Молекулярная биология. Репликация днк
 Скачать 0.64 Mb. Скачать 0.64 Mb.
|

|
Молекулярная биология Репликация днкСогласно гипотезе Дж.Уотсона и Ф.Крика, каждая из цепей двойной спирали ДНК служит матрицей для репликации комплементарных дочерних цепей. При этом образуются две дочерние двухцепочечные молекулы ДНК, идентичные родительской, причем каждая из этих молекул содержит одну неизменную цепь родительской ДНК. Этот механизм репликации ДНК, названный полуконсервативным, был подтвержден в опытах на клетках Е.соli в 1957 г. М. Мезелсоном и Ф. Сталем. Исключены консервативный способ репликации, при котором одна дочерняя ДНК должна содержать обе исходные цепи, а вторая состоять из двух новосинтезированных цепей, и дисперсивный механизм репликации, при котором каждая дочерняя цепь ДНК состоит из участков родительской и новообразованной ДНК (рис.29.1). 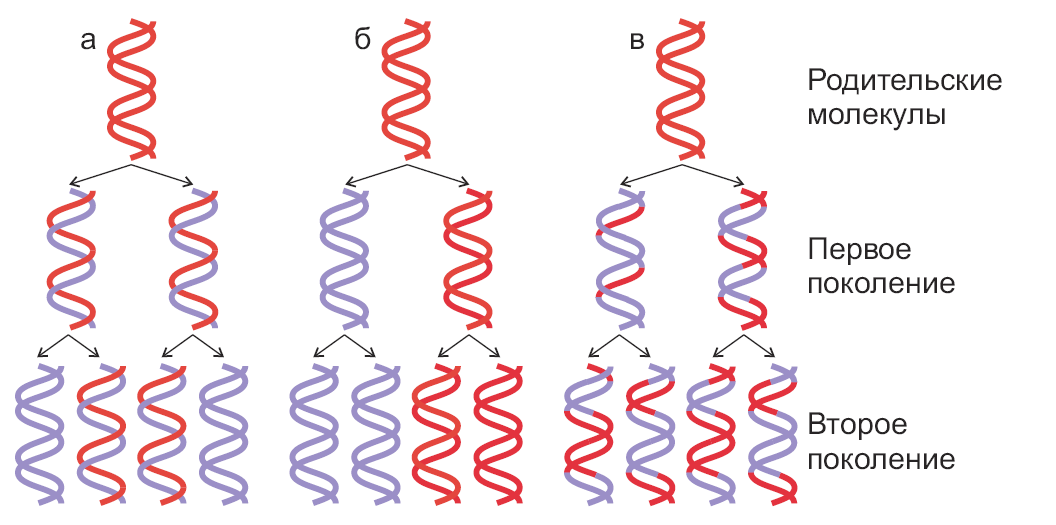 Рис.29.1. Три механизма репликации ДНК: а) полуконсервативный; б) консервативный; в) дисперсивный Для биосинтеза ДНК необходимы: 1) неспаренная цепь ДНК, которая служит матрицей, и цепь-затравка, к которой присоединяются новые нуклеотиды; 2) полный набор дезоксирибонуклеозидтрифосфатов (dNTP). При отсутствии хотя бы одного из них синтез ДНК не происходит. Цепь удлиняется от затравки, имеющей свободную 3´-ОН, в направлении 5´→3´ (путем присоединения следующего нуклеотида в соответствии с информацией, заложенной в матрице). Источником энергии в реакциях полимеризации мононуклеотидов является энергия, освобождаемая всеми четырьмя типами дезоксирибонуклеозидтрифосфатов, участвующих в синтезе ДНК. Расщепление пирофосфата до неорганического фосфата при участии неорганической пирофосфатазы сдвигает реакцию в сторону удлинения цепи; 3) ферменты и белки, участвующие в синтезе ДНК: ДНК-полимеразы, топоизомеразы (гиразы), хеликазы и лигазы, праймаза, ssb-белки. Весь комплекс, состоящий более чем из 20 репликативных ферментов и факторов, называется ДНК-репликазной системой, или реплисомой. ДНК-зависимые ДНК-полимеразы – ключевые ферменты репликативного процесса, использующие принцип комплементарности для наращивания полинуклеотидных цепей. У прокариот есть триДНК-полимеразы: Pol I, Pol II и Pol III. В репликации ДНК участвуют Pol I и Pol III. ДНК-полимераза I обладает полимеразной и (3ʹ→5ʹ , 5ʹ→3ʹ)-экзонуклеазной активностью, участвует в удалении праймера, застройке бреши, образовавшейся на месте праймера, коррекции ошибок при репликации, а также в репарации ДНК. В клетках E.coli насчитывается около 400молекул этого фермента. Pol III осуществляет репаративный синтез ДНК. Основным ферментом, катализирующим биосинтез новообразованной ДНК у прокариот, является ДНК-полимераза III (Pol III).