяюся, фермент гиалуронидаза и другие биологически активные вещества, содержащиеся в акросоме, выделяются наружу и растворяют участок яйцевой оболочки. Чаще всего сперматозоид полностью втягивается в яйцо, иногда жгутик остается снаружи и отбрасывается. С момента проникновения сперматозоида в яйцо гаметы перестают существовать, так как образуют единую клетку — зиготу.
Слияние генетического материала спермия и яйца Ядро сперматозоида набухает, его хроматин разрыхляется, ядерная оболочка растворяется, и он превращается в мужской пронуклеус. Это происходит одновременно с завершением второго деления мейоза ядра яйцеклетки, которое возобновилось благодаря оплодотворению. Постепенно ядро яйцеклетки превращается в женский пронуклеус. Пронуклеусы перемещаются к центру яйцеклетки, происходит репликация ДНК, и после их слияния набор хромосом и ДНК зиготы становится 2n4c. Объединение пронуклеусов и представляет собой собственно оплодотворение. Таким образом, оплодотворение заканчивается образованием зиготы с диплоидным ядром.
Оплодотворение — необратимый процесс, то есть однажды оплодотворенное яйцо не может быть оплодотворено вновь. В зависимости от количества сперматозоидов, проникающих в яйцеклетку при оплодотворении, различают:
моноспермию — оплодотворение, при котором в яйцо проникает только один сперматозоид (наиболее обычное оплодотворение);
полиспермию — оплодотворение, при котором в яйцеклетку проникает несколько сперматозоидов (некоторые птицы, рептилии). Но даже в этом случае с ядром яйцеклетки сливается ядро только одного из сперматозоидов, а остальные ядра разрушаются.
В зависимости от количества особей, принимающих участие в половом размножении, различают:
перекрестное оплодотворение — оплодотворение, в котором принимают участие гаметы, образованные разными организмами;
самооплодотворение — оплодотворение, при котором сливаются гаметы, образованные одним и тем же организмом (некоторые растения, паразитические черви).
Партеногенез В некоторых группах организмов половое размножение происходит при участии гамет, но без оплодотворения. Партеногенез (девственное размножение) — это развитие организма из неоплодотворенного яйца. Партеногенез известен у всех типов беспозвоночных животных и у всех позвоночных, кроме млекопитающих, у которых партеногенетические зародыши погибают на ранних стадиях эмбриогенеза. Он может быть:
Искусственным, вызывается человеком путем активизации яйцеклетки в результате воздействия на нее различными веществами, механическим раздражением, повышением температуры и т.д.
Естественным, если яйцо начинает дробиться и развивается в эмбрион без участия сперматозоида, только под влиянием внутренних или внешних причин. Причем различают:
соматический, или диплоидный, если мейоз не происходит, и развитие начинается с диплоидных ооцитов, или если мейоз произошел, но сливаются два гаплоидных ядра, восстанавливая диплоидный набор хромосом (тли, дафнии, одуванчики);
генеративный, или гаплоидный, если зародыш начинает развиваться из гаплоидной яйцеклетки (трутни пчел). Как правило, возникающие при этом организмы гаплоидны.
Если развитие яйцеклетки происходит без участия ядра сперматозоида (некоторые рыбы, круглые черви), то такая разновидность партеногенеза называется гиногенезом. Однако именно сперматозоид стимулирует начало дробления яйцеклетки, хотя и не оплодотворяет ее.
Если развитие яйца происходит только за счет генетического материала сперматозоидов и цитоплазмы яйцеклетки, то в этом случае говорят об андрогенезе. Этот тип развития может осуществляться в том случае, если ядро яйцеклетки погибает еще до оплодотворения, а в яйцеклетку попадает не один, а несколько сперматозоидов (тутовый шелкопряд).
38.5. Эмбриональное развитие
Дробление После оплодотворения зигота начинает делиться. Дроблением называют ряд последовательных митотических делений зиготы, в результате которых огромный объем цитоплазмы яйца разделяется на многочисленные, содержащие ядра клетки меньшего размера (рис. 312). В результате дробления образуются клетки, которые называют бластомерами. Важной отличительной особенностью дробления от обычного деления является то, что вновь образовавшиеся бластомеры не увеличиваются в размерах. Это осуществляется путем выпадения интерфазного периода роста между делениями. При этом синтетический период интерфазы начинается в телофазе предшествующего митотического цикла. Таким образом, количество бластомеров постепенно увеличивается, а их общий объем практически не изменяется. Цитоплазма клеток при дроблении делится путем возникновения впячиваний оболочки клетки (борозды дробления).
