Пособие по биологии. Пособие по биологии для поступающих в вузы Авторы Пименов А. В., Гончаров О. В
 Скачать 34.42 Mb. Скачать 34.42 Mb.
|

Глава 37. Обмен веществ37.1. Общая характеристикаВажнейшее свойство живых организмов — обмен веществ. Любой живой организм — открытая система, которая потребляет из окружающей среды различные вещества и использует их в качестве строительного материала, или как источник энергии и выделяет в окружающую среду продукты жизнедеятельности и энергию. Совокупность реакций обмена веществ, протекающих в организме, называется метаболизмом, состоящим из взаимосвязанных реакций ассимиляции (пластического обмена, анаболизма) и реакций диссимиляции (энергетического обмена, катаболизма). Ассимиляция — совокупность реакций биосинтеза, протекающих в клетке, диссимиляция — совокупность реакций распада и окисления высокомолекулярных веществ, идущих с выделением энергии. Эти две группы реакций взаимосвязаны, реакции биосинтеза невозможны без энергии, которая выделяется в реакциях энергетического обмена, реакции диссимиляции не идут без ферментов, образующихся в реакциях пластического обмена. Для поддержания различных процессов жизнедеятельности, например: для движения, для биосинтеза различных органических соединений; для поглощения веществ — организму необходима энергия. Одна группа организмов (фотоавтотрофы) использует солнечную энергию,вторая группа (хемоавтотрофы) использует энергию, выделяющуюся при окислении неорганических веществ, третья группа организмов (хемогетеротрофы) окисляет органические вещества и использует выделяющуюся при этом энергию. Если организмы в зависимости от условий ведут себя как авто– либо как гетеротрофы, то их называют миксотрофами. Метаболизм авто– и гетеротрофов различается. В качестве источника углерода автотрофы используют неорганические вещества (СО2), а гетеротрофы — органические. Различны и источники энергии: у автотрофов — энергия солнечного света или энергия, выделяющаяся при окислении неорганических соединений, у гетеротрофов — энергия окисления органических веществ. Важнейшими реакциями пластического обмена являются реакции биосинтеза белков и реакции фотосинтеза, реакции энергетического обмена рассмотрим на примере окисления углеводов. 37.2. Биосинтез белков, код ДНК, транскрипцияВ каждой клетке синтезируется несколько тысяч различных белковых молекул. Белки недолговечны, время их существования ограничено, после чего они разрушаются. Способность синтезировать строго определенные белки закреплена наследственно, информация о последовательности аминокислот в белковой молекуле закодирована в виде последовательности нуклеотидов в ДНК. В геноме человека менее 100 000 генов, которые находятся в 23 хромосомах. Одна хромосома содержит несколько тысяч генов, которые располагаются в линейном порядке в определенных участках хромосомы — локусах. Ген — участок молекулы ДНК, кодирующий первичную последовательность аминокислот в полипептиде или последовательность нуклеотидов в молекулах транспортных и рибосомальных РНК. Итак, последовательность нуклеотидов каким-то образом кодирует последовательность аминокислот. Все многообразие белков образовано из 20 различных аминокислот, а нуклеотидов в составе ДНК — 4 вида. Если предположить, что один нуклеотид кодирует одну аминокислоту, то 4 нуклеотидами можно закодировать 4 аминокислоты, если 2 нуклеотида кодируют одну аминокислоту, то количество кодируемых кислот возрастает до 42 — 16. Значит, код ДНК должен быть триплетным. Было доказано, что именно три нуклеотида кодируют одну аминокислоту, в этом случае можно будет закодировать 43 — 64 аминокислоты. А так как аминокислот всего 20, то некоторые аминокислоты должны кодироваться несколькими триплетами. В настоящее время известны следующие свойства генетического кода:
Таблица 7. Генетический код
Первый нуклеотид в триплете — один из четырех левого вертикального ряда, второй — один из верхнего горизонтального ряда, третий — из правого вертикального. В начале 50 гг. Ф. Крик сформулировал центральную догму молекулярной биологии: ДНКРНКбелок. Информация о белке находится на ДНК, на матрице ДНК синтезируется иРНК, которая является матрицей для синтеза белковой молекулы. Матричный синтез позволяет очень точно и быстро синтезировать макромолекулы полимеров, состоящие из огромного количества мономеров. С реакциями матричного синтеза мы встречались при удвоении молекулы ДНК, синтез иРНК (транскрипция) и синтез молекулы белка на иРНК (трансляция) — также реакции матричного синтеза. Транскрипция. В соответствии с принятыми соглашениями, начало гена на схемах изображают слева (рис. 292). У некодирующей цепи молекулы ДНК левый конец 5', правый 3'; у кодирующей, матричной, с которой идет транскрипция — противоположное направление. Фермент, отвечающий за синтез иРНК, РНК-полимераза, присоединяется к промотору, который находится на 3'-конце матричной цепи ДНК и движется всегда от 3' к 5' концу. Промотор — определенная последовательность нуклеотидов, к которой может присоединиться фермент РНК-полимераза. Необходим для того, чтобы синтез иРНК был начат строго в начале гена. Из свободных рибонуклеозидтрифосфатов (АТФ, УТФ, ГТФ, ЦТФ), комплементарных нуклеотидам ДНК, РНК-полимераза образует иРНК. 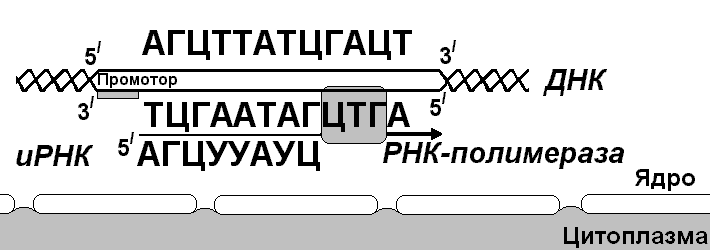 Рис. 292. Транскрипция, схема образования иРНК на матрице ДНК. Энергия для синтеза иРНК содержится в макроэргических связях рибонуклеозидтрифосфатов. Период полураспада мРНК исчисляется часами и даже сутками, т.е. они стабильны. Транскрипция и трансляция разобщены в пространстве и во времени, транскрипция протекает в ядре и в одно время, трансляция происходит в цитоплазме и совсем в другое время. Для транскрипции необходимы: 1 — кодирующая цепь ДНК, матрица; 2 — ферменты, один из них РНК-полимераза; 3 — рибонуклеозидтрифосфаты. 37.3. ТрансляцияТрансляция — процесс образования полипептидной цепи на матрице иРНК, или преобразование информации, закодированной в виде последовательности нуклеотидов иРНК, в последовательность аминокислот в полипептиде. Синтез белковых молекул происходит в цитоплазме или на шероховатой эндоплазматической сети. В цитоплазме синтезируются белки для собственных нужд клетки, белки, синтезируемые на ЭПС, транспортируются по ее каналам в комплекс Гольджи и выводятся из клетки. Для транспорта аминокислот к рибосомам используются транспортные РНК, тРНК. В клетке их более 30 видов, длина тРНК от 76 до 85 нуклеотидных остатков, они имеют третичную структуру за счет спаривания комплементарных нуклеотидов и по форме напоминают лист клевера. В тРНК различают антикодоновую петлю и акцепторный участок. На верхушке антикодоновой петли каждая тРНК имеет антикодон, комплементарный кодовому триплету определенной аминокислоты, а акцепторный участок на 3'-конце способен с помощью фермента аминоацил-тРНК-синтетазы присоединить именно эту аминокислоту (с затратой АТФ). Таким образом, у каждой аминокислоты есть свои тРНК и свои ферменты, присоединяющие аминокислоту к тРНК. Двадцать видов аминокислот кодируются 61 кодовым триплетом, теоретически может иметься 61 вид тРНК с соответствующими антикодонами, то есть у одной аминокислоты может быть несколько тРНК. Установлено существование нескольких тРНК, способных связываться с одним и тем же кодоном (последний нуклеотид в антикодоне не всегда важен). Обнаружено всего более 30 различных тРНК (рис. 293). 