Она обладает полимеразной и 3ʹ→5ʹ- экзонуклеазной активностью; синтезирует лидирующую и отстающую цепь ДНК, обладает корректорской функцией. В клетке содержится 10-20 молекул Pol III, она обладает повышенным сродством к матрице и обеспечивает высокую эффективность копирования. Структура ДНК-полимеразы Ш приведена на рис.29.2. 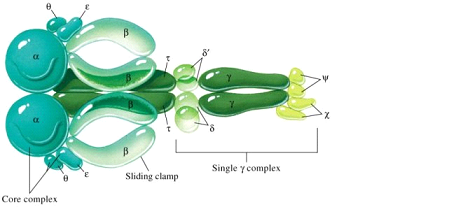 Рис. 29.2. Субъединичная структура ДНК-полимеразы III Кор-фермент состоит из субъединиц (αθε), β-белок выполняет функцию «скользящего зажима», τ-белок участвует в сборке и димеризации холофермента ДНК-полимеразы. γ-комплекс (γ,δ,δ´,χ,ψ) – ДНК-зависимая АТРаза, необходим для связывание затравки с матрицей и активации ДНК-полимеразы. ДНК-полимеразы – это полидезоксирибонуклеотид-синтетазы, ферменты класса трансфераз, катализирующие образование фосфодиэфирной связи при синтезе ДНК: R-цепь ДНК, В, Вʹ- азотистые основания: 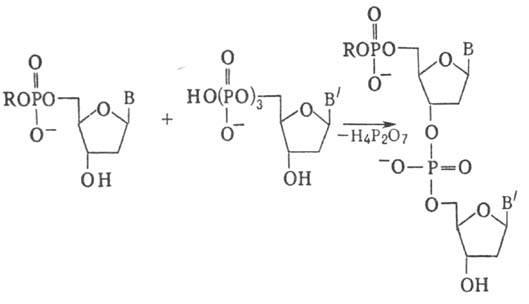 Имеются доказательства того, что в димерной форме ДНК-полимераза III катализирует сопряженный синтез ведущей (лидирующей) и отстающей цепей ДНК при репликации. ДНК-полимеразы нуждаются в затравке (праймере), поскольку они могут присоединять дезоксирибонуклеотиды только к 3ʹ-ОН-группе. Топоизомеразы участвуют в процессе раскручивания двойной спирали в репликативной вилке. Эти ферменты изменяют степень сверхспирализации и приводят к образованию «шарнира», который создает условия для непрерывного движения репликативной вилки. Идентифицированы два типа топоизомераз: топоизомеразы I типа надрезают одну из двух цепей ДНК, в результате чего концевой участок двойной спирали может повернуться вокруг интактной цепи, и затем воссоединяют концы разрезанной цепи. Топоизомеразы типа II вносят временные разрывы в обе комплементарные цепи, изменяют степень сверхспирализации, а затем соединяют разорванные концы. Топоизомеразы помогают хеликазе раскручивать ДНК для ее репликации. Хеликазы (от лат. helix - спираль, белок dnaB), осуществляют образование и продвижение вдоль спирали ДНК репликативной вилки – участка молекулы с расплетенными цепями. Эти ферменты для расплетения цепей используют энергию, высвобождающуюся при гидролизе АТР. Хеликазы действуют в комплексе с ssb-белками, которые связываются с одноцепочечными участками молекулы и тем самым стабилизируют расплетенный дуплекс. Праймазы. Репликация ДНК требует РНК-праймеров. РНК-праймеры синтезируются праймазой (рис. 29.3), кодируемой dnaG геном. Из рис 29.3 видно, что праймаза состоит из трех доменов: ■ – N-терминальный домен (110 аминокислот), содержит ДНК-связывающий мотив -цинковый палец; ■ – коровый (центральный) домен (322 аминокислоты) содержит каталитический центр; ■ – С-терминальный домен (151 аминокислота), взаимодействующий с dnaB. 