Постсинаптическое торможение docx. Постсинаптическое торможение
 Скачать 193.29 Kb. Скачать 193.29 Kb.
|

|
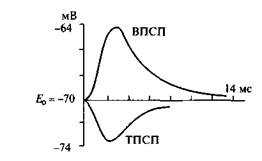
Постсинаптическое торможение открыл Дж. Экклс(1952) при регистрации потенциалов мотонейронов спинного мозга у кошки во время раздражения мышечных афферентов. При этом оказалось, что в мотонейронах мышцы - антагониста регистрируется не деполяризация, а гиперполяризационныйпостсинаптический потенциал, уменьшающий возбудимость мотонейрона, угнетающий его способность реагировать на возбуждающие влияния. По этой причине вызванный гиперполяризационный потенциал был назван тормозным постсинаптическим потенциалом – ТПСП .Амплитуда ТПСП 1-5 мВ, он способен суммироваться, более мощный афферентный залп вызывает возрастание амплитуды ТПСП. Рис 1
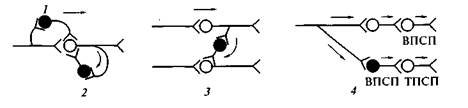
Рис.1. Возбуждающий (ВПСП) и тормозной (ТПСП) постсинаптические потенциалы Механизм постсинаптического торможения. ТПСП уменьшает возбудимость клетки, т.е. увеличивает пороговый потенциал (ДV), так как Екр(критический уровень деполяризации – КУД) остается на прежнем уровне, а мембранный потенциал (Е0)возрастает. ТПСП возникает под влиянием аминокислоты глицина, а также гамма-аминомасляной кислоты (ГАМК ). В спинном мозге глицин выделяется особыми тормозными клетками (клетками Реншоу)в синапсах, образуемых этими клетками на мембране нейрона-мишени. Действуя на ионотропный рецептор постсинаптической мембраны, глицин увеличивает ее проницаемость для С1-, при этом С1- поступает в клетку согласно концентрационному градиенту и вопреки электрическому градиенту, в результате чего развивается гиперполяризация. В среде, обедненной хлором, тормозная роль глицина не реализуется. Ареактивность нейрона к возбуждающим импульсам является следствием алгебраической сум-мации ТПСП и ВПСП, в связи с чем в зоне аксонного холмика не происходит выведения, мембранный потенциал не достигает критического уровня. При действии ГАМК на постсинаптическую мембрану ТПСП развивается в результате входа С1- в клетку или выхода К+ из клетки. Концентрационные градиенты ионов К+ в процессе развития торможения нейронов поддерживаются №+/К+-помпой, ионов С1- - С1--помпой. Разновидности постсинаптического торможения. Обычно выделяют возвратное, латеральное, параллельное и прямое (реципроктное) постсинаптическое торможение.Имеются и другие варианты классификаций. Некоторые авторы называют только два вида торможения: возвратное и прямое (трактуется по-разному). В реальности вариантов торможения больше: они определяются множеством связей различных нейронов, в частности их коллатералей. Возвратное постсинаптическое торможение – это такое торможение, когда тормозные вставочные нейрону действуют на те же нервные клетки, которые их активируют. В этом случае развивающееся торможение бывает тем глубже, чем сильнее было предшествующее возбуждение. Типичным примером возвратного постсинаптического торможения является торможение