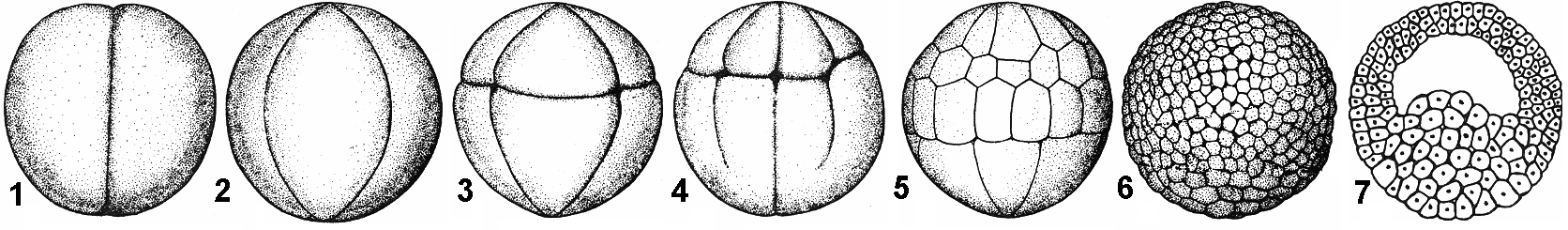
Рис. 312. Дробление яйцеклетки амфибий (лягушка):
1 — двуклеточная стадия; 2 — четырехклеточная стадия; 3 — восьмиклеточная стадия; 4 — переход от восьми- к шестнадцатиклеточной стадии (клетки анимального полюса уже поделились, а клетки вегетативного только начинают дробиться; 5 — более поздняя стадия дробления; 6 — бластула; 7 — бластула в разрезе.
Однако дробление не может происходить бесконечно. Так как каждое деление дробления сопровождается уменьшением размера клетки, постепенно происходит повышение величины ядерно-цитоплазматического отношения, сниженного в период роста ооцита. Наступает момент, когда это отношение достигает значения, типичного для соматических клеток данного вида.
Биологическое значение процесса дробления сводится к следующему:
благодаря повторяющимся циклам репродукции, происходит размножение генотипа зиготы;
происходит накопление клеточной массы для дальнейших преобразований, т.е. зародыш из одноклеточного превращается в многоклеточный.
Деление бластомеров бывает синхронным и несинхронным. У большинства видов оно несинхронно с самого начала развития, у других становится таковым уже после первых делений.
Характер дробления определяется, прежде всего, строением яйцеклетки, главным образом, количеством желтка и особенностями его распределения в цитоплазме. В этой связи по способу дробления выделяют два основных типа яиц (рис. 313):
полностью дробящиеся;
дробящиеся частично.
Полное дробление Полным дробление называется тогда, когда цитоплазма яйцеклетки полностью разделяется на бластомеры. Оно может быть:
равномерным, при котором все образовавшиеся бластомеры имеют одинаковые размеры и форму; оно характерно для алецитальных и изолецитальных яйцеклеток;
неравномерным, при котором образуются неравные по размерам бластомеры; свойственно телолецитальным яйцеклеткам с умеренным содержанием желтка; мелкие бластомеры возникают у анимального полюса, крупные — в области вегетативного полюса зародыша.

Рис. 313. Различные типы дробления:
А — полное; Б — частичное; В — дискоидальное.
Частичное дробление Частичное дробление — тип дробления, при котором цитоплазма яйцеклетки не полностью разделяется на бластомеры. Одним из видов частичного дробления является дискоидальное, при котором дроблению подвергается только лишенный желтка участок цитоплазмы у анимального полюса, где находится ядро. Участок цитоплазмы, подвергшийся дроблению, называется зародышевым диском. Этот тип дробления характерен для резко телолецитальных яиц с большим количеством желтка (рептилии, птицы, рыбы);
Образование бластулы Дробление у представителей разных групп животных имеет свои особенности, однако завершается оно образованием близкой по строению структуры — бластулы.