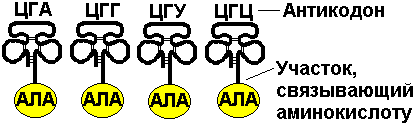 Рис. 293. Аланиновые тРНК, чьи антикодоны комплементарны кодовым триплетам ГЦУ, ГЦЦ, ГЦА, ГЦГ. Органоиды, отвечающие за синтез белков в клетке — рибосомы. У эукариот рибосомы находятся в некоторых органоидах — митохондриях и пластидах (70-S рибосомы) и в цитоплазме: в свободном виде и на мембранах эндоплазматической сети (80-S рибосомы). Малая субчастица рибосомы отвечает за генетические, декодирующие функции; большая — за биохимические, ферментативные. В малой субъединице рибосомы различают функциональный центр (ФЦР) с двумя участками — пептидильным (Р-участок) и аминоацильным (А-участок). В ФЦР может находиться шесть нуклеотидов иРНК, три в пептидильном и три в аминоацильном участках. С 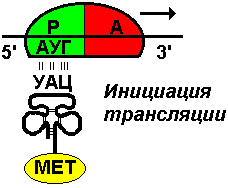 Рис. 294. Инициация трансляции. интез белка начинается с того момента, когда к 5'-концу иРНК присоединяется малая субъединица рибосомы, в Р-участок которой заходит метиониновая тРНК с аминокислотой метионин (рис. 294). Любая полипептидная цепь на N-конце сначала имеет метионин, который в дальнейшем чаще всего отщепляется. Синтез полипептида идет от N-конца к С-концу, то есть пептидная связь образуется между карбоксильной группой первой и аминогруппой второй аминокислоты. Затем происходит присоединение большой субчастицы рибосомы и в А-участок поступает вторая тРНК, чей антикодон комплементарно спаривается с кодоном иРНК, находящимся в А-участке. Пептидилтрансферазный центр большой субчастицы катализирует образование пептидной связи между метионином и второй аминокислотой. Отдельного фермента, катализирующего образование пептидных связей, не существует. Энергия для образования пептидной связи поставляется за счет гидролиза ГТФ (рис. 295). 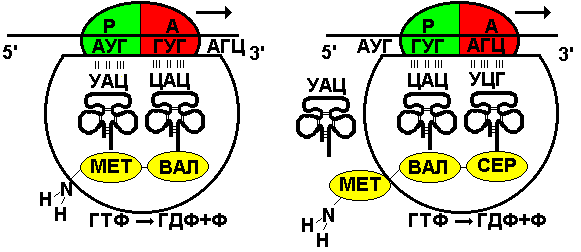 Рис. 295. Этапы трансляции. Как только образовалась пептидная связь, метиониновая тРНК отсоединяется от метионина, а рибосома передвигается на следующий кодовый триплет иРНК, который оказывается в А-участке рибосомы, а метиониновая тРНК выталкивается в цитоплазму. На один цикл расходуется 2 молекулы ГТФ. Затем все повторяется, образуется пептидная связь между второй и третьей аминокислотами. Трансляция идет до тех пор, пока в А-участок не попадает стоп-кодон (УАА, УАГ или УГА), с которым связывается особый белковый фактор освобождения, белковая цепь отделяется от тРНК и покидает рибосому. Происходит диссоциация, разъединение субчастиц рибосомы. Многие белки синтезируются в виде предшественников, содержащих ЛП — лидерную последовательность (15 — 25 аминокислотных остатков на N-конце, «паспорт белка»). ЛП определяют места назначения белков, "направление" белка (в ядро, в митохондрию, в пластиды, в комплекс Гольджи). Затем протеолитические ферменты отщепляют ЛП. Скорость передвижения рибосомы по иРНК — 5–6 триплетов в секунду, на синтез белковой молекулы, состоящей из сотен аминокислотных остатков, клетке требуется несколько минут. Первым белком, синтезированным искусственно, был инсулин, состоящий из 51 аминокислотного остатка. Потребовалось провести 5000 операций, в работе принимали участие 10 человек в течение трех лет. Таким образом, для трансляции необходимы: 1 — иРНК, кодирующая последовательность аминокислот в полипептиде; 2 — рибосомы, декодирующие иРНК и образующие полипептид; 3 — тРНК, транспортирующие аминокислоты в рибосомы; 4 — энергия в форме АТФ и ГТФ для присоединения аминокислот к рибосоме и для работы рибосомы; 5 — аминокислоты, строительный материал; 6 — ферменты (аминоацил-тРНК-синтетазы и др.). 