в мотонейронах спинного мозга. Как следует из рис.2, мотонейроны посылают коллатерали к тормозным вставочным нейронам, аксоны которых, в свою очередь, образуют синапсы на тех же мотонейронах, которые возбуждают тормозную клетку Реншоу. Такая тормозная цепь называется торможением Реншоу (в честь ученого, который ее открыл), а тормозные вставочные нейроны в этой цепи – клетками Реншоу. Это торможение обеспечивает, например, поочередное сокращение и расслабление скелетных мышц-сгибателей и разгибателей, что необходимо для координации движений конечностей при ходьбе. Сама клетка Реншоу возбуждается под влиянием ацетилхолина, воздействующего на
Рис. 5. Разновидности постсинаптического торможения: 2 — возвратное; 3 — латеральное; 4 — прямое; Подобную роль может выполнять и параллельное торможение, когда возбуждение блокирует само себя за счет дивергенции по коллатерали с включением тормозной клетки на своем пути и возвратом импульсов к нейрону, который активировался этим же возбуждением. Латеральное постсинаптическое торможение графически представлено на рис.2 (3). Тормозные вставочные нейроны соединены таким образом, что они активируются импульсами от возбужденного центра и влияют на соседние клетки с такими же функциями. В результате в этих соседних клетках развивается очень глубокое торможение, называемое латеральным, так как образующаяся зона торможения находится «сбоку» по отношению к возбужденному нейрону и инициируется им. Латеральное торможение играет особенно важную роль в афферентных системах: оно может образовать тормозную зону, которая окружает возбуждающие нейроны. Примером прямого торможения может служить реципрокное торможение,вызывающее угнетение центра-антагониста. Например, при раздражении кожных рецепторов возникает защитный сгибательный рефлекс: центр сгибания возбужден, а центр разгибания заторможен. В этом случае возбуждающие импульсы поступают к центру мышцы-сгибателя, а через тормозную клетку Реншоу – к центру мышцы-антагониста, т.е. разгибателю, что предотвращает ее сокращение. Если бы возбуждались одновременно центры мышц-сгибателей и мышц-разгибателей, сгибание конечности в суставе было бы невозможным. Основными медиаторами вызывающими постсинаптическое торможение являются : Гамма-аминомасляная кислота (ГАМК) — важнейший тормозной нейромедиатор центральной нервной системы человека и млекопитающих. Глицин— как нейромедиаторная аминокислота, проявляет двоякое действие. Глициновые рецепторы имеются во многих участках головного мозга и спинного мозга. Связываясь с рецепторами, глицин вызывает «тормозящее» воздействие на нейроны, уменьшает выделение из нейронов «возбуждающих» аминокислот, таких как глутамат, и повышает выделение ГАМК. В спинном мозге глицин приводит к торможению мотонейронов, что позволяет использовать глицин в неврологической практике для устранения повышенного мышечного тонуса. 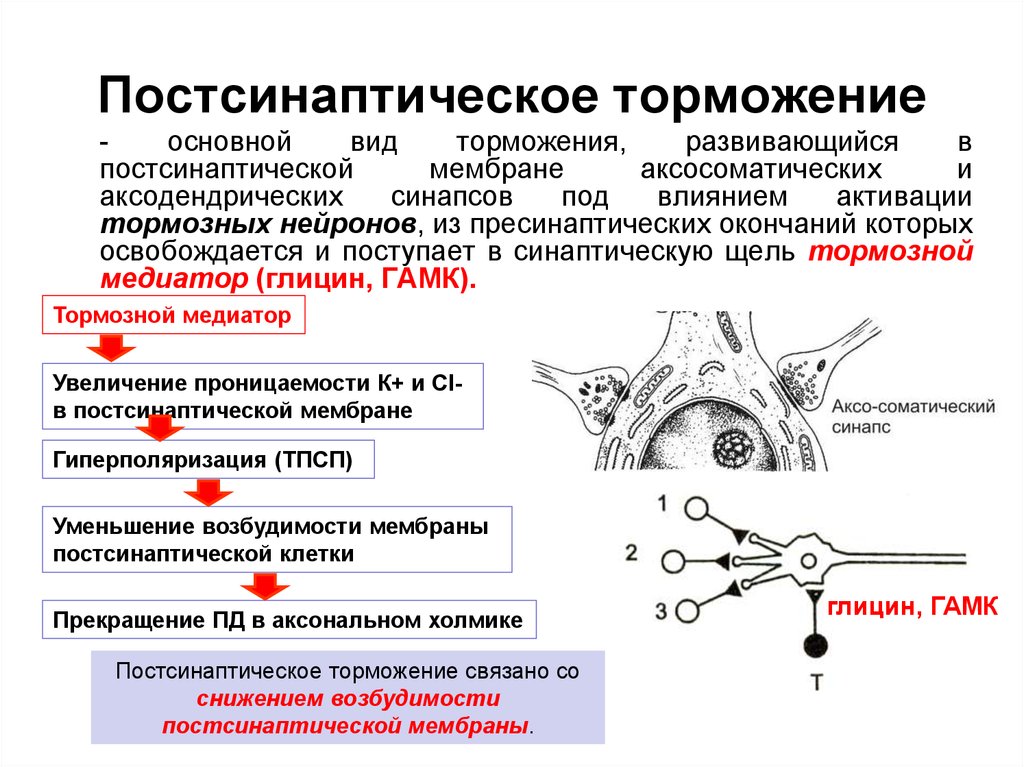 Схема постсинаптического торможения |