Пособие по биологии. Пособие по биологии для поступающих в вузы Авторы Пименов А. В., Гончаров О. В
 Скачать 34.42 Mb. Скачать 34.42 Mb.
|

Глава 36. Строение клеткиКлетка — элементарная живая система, единица строения, жизнедеятельности, размножения и развития живых организмов. Это самая простая (элементарная) живая система, способная к самообновлению, саморегуляции и самовоспроизведению. В зависимости от количества клеток, образующих организм, различают:
Клетки живых организмов очень разнообразны: они отличаются друг от друга формой, размерами, особенностями организации и функциями. По форме различают шаровидные, цилиндрические, призматические, кубические, удлиненные, дисковидные, звездчатые и другие клетки. Наиболее часто встречаются клетки шаровидной или овальной формы. 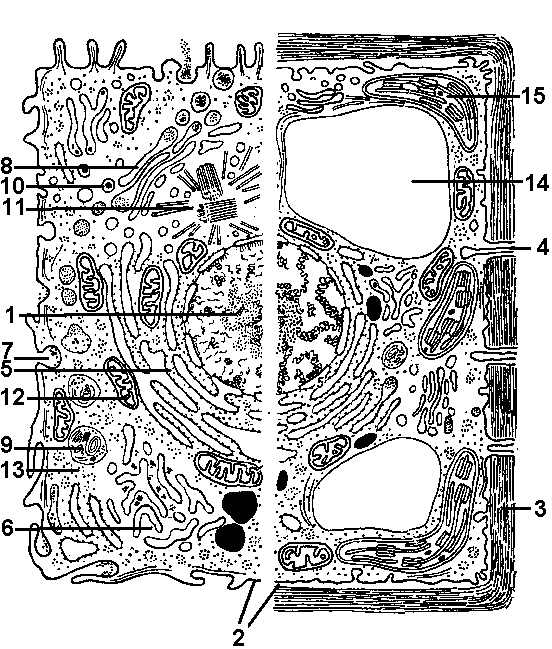 Рис. 279. Схема строения эукариотической клетки (слева животной, справа — растительной): 1 — ядро с хроматином и ядрышком; 2 — цитоплазматическая мембрана; 3 — клеточная стенка; 4 — плазмодесмы; 5 — гранулярная эндоплазматическая сеть; 6 агранулярная эндоплазматическая сеть; 7 — пиноцитозная вакуоль; 8 — комплекс Гольджи; 9 — лизосома; 10 — жировые включения; 11 — центриоль и микротрубочки центросферы; 12 — митохондрии; 13 — полисомы; 14 — вакуоль; 15 — хлоропласты. Разнообразны и размеры клеток. Большинство клеток имеют размеры от 10 до 100 мкм, реже — 1-10 мм (клетки мякоти арбуза) и очень редко от 5 до 10 см (яйца птиц — гусей, пингвинов, страусов). В зависимости от наличия в клетке оформленного ядра различают два уровня клеточной организации:
В этой главе будут рассмотрены особенности организации только эукариотической клетки. Как правило, эукариотическая клетка состоит из трех неразрывно связанных жизненно важных частей (рис. 279):
36.1. Клеточные мембраныВ основе структурной организации клетки лежит мембранный принцип строения, то есть клетка в основном построена из мембран. Все биологические мембраны имеют общие структурные особенности и свойства. В настоящее время общепринята жидкостно-мозаичная модель строения мембраны. Химический состави строение мембраныОснову мембраны составляет липидный бислой, образованный в основном фосфолипидами. Липиды составляют в среднем ≈40% химического состава мембраны. В бислое хвосты молекул в мембране обращены друг к другу, а полярные головки — наружу, поэтому поверхность мембраны гидрофильна. Липиды определяют основные свойства мембран.Помимо липидов в состав мембраны входят белки (в среднем ≈60%). Они определяют большинство специфических функций мембраны. Молекулы белков не образуют сплошного слоя (рис. 280). В зависимости от локализации в мембране различают:
Мембранные белки могут выполнять различные функции:
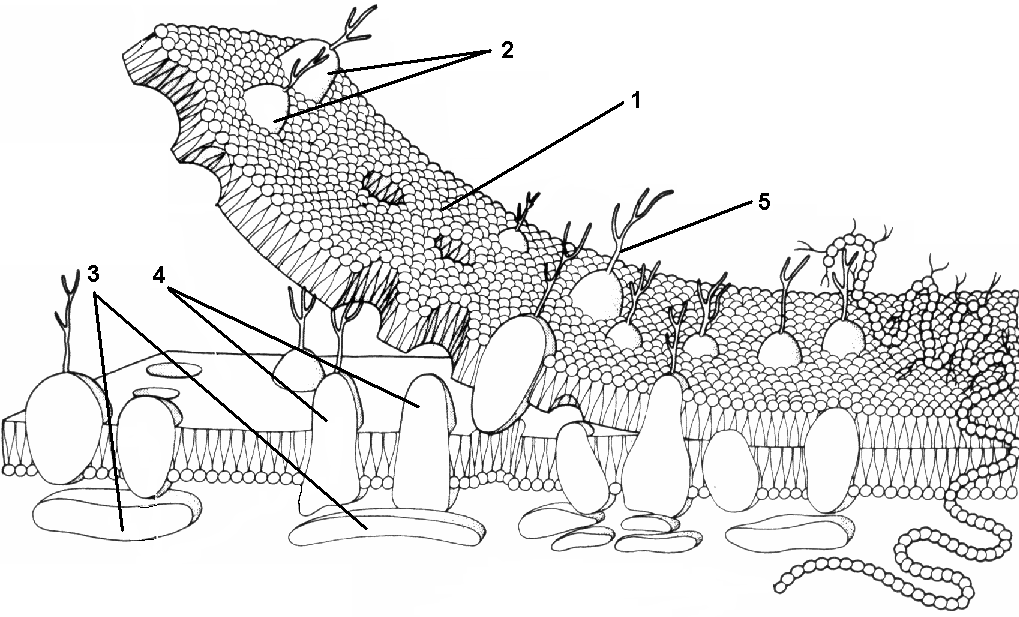
В состав мембраны может входить от 2 до 10% углеводов. Углеводный компонент мембран обычно представлен олигосахаридными или полисахаридными цепями, связанными с молекулами белков (гликопротеины) или липидов (гликолипиды). В основном углеводы располагаются на наружной поверхности мембраны. Функции углеводов клеточной мембраны до конца не выяснены, однако можно сказать, что они обеспечивают рецепторные функции мембраны. В животных клетках гликопротеины образуют надмембранный комплекс — гликокаликс, имеющий толщину в несколько десятков нанометров. В нем происходит внеклеточное пищеварение, располагаются многие рецепторы клетки, с его помощью, по-видимому, происходит адгезия клеток. Молекулы белков и липидов подвижны, способны перемещаться, главным образом, в плоскости мембраны. Мембраны асимметричны, то есть липидный и белковый состав наружной и внутренней поверхности мембраны различен. Толщина плазматической мембраны в среднем 7,5 нм. Функции мембранКлеточные мембраны играют важную роль по ряду причин:
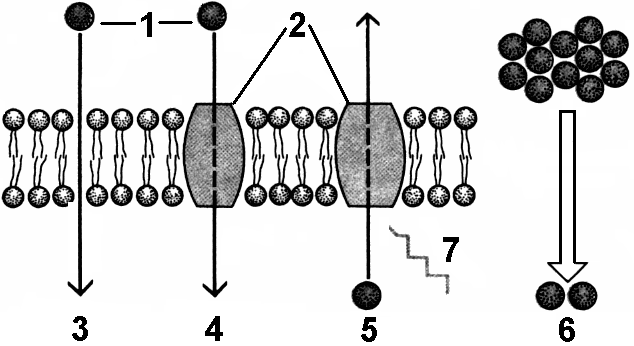