37.4. Фотосинтез, хемосинтезРастения относятся к фотоавтотрофным организмам, способным к синтезу органических веществ за счет энергии солнечного света. Фотосинтез — процесс образования органических веществ из углекислого газа и воды за счет энергии света, при этом выделяется кислород. 6СО2 + 6Н2О + Q света= С6Н12О6 + 6О2 Главным органом фотосинтеза является лист, в клетках которого имеются специализированные органоиды, ответственные за фотосинтез — хлоропласты. В хлоропластах находятся фотосинтетические пигменты: хлорофилл а — у всех фотосинтезирующих растений и синезеленых (формы 670, 680, 690, 700); хлорофилл b — вспомогательный пигмент; хлорофилл с — у бурых водорослей вместо хлорофилла b. Поглощают в основном синие и красные лучи, отражают зеленые — отсюда и зеленая окраска растений. Кроме хлорофиллов в мембранах тилакоидов имеются сопровождающие пигменты фотосинтеза — каротиноиды — желтые, оранжевые или красные. (Поглощают сине-зеленые лучи). Свет с длиной волны меньше 290 нм, жесткий ультрафиолет (ЖУФ) отражается озоновым экраном, значительная часть инфракрасных волн поглощается водяными парами и углекислотой атмосферы (рис. 296). Энергия фотона всегда обратно пропорциональна длине волны, фотоны фиолетового света несут почти вдвое больше энергии, чем фотоны красного света. 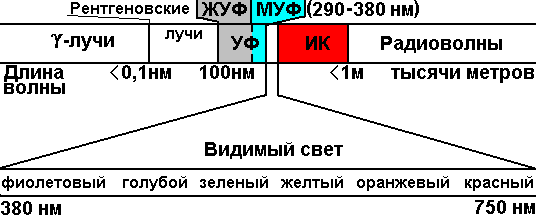 Рис. 296. Длины волн солнечного излучения В процессе фотосинтеза различают две фазы: световую и темновую. Световая фаза происходит только на свету в мембранах тилакоидов. Тилакоид представляет собой уплощенный мешочек, образованный мембранами, содержащими молекулы хлорофилла, белки цепи переноса электронов и особые ферменты — АТФ-синтетазы. Молекулы хлорофилла в мембранах тилакоидов организованы в фотосистемы, содержащие около 300 молекул. Более древняя фотосистема появилась у фотосинтезирующих бактерий — фотосистема-1, она способна отбирать электроны и протоны у сероводорода, при этом не происходит выделения О2: СО2 + 2Н2S + световая энергия (СН2О) + Н2О + 2S У сине-зеленых водорослей, а затем у всех настоящих растений, кроме фотосистемы-1, появляется фотосистема-2, способная разлагать воду с выделением О2, способная отбирать электроны у водорода воды: СО2 + 2Н2О + световая энергия (СН2О) + Н2О + О2 Под действием энергии кванта света электроны хлорофилла возбуждаются, покидают молекулу и попадают на внешнюю сторону мембраны тилакоида, которая в итоге заряжается отрицательно. Окисленные молекулы хлорофилла восстанавливаются, разлагая воду — отбирая электроны у водорода воды с помощью особого фермента, связанного с фотосистемой-2. Кислород при этом удаляется во внешнюю среду, а протоны накапливаются в полости тилакоида, так называемом «протонном резервуаре». 