Бластула — это однослойный зародыш. Она состоит из слоя клеток — бластодермы, ограничивающей полость — бластоцель, или первичную полостью тела. Бластула формируется начиная с ранних этапов дробления, благодаря расхождению бластомеров. Возникающая при этом полость заполняется жидкостью.
Типы бластул Строение бластулы во многом зависит от типа дробления (рис. 314).
Целобластула (типичная бластула). Образуется при равномерном дроблении. Имеет вид однослойного пузырька с большим бластоцелем (у ланцетника).
Амфибластула. При дроблении телолецитальных яиц бластодерма построена из бластомеров разного размера: микромеров на анимальном и макромеров на вегетативном полюсах. Бластоцель при этом смещается в сторону анимального полюса (у земноводных).
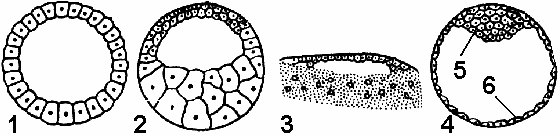
Рис. 314. Типы бластул:
1 — целобластула; 2 — амфибластула; 3 — дискобластула; 4 — бластоциста; 5 — эмбриобласт; 6 — трофобласт.
Дискобластула. Образуется при дискоидальном дроблении. Полость бластулы имеет вид узкой щели, находящейся под зародышевым диском (у птиц).
Бластоциста. Представляет собой однослойный пузырек, заполненный жидкостью, в котором различают эмбриобласт (из него развивается зародыш) и трофобласт, обеспечивающий питание зародыша (у млекопитающих).
Гаструляция После того как сформировалась бластула, начинается новый этап эмбриогенеза — гаструляция (образование зародышевых листков). Для гаструляции характерны интенсивные перемещения отдельных клеток и клеточных масс. Деление клеток при гаструляции отсутствует или выражено очень слабо. В результате гаструляции образуется двухслойный, а затем трехслойный зародыш (у большинства животных) — гаструла (рис. 315). Первоначально образуются наружный (эктодерма) и внутренний (энтодерма). Позже между экто- и энтодермойзакладывается третий зародышевый листок — мезодерма.
Зародышевые листки — это отдельные пласты клеток, занимающие определенное положение в зародыше и дающие начало соответствующим органам и системам органов. Зародышевые листки возникают не только в результате перемещения клеточных масс, но и в результате дифференциации сходных между собой сравнительно однородных клеток бластулы. В процессе гаструляции зародышевые листки занимают положение, соответствующее плану строения взрослого организма. Дифференциация — это процесс появления и нарастания морфологических и функциональных различий между отдельными клетками и частями зародыша.
Способы гаструляции
В
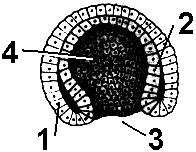
Рис. 315. Гаструла.
1 — эктодерма; 2 — энтодерма; 3 — бластопор;
4 — гастроцель.
зависимости от типа бластулы и от особенностей перемещения клеток, различают следующие основные способы образования двухслойного зародыша, или способы гаструляции (рис. 316):
Инвагинация. При данном способе один из участков бластодермы начинает впячиваться внутрь бластоцеля (у ланцетника). При этом бластоцель практически полностью вытесняется. Образуется двухслойный мешок, наружная стенка которого является первичной эктодермой, а внутренняя — первичной энтодермой, выстилающей полость первичной
кишки, или гастроцель. Отверстие, при помощи которого полость сообщается с окружающей средой, называется бластопором, или первичным ртом. У представителей разных групп животных судьба бластопора различна. У первичноротых животных он превращается в ротовое отверстие. У вторичноротых бластопор зарастает, и на его месте нередко возникает анальное отверстие, а ротовое отверстие прорывается на противоположном полюсе (переднем конце тела).