Транспорт веществ через мембрануОдна из основных функций мембраны — транспортная, обеспечивающая обмен веществ между клеткой и внешней средой. Мембраны обладают свойством избирательной проницаемости, то есть хорошо проницаемы для одних вещества или молекул и плохо проницаемы (или совсем непроницаемы) для других. Проницаемость мембран для разных веществ зависит и от свойств их молекул (полярность, размер и т.д.), и от характеристики мембран (внутренняя часть липидного слоя гидрофобна).Существуют различные механизмы транспорта веществ через мембрану (рис. 281). В зависимости от необходимости использования энергии для осуществления транспорта веществ, различают:
Пассивный транспортВ основе пассивного транспорта лежит разность концентраций и зарядов. При пассивном транспорте вещества всегда перемещаются из области с более высокой концентрацией в область с более низкой, то есть по градиенту концентрации. Если молекула заряжена, то на ее транспорт влияет и электрический градиент. Поэтому часто говорят об электрохимическом градиенте, объединяя оба градиента вместе. Скорость транспорта зависит от величины градиента.Различают три основных механизма пассивного транспорта:
Активный транспортНеобходимость активного транспорта возникает тогда, когда требуется обеспечить перенос через мембрану молекул против электрохимического градиента. Этот транспорт осуществляется белками-переносчиками, деятельность которых требует затрат энергии. Источником энергии служат молекулы АТФ.Одной из наиболее изученных систем активного транспорта является натрий-калиевый насос. Концентрация K внутри клетки значительно выше, чем за ее пределами, а Na — наоборот. Поэтому К через водяные поры мембраны пассивно диффундирует из клетки, а Na — в клетку. Вместе с тем, для нормального функционирования клетке важно поддерживать определенное соотношение ионов К и Na в цитоплазме и во внешней среде. Это оказывается возможным потому, что мембрана, благодаря наличию (Na + K)-насоса, активно перекачивает Na из клетки, а K в клетку. На работу (Na + K)-насоса тратится почти треть всей энергии, необходимой для жизнедеятельности клетки. 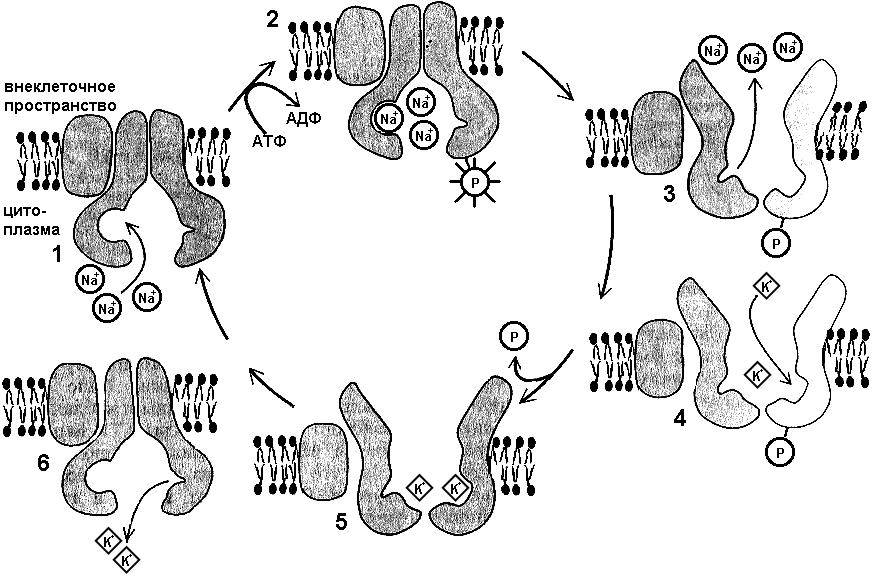 Рис. 282. Натрий-калиевый насос: 1 — присоединение ионов натрия; 2 — фосфорилирование транспортного белка; 3 — высвобождение ионов натрия; 4 — присоединение ионов калия; 5 — дефосфорилирование транспортного белка; 6 — высвобождение ионов калия. Насос представляет собой особый трансмембранный белок мембраны, способный к конформационным изменениям, благодаря чему он может присоединять к себе как ионы К, так и ионы Na. Цикл работы (Na + K)-насоса складывается из нескольких фаз (рис. 282):
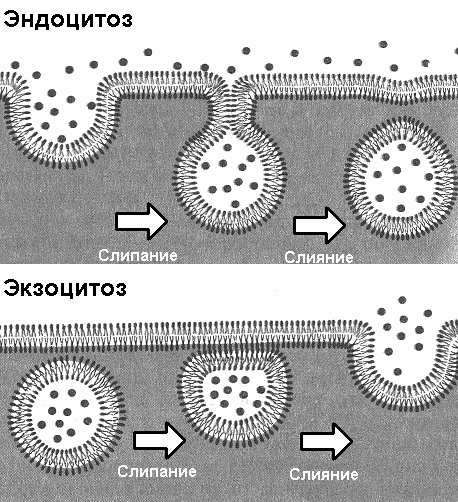
За один цикл работы насос выкачивает из клетки 3 иона Na и закачивает 2 иона К. Такая разница в количестве переносимых ионов связана с тем, что проницаемость мембраны для ионов К выше, чем для ионов Na. Соответственно K быстрее пассивно диффундирует из клетки, чем Na в клетку. Эндоцитоз и экзоцитозКлетка имеет механизмы, благодаря которым может осуществлять транспорт через мембрану крупных частиц и макромолекул (рис. 283). Процесс поглощения макромолекул клеткой называется эндоцитозом. При эндоцитозе плазматическая мембрана образует впячивание, края ее сливаются, и происходит отшнуровывание в цитоплазму везикул — мешочкоподобных структур, отграниченных от цитоплазмы одиночной мембраной, являющейся частью наружной цитоплазматической мембраны. Различают два типа эндоцитоза:
Экзоцитоз — процесс выведения различных веществ из клетки. При экзоцитозе мембрана везикулы (или вакуоли), при соприкосновении с наружной цитоплазматической мембраной, сливается с ней. Содержимое везикулы выводится за пределы летки, а ее мембрана включается в состав наружной цитоплазматической мембраны. 36.2. Цитоплазма. ОрганоидыЦитоплазма — обязательная часть клетки, заключенная между плазматической мембраной и ядром и представляющая собой сложный гетерогенный структурный комплекс клетки, состоящий из:
Химический состав цитоплазмы разнообразен. Ее основу составляет вода (60-90% всей массы цитоплазмы). Цитоплазма богата белками (10-20%, иногда до 70% и более сухой массы), которые составляют ее основу. Помимо белков, в состав цитоплазмы могут входить жиры и жироподобные вещества (2-3%), различные органические и неорганические соединения (по 1,5%). Цитоплазма имеет щелочную реакцию Одна из характерных особенностей цитоплазмы — постоянное движение (циклоз). Оно обнаруживается, прежде всего, по перемещению органелл клетки, например хлоропластов. Если движение цитоплазмы прекращается, клетка погибает, так как, только находясь в постоянном движении, она может выполнять свои функции. ГиалоплазмаОсновное вещество цитоплазмы — гиалоплазма11 (основная плазма, матрикс12 цитоплазмы) представляет собой бесцветный, слизистый, густой и прозрачный коллоидный раствор. Именно в ней протекают все процессы обмена веществ, она обеспечивает взаимосвязь ядра и всех органоидов. Жидкая часть гиалоплазмы представляет собой истинный раствор ионов и малых молекул, в которой во взвешенном состоянии находятся крупные молекулы белков и РНК. В зависимости от преобладания в гиалоплазме жидкой части или крупных молекул, различают две формы гиалоплазмы:
Между ними возможны взаимопереходы: гель легко превращается в золь и наоборот. ОрганоидыОрганоиды (органеллы) — постоянные клеточные структуры, обеспечивающие выполнение клеткой специфических функций. Каждый органоид имеет определенное строение и выполняет определенные функции. В зависимости от особенностей строения, различают:мембранные органоиды — имеющие мембранное строение, причем они могут быть: одномембранными (эндоплазматический ретикулум, аппарат Гольджи, лизосомы, вакуоли растительных клеток); двумембранными (митохондрии, пластиды); немембранные органоиды — не имеющие мембранного строения (хромосомы, рибосомы, клеточный центр и центриоли, реснички и жгутики с базальными тельцами, микротрубочки, микрофиламенты). Есть органоиды, свойственные всем клеткам, – митохондрии, клеточный центр, аппарат Гольджи, рибосомы, эндоплазматический ретикулум, лизосомы. Их называют органоидами общего значения. Имеются органоиды, характерные только для определенных типов клеток, специализированных к выполнению определенной функции (например, миофибриллы, обеспечивающие сокращение мышечного волокна). Их называют специальными органоидами. Эндоплазматическийретикулум (ЭПР)Одномембранный органоид, представляющий собой систему мембран, формирующих цистерны и каналы, соединенных друг с другом и ограничивающих единое внутреннее пространство — полость ЭПР. Мембраны с одной стороны связаны с наружной цитоплазматической мембраной, с другой — с наружной оболочкой ядерной мембраны. Наибольшего развития ЭПР достигает в клетках с интенсивным обменом веществ. В среднем он составляет от 30 до 50 % всего объема клетки.Различают три вида ЭПР:
Функции ЭПР:
Аппарат ГольджиП 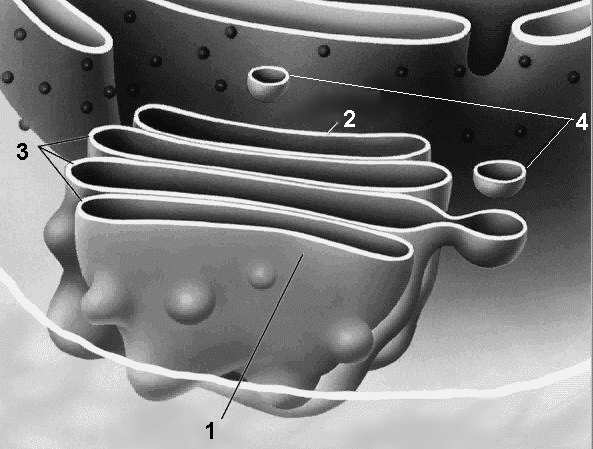 Рис.284. Аппарат Гольджи: 1 — секретирующий полюс; 2 — формирующий полюс; 3 — цистерны аппарата Гольджи; 4 — пузырьки Гольджи. ластинчатый комплекс, комплекс Гольджи (рис. 284). Одномембранный органоид, обычно расположенный около клеточного ядра (в животных клетках часто вблизи клеточного центра). Представляет собой стопку уплощенных цистерн с расширенными краями, с которой связана система мелких одномембранных пузырьков (пузырьки Гольджи). Каждая стопка обычно состоит из 4-6 цистерн. Число стопок Гольджи в клетке колеблется от одной до нескольких сотен. Пузырьки Гольджи в основном сконцентрированы на стороне, примыкающей к ЭПР, и по периферии стопок. Полагают, что они переносят в аппарат Гольджи белки и липиды, молекулы которых, передвигаясь из цистерны в цистерну, подвергаются химической модификации. Важнейшая функция комплекса Гольджи — выведение из клетки различных секретов (ферментов, гормонов), поэтому он хорошо развит в секреторных клетках. У аппарата Гольджи выделяют две разные стороны:
Наружная часть аппарата Гольджи постоянно расходуется в результате отшнуровывания пузырьков, а внутренняя — постепенно формируется за счет деятельности ЭПР. Функции аппарата Гольджи:
ЛизосомыСамые мелкие одномембранные органоиды клетки, представляющие собой пузырьки диаметром 0,2-0,8 мкм, содержащие около 40 гидролитических ферментов (протеазы, липазы, нуклеазы, фосфотазы), активных в слабокислой среде (рис. 285). Образование лизосом происходит в аппарате Гольджи, куда из ЭПР поступают синтезированные в нем ферменты. Расщепление веществ с помощью ферментов называют лизисом, отсюда и название органоида.Р 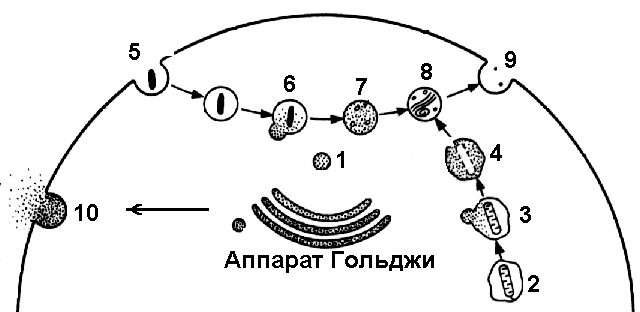 Рис. 285. Лизосомы: 1 — первичная лизосома; 2 — митохондрия, окруженная мембраной; 3 — автофагичнеская вакуоль; 4 — переваривание митохондрии; 5 — эндоцитоз; 6 — образование вторичной вакуоли; 7 — переваривание; 8 — остаточное тельце; 9 — выделение содержимого остаточного тельца путем экзоцитоза; 10 — выделение лизосомных ферментов путем экзоцитоза. азличают:
Продукты переваривания усваиваются цитоплазмой клетки, но часть материала так и остается непереваренной. Вторичная лизосома, содержащая этот непереваренный материал, называется остаточным тельцем. Путем экзоцитоза непереваренные частицы удаляются из клетки. Вторичная лизосома, переваривающая отдельные составные части клетки, называется автофагической вакуолью. Подлежащие уничтожение части клетки окружаются одинарной мембраной, обычно отделяющейся от гладкого ЭПР, а затем образовавшийся мембранный мешочек сливается с первичной лизосомой, в результате чего и происходит образование автофагической вакуоли. Иногда с участием лизосом происходит саморазрушение клетки. Этот процесс называют автолизом. Обычно это происходит при некоторых процессах дифференцировки (например, замена хрящевой ткани костной, исчезновение хвоста у головастика лягушек). Функции лизосом:
МитохондрииДвумембранные органоиды эукариотической клетки, обеспечивающие организм энергией (рис. 286). Они имеют палочковидную, нитевидную, шаровидную, спиральную, чашевидную и т.д. форму. Длина митохондрий 1,5-10 мкм, диаметр — 0,25-1,00 мкм.К 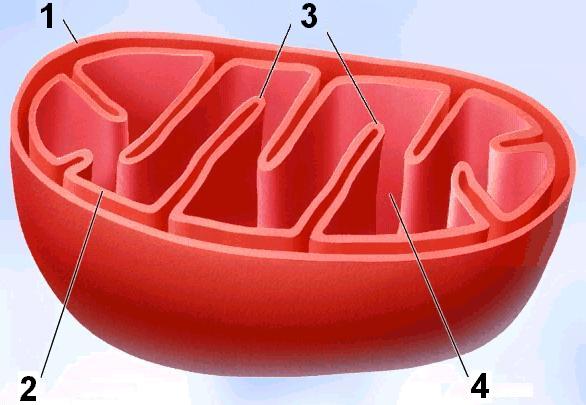 Рис. 286. Митохондрия: 1 — наружная мембрана; 2 — внутренняя мембрана; 3 — кристы; 4 — матрикс. оличество митохондрий в клетке колеблется в широких пределах, от 1 до 100 тыс., и зависит от ее метаболической активности. Число митохондрий может увеличиваться путем деления, так как эти органоиды имеют собственную ДНК. Наружная мембрана митохондрий гладкая, внутренняя мембрана образует многочисленные впячивания (гребни) или трубчатые выросты — кристы13, обладающие строго специфичной проницаемостью и системами активного транспорта. Число крист может колебаться от нескольких де- сятков до нескольких сотен и даже тысяч, в зависимости от функций клетки. Они увеличивают поверхность внутренней мембраны, на которой размещаются мультиферментные системы, участвувующие в синтезе молекул АТФ. Внутренняя мембрана содержит белки двух главных типов:
Наружная мембрана отделена от внутренней межмембранным пространством. Внутреннее пространство митохондрий заполнено гомогенным веществом — матриксом. В матриксе содержатся кольцевые молекулы митохондриальной ДНК, специфические иРНК, тРНК и рибосомы (прокариотического типа), осуществляющие автономный биосинтез части белков, входящих в состав внутренней мембраны. Но большая часть генов митохондрии перешла в ядро, и синтез многих митохондриальных белков происходит в цитоплазме. Кроме того, содержатся ферменты, образующие молекулы АТФ. Митохондрии способны размножаться путем деления или отшнуровывания мелких фрагментов. Функции митохондрий:
РибосомыН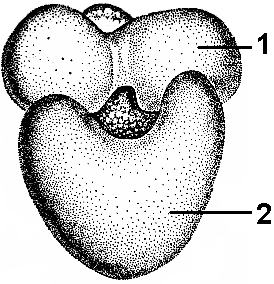 Рис. 287. Рибосома: 1 — малая субъединица; 2 — большая субъединица. емембранные органоиды, встречающиеся в клетках всех организмов. Это мелкие органеллы, представленные глобулярными частицами диаметром порядка 20 нм (рис. 287). Рибосомы состоят из двух субъединиц неравного размера — большой и малой, на которые они могут диссоциировать. В состав рибосом входят белки и рибосомальные РНК (рРНК). Молекулы рРНК составляют 50-63% массы рибосомы и образуют ее структурный каркас. Большинство белков специфически связано с определенными участками рРНК. Некоторые белки входят в состав рибосом только во время биосинтеза белка. Различают два основных типа рибосом: эукариотические (с константами седиментации целой рибосомы — 80S14, малой субъединицы — 40S, большой — 60S) и прокариотические (соответст- венно 70S, 30S, 50S). В состав рибосом эукариот входит 4 молекулы рРНК и около 100 молекул белка, прокариот — 3 молекулы рРНК и около 55 молекул белка. В зависимости от локализации в клетке, различают