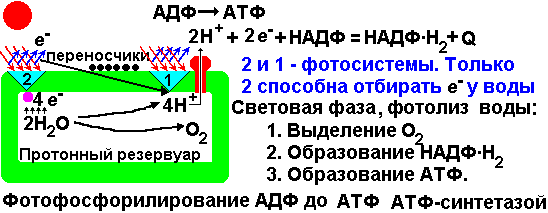 Рис. 297. Фотолиз воды в световую фазу фотосинтеза в тилакоиде. Когда разность потенциалов между наружной и внутренней сторонами мембраны тилакоида достигает 200 мВ, срабатывает фермент АТФ-синтетаза, протоны проталкиваются через его канал и происходит фосфорилирование АДФ до АТФ, а атомарный водород идет на восстановление специфического переносчика НАДФ+ (никотинамидадениндинуклеотидфосфата) до НАДФ·Н2 (рис. 297). Т 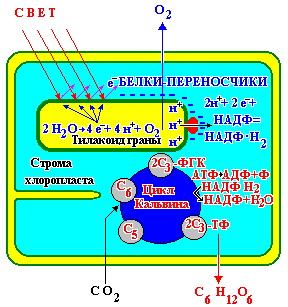 Рис. 298. Фотосинтез аким образом, в световую фазу происходит фотолиз воды, который сопровождается тремя важнейшими процессами: 1 — образованием кислорода; 2 — образованием АТФ; 3 — образованием НАДФ·Н2. Темновая фаза протекает в другое время и в другом месте — в строме хлоропласта. Для ее реакций не нужна энергия света. Происходит фиксация углекислого газа, содержащегося в воздухе, причем акцептором углекислого газа является пятиуглеродный сахар рибулозобисфосфат (рис. 298). Мелвин Кальвин, лауреат Нобелевской премии, показал, как происходит образование углеводов в темновую фазу фотосинтеза. Фермент РиБФ-карбоксилаза (самый распространенный в мире фермент) катализирует реакцию карбоксилирования рибулозобисфосфата с образованием 6-углеродного соединения. Затем происходит цикл реакций, в которых через ряд промежуточных продуктов происходит образование глюкозы и регенерация рибулозобисфосфата. В этих реакциях используется энергия АТФ и НАДФ·Н2, образованных в световую фазу, цикл этих реакций получил название "цикл Кальвина". 6СО2 + 24Н + АТФ С6Н12О6 + 6Н2О Кроме глюкозы, в процессе фотосинтеза образуются другие мономеры сложных органических соединений — аминокислоты, глицерин и жирные кислоты, нуклеотиды. Благодаря фотосинтезу, ежегодно из атмосферы поглощаются миллиарды тонн углекислого газа, выделяются миллиарды тонн кислорода, фотосинтез является основным источником образования органических веществ. Из кислорода образуется озоновый слой, защищающий живые организмы от коротковолновой ультрафиолетовой радиации. При фотосинтезе зеленый лист использует лишь около 1% падающей на него солнечной энергии, продуктивность составляет около 1 г органического вещества на 1 м.2 поверхности в час. Кроме процесса фотосинтеза, в листьях протекает и противоположный процесс — дыхание, при котором поглощается кислород и выделяется углекислый газ. Но при фотосинтезе выделяется кислорода в 20 — 30 раз больше, чем поглощается при дыхании. Кроме фотоавтотрофных организмов, существует группа хемоавтотрофных организмов, использующих неорганический источник углерода и энергию окисления неорганических соединений. К ним относятся: нитрифицирующие бактерии, окисляющие аммиак нитратов (Nitrosomonas): NH3 → HNO2 → HNO3; железобактерии, окисляющие закисное железо в окисное: Fe2+ → Fe3+; серобактерии, окисляющие сероводород до серы или до серной кислоты: Н2S+1/2O2 → S+H2O; H2S+2O2 → H2SO4. 37.5. Энергетический обменОрганические вещества пищи являются основным источником не только материи, но и энергии для жизнедеятельности клеток организма. При образовании сложных органических молекул была затрачена энергия, потенциально она находится в форме образованных химических связей. В результате реакций энергетического обмена происходит окисление сложных молекул до более простых и разрушение химических связей, при этом происходит высвобождение энергии. Биологическое окисление в клетках происходит с участием О2: А + О2 АО2 и без его участия, за счет переноса атомов водорода или электронов от одного вещества к другому: АН2 + В А + ВН2, где вещество А окисляется за счет вещества В; Fe2+ Fe3+ + e-, где двухвалентное железо окисляется до трехвалентного. Процесс энергетического обмена можно разделить на три этапа: на первом этапе происходит пищеварение, то есть сложные органические молекулы расщепляются до мономеров, на втором происходит бескислородное окисление этих мономеров — гликолиз, и на последнем этапе происходит окисление с участием кислорода в митохондриях. Подготовительный этап. Под действием ферментов пищеварительного тракта или ферментов лизосом белковые молекулы расщепляются до аминокислот, жиры — до глицерина и карбоновых кислот, углеводы — до глюкозы, нуклеиновые кислоты — до нуклеотидов. Вся энергия при этом рассеивается в виде тепла. Гликолиз, или бескислородное окисление. Окисление глюкозы в клетках без участия кислорода происходит путем дегидрирования, акцептором Н служит кофермент НАД+. Реакции протекают в цитоплазме, глюкоза с помощью 10 ферментативных реакций превращается в 2 молекулы ПВК — пировиноградной кислоты и образуется восстановленная форма переносчика водорода НАД·Н2 никотинамидаденин-динуклеотида. При этом образуется 200 кДж энергии, 120 рассеивается в форме тепла, 80 кДж запасается в форме 2 моль АТФ: С6Н12О6 + 2АДФ + 2Н3РО4 + 2НАД+ 2 С3Н4О3 + 2АТФ + 2Н2О + 2НАД·Н2 Дальнейшая судьба ПВК зависит от присутствия О2 в клетке, если О2 нет, происходит анаэробное дыхание, причем у дрожжей и растений происходит спиртовое брожение, при котором сначала происходит образование уксусного альдегида, а затем этилового спирта: I. С3Н4О3 СО2 + СН3СОН (уксусный альдегид) II. СН3СОН + НАД·Н2 С2Н5ОН + НАД+ У животных и некоторых бактерий при недостатке О2 происходит молочнокислое брожение с образованием молочной кислоты: С3Н4О3 + НАД·Н2 С3Н6О3 + НАД+ Третий этап энергетического обмена — кислородное окисление, или дыхание, происходит в митохондриях. Пировиноградная кислота проникает в митохондрии, происходит ее дегидрирование (отщепление водорода) и декарбоксилирование (отщепление углекислого газа) с образованием двухуглеродной ацетильной группы, которая вступает в цикл реакций, получивших название реакций цикла Кребса (рис. 299). Здесь происходит дальнейшее окисление, связанное с дегидрированием и декарбоксилированием. В результате на каждую разрушенную моль ПВК из митохондрии удаляется 3 моль СО2, образуется 5 пар атомов водорода, связанных с переносчиками (4 НАДН2, ФАДН2), а также моль АТФ. Суммарная реакция гликолиза и разрушения ПВК в митохондриях до водорода и углекислого газа выглядит следующим образом: С6Н12О6 + 6Н2О 6СО2 + 4АТФ + 12Н2 2АТФ образуются при гликолизе, две — в цикле Кребса; 2 пары атомов (2НАД·Н2)образовались при гликолизе, 10 пар — в цикле Кребса. 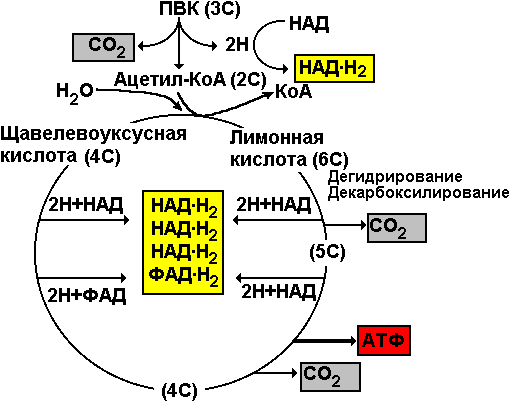 Рис.299. Цикл Кребса. Последним этапом является окисление пар атомов водорода с участием О2 до Н2О с одновременным фосфорилированием АДФ до АТФ. Этот процесс происходит на внутренней мембране митохондрий. Водород передается по трем большим ферментным комплексам дыхательной цепи (флавопротеин, кофермент Q, цитохромы), расположенным во внутренней мембране митохондрий. У водорода отбираются электроны, а протоны закачиваются в межмембранное пространство митохондрий, в «протонный резервуар». Внутренняя мембрана непроницаема для ионов водорода. Электроны передаются по ферментам дыхательной цепи на цитохромоксидазу. Когда разность потенциалов на внешней и внутренней стороне внутренней мембраны достигает 200 мВ, протоны (12Н2) проходят через канал фермента АТФ-синтетазы и с помощью цитохромоксидазы происходит восстановление кислорода до воды (12Н2О) с выделением энергии, часть которой (55%) запасается в форме 34АТФ (рис. 300). 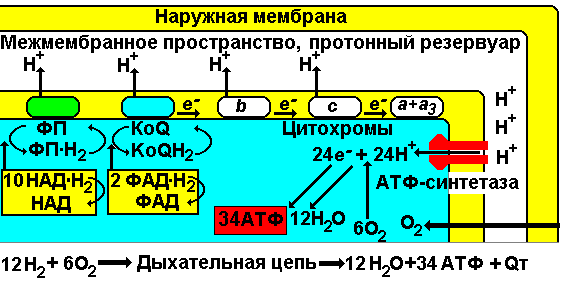 Рис. 300. Дыхательная цепь и АТФ-синтетаза. Суммарная реакция энергетического обмена выглядит так: | ||||||||||||||||||||||||||||||||||