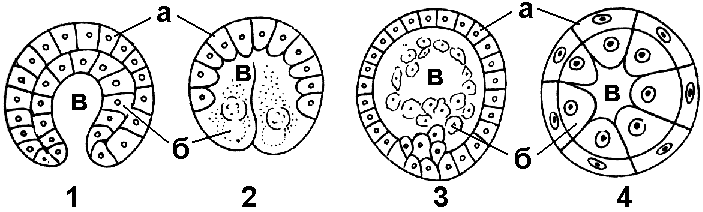
Рис. 316. Типы гаструл:
1 — инвагинационная; 2 — эпиболическая; 3 — иммиграционная; 4 — деламинационная; а — эктодерма; б — энтодерма; в — гастроцель
Иммиграция — выселение части клеток бластодермы в полость бластоцеля (у высших позвоночных). Из них образуется энтодерма.
Деламинация встречается у животных, имеющих бластулу без бластоцеля (у птиц). При таком способе гаструляции клеточные перемещения минимальны или совсем отсутствуют, так как происходит расслоение — наружные клетки бластулы преобразуются в эктодерму, а внутренние формируют энтодерму.
Эпиболия происходит, когда более мелкие бластомеры анимального полюса дробятся быстрее и обрастают более крупные бластомеры вегетативного полюса, образуя эктодерму (у земноводных). Клетки вегетативного полюса дают начало внутреннему зародышевому листку — энтодерме.
Описанные способы гаструляции редко встречаются в чистом виде и обычно наблюдаются их сочетания (инвагинация с эпиболией у амфибий или деляминация с иммиграцией у иглокожих).
Образование мезодермы
Чаще всего клеточный материал мезодермы входит в состав энтодермы. Он впячивается в бластоцель в виде карманообразных выростов, которые затем отшнуровываются.
При образовании мезодермы происходит образование вторичной полости тела, или целома.
Первичный органогенез Процесс формирования органов в эмбриональном развитии называют органогенезом. В построении любого органа участвуют несколько тканей. Поэтому стадия органогенеза является и стадией гистогенеза.
В органогенезе можно выделить две фазы:
нейруляция — образование комплекса осевых органов (нервная трубка, хорда, кишечная трубка и мезодерма сомитов), в который вовлекается почти весь зародыш;
построение остальных органов, приобретение различными участками тела типичной для них формы и черт внутренней организации, установление определенных пропорций (пространственно ограниченные процессы).
По теории зародышевых листков Карла Бэра, возникновение органов обусловлено преобразованием того или иного зародышевого листка — экто-, мезо- или энтодермы. Некоторые органы могут иметь смешанное происхождение, то есть они образованы при участии сразу несколько зародышевых листков. Например, мускулатура пищеварительного тракта является производным мезодермы, а его внутренняя выстилка — производное энтодермы. Однако, несколько упрощая, происхождение основных органов и их систем все-таки можно связать с определенными зародышевыми листками.
Нейруляция Зародыш на стадии нейруляции называется нейрулой (рис. 317). Материал, используемый на построение нервной системы у позвоночных животных — нейроэктодерма, входит в состав спинной (дорсальной) части эктодермы. Он располагается над зач
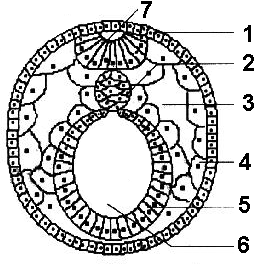
Рис. 317. Нейрула:
1 — эктодерма; 2 — хорда; 3 — вторичная полость тела; 4 — мезодерма; 5 — энтодерма; 6 — кишечная полость; 7 — нервная трубка.
атком хорды. Взаимодействие этихзачатков
является одним из наиболее важных во всем развитии. Сначала в области нейроэктодермы происходит уплощение клеточного пласта, что приводит к образованию нервной пластинки. Затем края нервной пластинки утолщаются и приподнимаются, образуя нервные валики. В центре пластинки за счет перемещения клеток по средней линии возникает нервный желобок, разделяющий зародыш на будущие правую и левую половины. Нервная пластинка начинает складываться по средней линии. Края ее соприкасаются, а затем смыкаются. В результате этих процессов возникает нервная трубка с полостью — невроцелем.
Смыкание валиков происходит сначала в средней, а затем в задней части нервного желобка. В последнюю очередь это происходит в головной части, которая по ширине превосходит другие. Передний, расширенный отдел в дальнейшем образует головной мозг, остальная часть нервной трубки — спинной. В результате нервная пластинка превращается в нервную трубку, лежащую под эктодермой.