Во время биосинтеза белка рибосомы могут «работать» поодиночке или объединяться в комплексы — полирибосомы (полисомы). В таких комплексах они связаны друг с другом одной молекулой иРНК. Рибосомы эукариот образуются в ядрышке. Сначала на ядрышковой ДНК синтезируются рРНК, которые затем покрываются поступающими из цитоплазмы рибосомальными белками, расщепляются до нужных размеров и формируют субъединицы рибосом. Полностью сформированных рибосом в ядре нет. Объединение субъединиц в целую рибосому происходит в цитоплазме, как правило, во время биосинтеза белка. ЦитоскелетОдной из отличительных особенностей эукариотической клетки является наличие в ее цитоплазме скелетных образований в виде микротрубочек и пучков белковых волокон. Элементы цитоскелета, тесно связанные с наружной цитоплазматической мембраной и ядерной оболочкой, образуют сложные переплетения в цитоплазме.Цитоскелет образован микротрабекулярной системой, микротрубочками и микрофиламентами. Цитоскелет определяет форму клетки, участвует в движениях клетки, в делении и перемещениях самой клетки, во внутриклеточном транспорте органоидов и отдельных соединений. Микрофиламенты выполняют также функцию арматуры клетки. Микротрабекулярная системаМикротрабекулярная система представляет собой сеть из тонких фибрилл — трабекул (перекладин), в точках пересечения или соединения концов которых располагаются рибосомы.Микротрабекулярная система — динамичная структура: при изменении условий она может распадаться и вновь собираться. Функции микротрабекулярной решетки:
МикротрубочкиСодержатся во всех эукариотических клетках и представляют собой полые неразветвленные цилиндры, диаметр которых не превышает 30 нм, а толщина стенки — 5 нм. В длину они могут достигать нескольких микрометров. Легко распадаются и собираются вновь.Стенка микротрубочек в основном построена из спирально уложенных субъединиц белка тубулина. Считают, что роль матрицы (организатора микротрубочек) могут играть центриоли, базальные тельца жгутиков и ресничек, центромеры хромосом. Функции микротрубочек:
ЦентриолиЦентриоль представляет соб 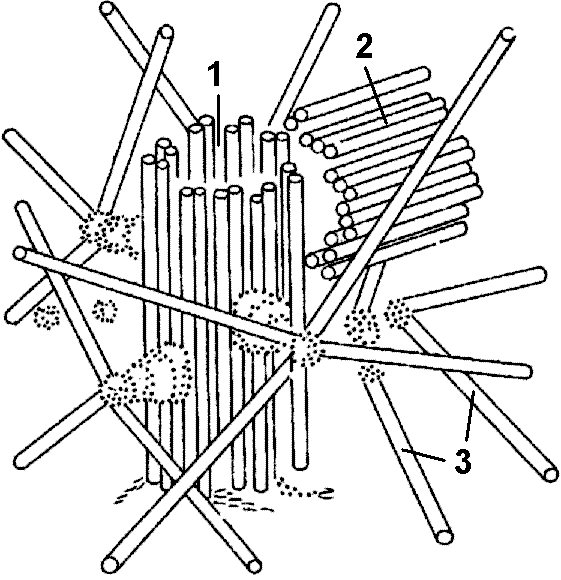 Рис. 288. Клеточный центр: 1. — материнская центриоль; 2 — дочерние центриоли; 3 — микротрубочки. ой цилиндр (длиной 0,3 мкм и диаметром 0,1 мкм), стенка которого образована девятью группами из трех слившихся микротрубочек (9 триплетов), соединенных между собой через определенные интервалы поперечными сшивками. Часто центриоли объединены в пары, где они расположены под прямым углом друг к другу. Если центриоль лежит в основании реснички или жгутика, то ее называют базальным тельцем. Почти во всех животных клетках имеется пара центриолей, являющихся срединным элементом центросомы, или клеточного центра (рис. 288). Перед делением центриоли расходятся к противоположным полюсам и возле каждой из них возникает дочерняя центриоль. От центриолей, расположенных на разных полюсах клетки, образуются микротрубочки, растущие навстречу друг другу. Они формируют митотическое веретено, способствующее равномерному распределению генетического материала между дочерними клетками, являются центром организации цитоскелета. Часть нитей веретена прикрепляется к хромосомам. В клетках высших растений клеточный центр центриолей не имеет. Центриоли относятся к самовоспроизводящимся органоидам цитоплазмы. Они возникают в результате дупликации уже имеющихся. Это происходит при расхождении центриолей. Незрелая центриоль содержит 9 одиночных микротрубочек; по-видимому, каждая микротрубочка является матрицей при сборке триплетов, характерных для зрелой центриоли. Реснички и жгутикиЭто волосовидные образования толщиной около 0,25 мкм, построенные из микротрубочек, у эукариот они покрытыресничек лишь длиной.Реснички и жгутики — органоиды движения клеток многих типов. Чаще всего реснички и жгутики встречаются у бактерий, некоторых простейших, зооспор и сперматозоидов. Жгутики бактерий имеют иное строение, чем жгутики эукариот. Реснички и жгутики образованы девятью сдвоенными микротрубочками, образующими стенку цилиндра, покрытого мембраной; в его центре находятся две одиночные микротрубочки. Такая структура типа 9+2 характерна для ресничек и жгутиков почти всех эукариотических организмов, от простейших до человека. Реснички и жгутики укреплены в цитоплазме базальными тельцами, лежащими в основании этих органоидов. Каждое базальное тельце состоит из девяти троек микротрубочек, в его центре микротрубочек нет. МикрофиламентыМикрофиламенты представлены нитями диаметром 6 нм, состоящими из белка актина, близкого к актину мышц. Актин составляет 10-15% общего количества белка клетки. В большинстве животных клеток образуется густая сеть из актиновых филаментов и связанных с ними белков под самой плазматической мембраной. Эта сеть придает поверхностному слою клетки механическую прочность и позволяет клетке изменять свою форму и двигаться.Помимо актина, в клетке обнаруживаются и нити миозина. Однако количество их значительно меньше. Благодаря взаимодействию актина и миозина происходит сокращение мышц. Микрофиламенты связаны с движением всей клетки либо ее отдельных структур внутри нее. В некоторых случаях движение обеспечивается только актиновыми филаментами, в других — актином вместе с миозином. ВключенияВключения — временные компоненты цитоплазмы, то возникающие, то исчезающие. Как правило, они содержатся в клетках на определенных этапах жизненного цикла. Специфика включений зависит от специфики соответствующих клеток тканей и органов. Включения встречаются преимущественно в растительных клетках. Они могут возникать в гиалоплазме, различных органеллах, реже в клеточной стенке.В функциональном отношении включения представляют собой:
Крахмальные зернаЭто наиболее распространенные включения растительных клеток. Крахмал запасается у растений исключительно в виде крахмальных зерен.Они образуются только в строме пластид живых клеток. В процессе фотосинтеза в зеленых листьях образуется ассимиляционный, или первичный крахмал. Ассимиляционный крахмал в листьях не накапливается и, быстро гидролизуясь до сахаров, оттекает в части растения, в которых происходит его накопление. Там он вновь превращается в крахмал, который называют вторичным. Вторичный крахмал образуется и непосредственно в клубнях, корневищах, семенах, то есть там, где он откладывается в запас. Тогда его называют запасным. Лейкопласты, накапливающие крахмал, называют амилопластами15. Особенно богаты крахмалом семена, подземные побеги (клубни, луковицы, корневища), паренхима проводящих тканей корней и стеблей древесных растений. Липидные каплиВстречаются практически во всех растительных клетках. Наиболее богаты ими семена и плоды. Жирные масла в виде липидных капель — вторая по значению (после крахмала) форма запасных питательных веществ. Семена некоторых растений (подсолнечник, хлопчатник и т.д.) могут накапливать до 40% масла от массы сухого вещества.Липидные капли, как правило, накапливаются непосредственно в гиалоплазме. Они представляют собой сферические тела обычно субмикроскопического размера. Липидные капли могут накапливаться и в лейкопластах, которые называют элайопластами. Белковые включенияБелковые включения образуются в различных органеллах клетки в виде аморфных или кристаллических отложений разнообразной формы и строения. Наиболее часто кристаллы можно встретить в ядре — в нуклеоплазме, иногда в перинуклеарном пространстве, реже в гиалоплазме, строме пластид, в расширениях цистерн ЭПР, матриксе пероксисом и митохондриях. В вакуолях встречаются как кристаллические, так и аморфные белковые включения. В наибольшем количестве кристаллы белка встречаются в запасающих клетках сухих семян в виде так называемых алейроновых16 зерен или белковых телец.