В ходе нейруляции часть клеток нервной пластинки не входят в состав нервной трубки. Они образуют ганглиозную пластинку, или нервный гребень, — скопление клеток вдоль нервной трубки. Позднее эти клетки мигрируют по всему зародышу, образуя клетки нервных узлов, мозгового вещества надпочечников, пигментные клетки и т.п.
Образование систем органов Из материала эктодермы, помимо нервной трубки, развиваются эпидермис и его производные (перо, волосы, ногти, когти, кожные железы и т.д.), компоненты органов зрения, слуха, обоняния, эпителий ротовой полости, эмаль зубов.
Мезодермальные и энтодермальные органы формируются не после образования нервной трубки, а одновременно с ней. Практически одновременно с нейруляцией происходят процессы закладки мезодермы и хорды. Вначале вдоль боковых стенок первичной кишки путем выпячивания энтодермы образуются карманы, или складки. Участок энтодермы, расположенный между этими складками, утолщается, прогибается, сворачивается и отшнуровывается от основной массы энтодермы. Так появляется хорда. Возникшие карманообразные выпячивания энтодермы отшнуровываются от первичной кишки и превращаются в ряд сегментарно-расположенных замкнутых мешков, называемых также целомическими мешками. Их стенки образованы мезодермой, а полость внутри представляет собой вторичную полость тела (или целом).
Из мезодермы развиваются все виды соединительной ткани, дерма, скелет, поперечно-полосатая и гладкая мускулатура, кровеносная и лимфатическая системы, половая система.
Из материала энтодермы развивается эпителий кишечника и желудка, клетки печени, секретирующие клетки поджелудочной, кишечных и желудочных желез. Передний отдел эмбриональной кишки образует эпителий легких и воздухоносных путей, секретирующие отделы передней и средней доли гипофиза, щитовидной и паращитовидной желез.
Эмбриональная индукция Наблюдения за оплодотворенной яйцеклеткой лягушки позволили проследить путь развития клеток, входящих в состав того или иного участка зародыша. Оказалось, что определенные клетки, занимающие соответствующее место в бластуле, дают начало строго определенным зачаткам органов. Удалось выяснить, какие группы клеток дают начало нервной трубке, хорде, мезодерме, кожному эпителию и т.д. Действительно, в развивающемся организме различные группы клеток дают начало определенным органам и тканям, а культивирование клеток вне зародыша (в пробирке) не приводит к формированию типичных тканевых структур, которые должны были бы образоваться из клеток. Чем же вызывается преобразование тех или иных клеток зародыша в конкретные ткани и органы?
В 1924 г. были опубликованы результаты опытов Г.Шпемана и Г.Мангольда, посвященные выяснению этого вопроса (рис. 318). На стадии ранней гаструлы зачаток эктодермы, который в нормальных условиях должен был развиться в структуры нервной системы, из зародыша гребенчатого (непигментриованного) тритона пересаживался под эктодерму брюшной стороны, дающую начало эпидермису кожи, зародыша обыкновенного (пигментированного) тритона. В итоге на брюшной стороне зародыша-реципиента возникала сначала нервная трубка и другие компоненты комплекса осевых органов, а затем формировался дополнительный зародыш. Причем, наблюдения показали, что ткани дополнительного зародыша формируются почти исключительно из клеточного материала реципиента.
Эти данные доказывают, что в ходе эмбриогенеза некоторые части зародыша влияют на пути развития соседних участков. Такое влияние одного зачатка на другой получило название эмбриональной индукции. Насколько важную роль играет эмбриональная индукция в развитии, показывает следующий опыт. Если на стадии ранней гаструлы полностью удалить
з
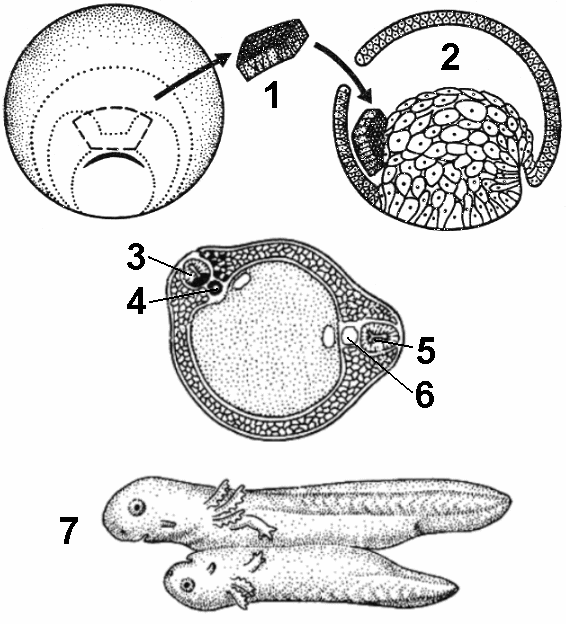
Рис 318. Эмбриональная индукция:
1 — зачаток хордомезодермы; 2 — полость бластулы; 3 — индуцирорванная нервная трубка; 4 — индуцированная хорда; 5 — первичная нервная трубка; 6 — первичная хорда; 7 — формирование вторичного зародыша, соединенного с зародышем-хозяином.
ачаток хорды, то нервная трубка совсем не развивается. Эктодерма на спинной стороне зародыша, из которой в норме формируется нервная трубка, образует кожный эпителий.
При дальнейшем изучении развития зародышей оказалось, что зачаток хордомезодермы представляет собой не только индуктор нервной трубки, но и сам для дифференцировки нуждается в индуцирующем влиянии со стороны зачатка нервной системы. Во время эмбрионального развития имеет место не односторонняя индукция, а взаимодействие частей развивающегося зародыша. Таким образом, эмбриональную индукцию можно определить как явление, при котором в процессе эмбриогенеза один зачаток влияет на другой, определяя путь его развития, и, кроме того, сам подвергается индуцирующему воздействию со стороны первого зачатка.
38.6. Постэмбриональное развитие
Постэмбриональный период развития начинается в момент рождения или выхода организма из яйцевых оболочек и продолжается вплоть до его смерти. Постэмбриональное развитие включает в себя:
рост организма;
установление окончательных пропорций тела;
переход систем органов на режим взрослого организма (в частности, половое созревание).
Типы постэмбрионального развития Различают два основных типа постэмбрионального развития:
Прямое, при котором из тела матери или яйцевых оболочек выходит особь, отличающаяся от взрослого организма только меньшим размером (птицы, млекопитающие). Различают:
неличиночный (яйцекладный) тип, при котором зародыш развивается внутри яйца (рыбы, птицы);
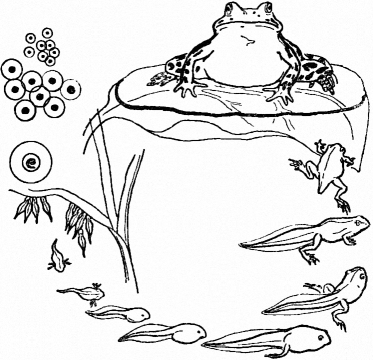
Рис. 319. Развитие лягушки
внутриутробный тип, при котором зародыш развивается внутри организма матери и связан с ним через плаценту (плацентарные млекопитающие).
С превращением (метаморфозом), при котором из яйца выходит личинка, устроенная проще взрослого животного (иногда сильно отличающаяся от него); как правило, она имеет специальные личиночные органы, отсутствующие у взрослого животного, и не способна к размножению; часто личинка ведет иной образ жизни, чем взрослое животное (насекомые, некоторые паукообразные, амфибии).
Примером животных, имеющих постэмбриональное развитие с метаморфозом, служат бесхвостые земноводные (рис. 319). Из яйцевых оболочек земноводных выходит личинка — головастик, больше напоминающий рыбу, чем земноводное. Он имеет обтекаемую форму тела, хвостовой плавник, жаберные щели и жабры, органы боковой линии, двухкамерное сердце, один круг кровообращения. Со временем, под влиянием гормона щитовидной железы, головастик претерпевает метаморфоз. У него рассасывается хвост, появляются конечности,
исчезает боковая линия, развиваются легкие и второй круг кровообращения, то есть постепенно он приобретает признаки, характерные для земноводных.
|
|
|
 Скачать 34.42 Mb.
Скачать 34.42 Mb.