Запасные белки синтезируются рибосомами во время развития семени и откладываются в вакуоли. При созревании семян, сопровождающемся их обезвоживанием, белковые вакуоли высыхают, и белок кристаллизуется. В результате этого в зрелом сухом семени белковые вакуоли превращаются в белковые тельца (алейроновые зерна). Кристаллы оксалата кальцияВключения, образующиеся в вакуолях, как правило, клеток листьев или коры. Это либо одиночные кристаллы, либо группы кристаллов разнообразной формы.Представляют собой конечные продукты жизнедеятельности клеток, образующиеся как приспособление для вывода из обмена веществ излишков кальция. Кроме оксалата кальция, в клетках могут накапливаться кристаллы карбоната кальция и кремнезема. 36.3. ЯдроН 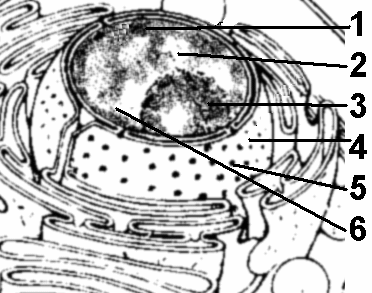 Рис. 289. Ядро: 1 — гетерохроматин; 2 — эухроматин; 3 — ядрышко; 4 — ядерная оболочка; 5 — пора ядерной оболочки; 6 — кариоплазма. аиболее важный компонент эукариотических клеток. Безъядерная клетка долго не существует. Ядро также не способно к самостоятельному существованию. Большинство клеток имеет одно ядро, но встречаются и многоядерные клетки (у ряда простейших, в скелетных мышцах позвоночных). Число ядер может достигать нескольких десятков. Некоторые высокоспециализированные клетки утрачивают ядро (эритроциты млекопитающих и клетки ситовидных трубок у покрытосеменных растений). Форма и размер ядер клеток разнообразны. Обычно ядро имеет диаметр от 3 до 10 мкм. Форма в большинстве случаев связана с формой клетки, но часто отличается от нее. Как правило, имеет шаровидную или овальную форму, реже может быть сегментированным, веретеновидным. Главными функциями ядра являются:
В состав ядра входят (рис. 289):
Ядерная оболочкаЯдро отграничено от остальной цитоплазмы ядерной оболочкой, состоящей из двух мембран типичного строения. Между мембранами имеется узкая щель, заполненная полужидким веществом, — перинуклеарное пространство. В некоторых местах обе мембраны сливаются друг с другом, образуя ядерные поры, через которые происходит обмен веществ между ядром и цитоплазмой. Из ядра в цитоплазму и обратно вещества могут попадать также вследствие отшнуровывания впячиваний и выростов ядерной оболочки.Несмотря на активный обмен веществ, ядерная оболочка обеспечивает различия в химическом составе ядерного сока и цитоплазмы, что необходимо для нормального функционирования ядерных структур. Наружная ядерная мембрана со стороны, обращенной в цитоплазму, покрыта рибосомами, придающими ей шероховатость, внутренняя мембрана гладкая. Ядерная оболочка — часть мембранной системы клетки. Выросты внешней ядерной мембраны соединяются с каналами эндоплазматической сети, образуя единую систему сообщающихся каналов. КариоплазмаКариоплазма — внутреннее содержимое ядра. Представляет собой гелеобразный матрикс, в котором располагаются хроматин и одно или несколько ядрышек. В состав ядерного сока входят различные белки (в том числе ферменты ядра), свободные нуклеотиды, а также продукты жизнедеятельности ядрышка и хроматина.ЯдрышкоТретья характерная для ядра клетки структура — ядрышко, представляющее собой округлое плотное тельце, погруженное в ядерный сок. Количество ядрышек зависит от функционального состояния ядра и может колебаться от 1 до 5–7 и более (даже в одной и той же клетке). Ядрышки обнаруживаются только в неделящихся ядрах, во время митоза они исчезают, а после завершения деления возникают вновь. Ядрышко не является самостоятельной структурой ядра. Оно образуется в результате концентрации в определенном участке кариоплазмы участков хромосом, несущих информацию о структуре рРНК. Эти участки хромосом называют ядрышковыми организаторами. Они содержат многочисленные копии генов, кодирующих рРНК. Поскольку в ядрышке интенсивно идет процесс синтеза рРНК и формирование субъединиц рибосом, можно говорить, что ядрышко — это скопление рРНК и рибосом на разных этапах формирования.ХроматинХроматином называют глыбки, гранулы и сетевидные структуры ядра, интенсивно окрашивающиеся некоторыми красителями и отличающиеся по форме от ядрышка. Хроматин представляет собой молекулы ДНК, связанные с белками — гистонами. В зависимости от степени спирализации различают:
Хроматин представляет собой форму существования генетического материала в неделящихся клетках и обеспечивает возможность удвоения и реализации заключенной в нем информации. В процессе деления клеток ДНК спирализуется и хроматиновые структуры образуют хромосомы. Хромосомами называются постоянные компоненты ядра клетки, имеющие особую организацию, функциональную и морфологическую специфичность, способные к самовоспроизведению и сохранению свойств на протяжении всего онтогенеза. Хромосомы — плотные, интенсивно окрашивающиеся структуры (отсюда и их название). Впервые они были обнаружены Флемингом (1882) и Страсбургером (1884). Термин “хромосома” предложил Вальдейер в 1888 г. Функции хромосом:
Главными химическими компонентами хромосом являются ДНК (40%) и белки (60%). Основным компонентом хромосом является ДНК, так как в ее молекулах закодирована наследственная информация, белки же выполняют структурную и регуляторную функции. Различают две основные формы хромосом, приуроченные к определенным фазам и периодам митотического цикла:
Реорганизация хромосом происходит в процессе спирализации (конденсации) или деспирализации (деконденсации). В неделящихся клетках хромосомы находятся в деконденсированном состоянии, так как только в этом случае может считываться заложенная в них информация. Во время деления клетки спирализацией достигается плотная упаковка наследственного материала, что важно для перемещения хромосом во время митоза. Общая длина ДНК клетки человека — 2 метра, совокупная же длина всех хромосом клетки — всего лишь 150 мкм. Все сведения о хромосомах получены при изучении метафазных хромосом. Каждая метафазная хромосома состоит из двух хроматид, являющихся дочерними хромосомами (рис. 290). В процессе митоза они разойдутся в дочерние клетки и станут самостоятельными хромосомами. Хроматиды— сильно спирализованные идентичные молекулы ДНК, образо- в 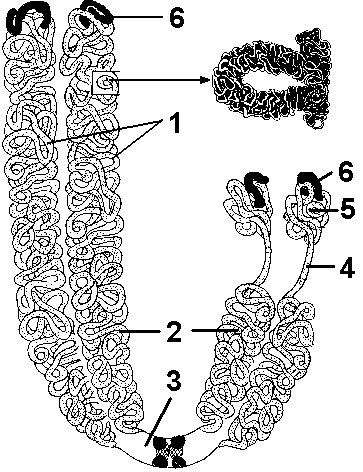 Рис. 290. Строение метафазной хромосомы: 1 — дочерние хроматиды; 2 — плечи; 3 — первичная перетяжка; 4 — вторичная перетяжка; 5 — спутник; 6 — теломеры. авшиеся в результате репликации. Они соединяются между собой в области первичной перетяжки (центромеры), к которой прикрепляются нити веретена деления. Фрагменты, на которые первичная перетяжка делит хромосому, называются плечами, а концы хромосомы — теломерами. Теломеры предохраняют концы хромосом от слипания, способствуя тем самым сохранению целостности хромосом. В зависимости от места положения центромеры различают (рис. 291):
Некоторые хромосомы имеют вторичные перетяжки, возникающие в участках неполной конденсации хроматина. Они являются ядрышковыми организаторами. Иногда вторичная перетяжка очень длинная и отделяет от основного тела хромосомы небольшой участок — спутник. Такие хромосомы называют спутничными. Хромосомы обладают индивидуальными особенностями: длиной, положением центромеры, формой. Каждый вид живых организмов имеет в своих клетках определенное и постоянное число хромосом. Хромосомы ядра одной клетки всегда парные. Каждая пара образована хромосомами, имеющими одинаковый размер, форму, положение первичной и вторичной перетяжек. Такие хромосомы называют гомологичными. У человека 23 пары гомологичных хромосом. Совокупность количественных (число и размеры) и качественных (форма) признаков хромосомного набора соматической клетки называется кариотипом. Число хромосом в кариотипе всегда четное, так как соматические клетки имеют две одинаковые по форме и размеру хромосомы: одну — отцовскую, другую — материнскую. Хромосомный набор всегда видоспецифичен, то есть, характерен только для данного вида организмов. Если в ядрах клеток хромосомы образуют гомологичные пары, то такой набор хромосом называют диплоидным (двойным) и обозначают — 2n. Количество ДНК, соответствующее диплоидному набору хромосом, обозначают 2с. Диплоидный набор хромосом характерен для соматических клеток. В ядрах половых клеток каждая хромосома представлена в единственном числе. Такой набор хромосом называют гаплоидным (одинарным) и обозначают — n. У человека диплоидный набор содержит 46 хромосом, а гаплоидный — 23. |