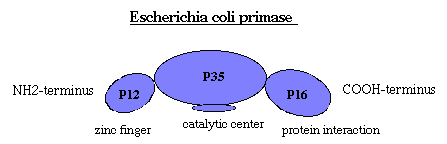 Рис. 29.3. Праймаза E.coli Праймеры, синтезируемые праймазой E.coli, начинаются с последовательности pppAG на 5´-конце и состоят примерно из 10-12 нуклеотидов. Праймазы различаются как по структуре, так и по специфичности действия. ДНК-лигазы катализируют процессы воссоединения фрагментов цепей ДНК, участвуя в образовании ковалентных связей между 5ʹ-Р- и 3ʹ-ОН-группами соседних дезоксирибонуклеотидов. Эти ферменты также используют энергию макроэргических связей, образующуюся при гидролизе АТР. В таблице 29.1 сведены основные функции ферментов и белков, участвующих в процессе репликации. Функции белков и ферментов, участвующих в репликации
Репликация ДНК идет в три стадии: инициация, элонгация и терминация. У бактерий инициация репликации ДНК начинается в уникальном сайте хромосомы, точке репликации – oriC, из которой репликация осуществляется двунаправлено до точки окончания (terminus). В результате образуются две репликативные вилки, которые продвигаются в противоположных направлениях, т. е. обе цепи реплицируются одновременно. Инициаторный белок dnaA связывается с повторяющимися сайтами связывания на oriC, образуя специализированную нуклеопротеиновую структуру. Это приводит к локальному расхождению АТ-богатой последовательности oriC, которая служит зоной связывания для репликативной хеликазы (dnaB), и белка dnaC(рис.29.4). 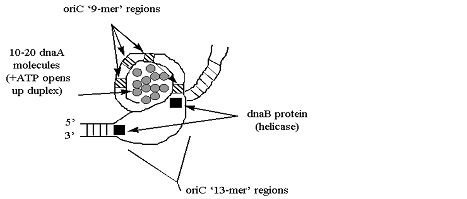 Рис.29.4. Инициация репликации ДНК Далее dnaB активируется удалением dnaC, движется на определенное расстояние в направлении 5ʹ→3ʹ и взаимодействует с праймазой dnaG. Праймаза синтезирует короткие РНК-праймеры для холофермента ДНК-полимеразы Ш. В месте инициации образуется промежуточный комплекс, состоящий по меньшей мере из пяти белков. Один из них – белок dnaB – может передвигаться вдоль ДНК, используя энергию гидролиза АТР, а также служит сигналом для активации праймазы (рис.29.5). 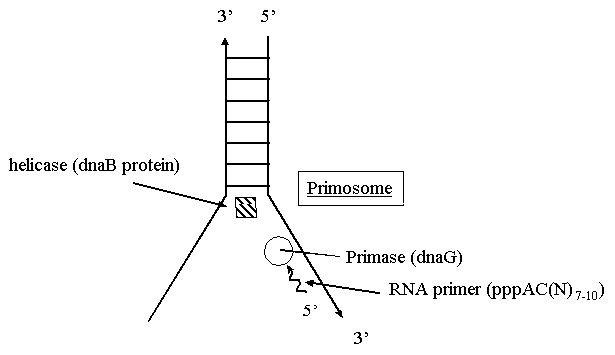 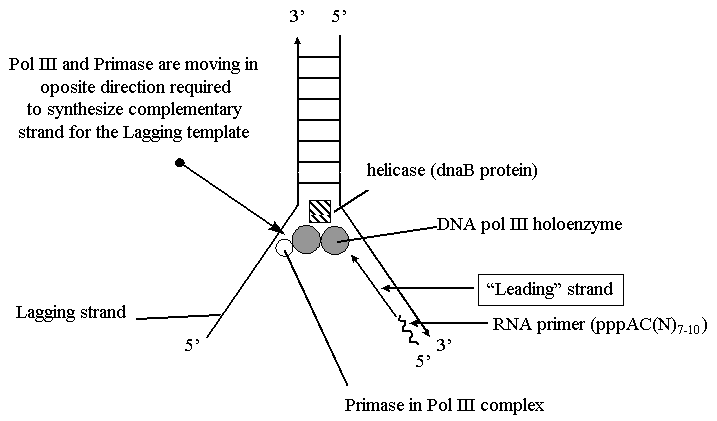 а) б) Рис. 29.5. Образование репликативной вилки и праймера на лидирующей цепи ДНК (а); переход Pol III вместе с праймазой на отстающую цепь ДНК Праймаза является компонентом праймосомы, состоящей из нескольких различных субъединиц. В состав праймосомы входит также комплекс белков DnaВ и DnaС, который вблизи репликационной вилки периодически участвует в формировании специфической вторичной структуры ДНК, подходящей для узнавания праймазой. Инициация репликации ДНК заканчивается образованием репликативной вилки и синтезом РНК-затравки на лидирующей цепи ДНК (рис.29.5) благодаря формированию репликационного комплекса (рис.29.6). 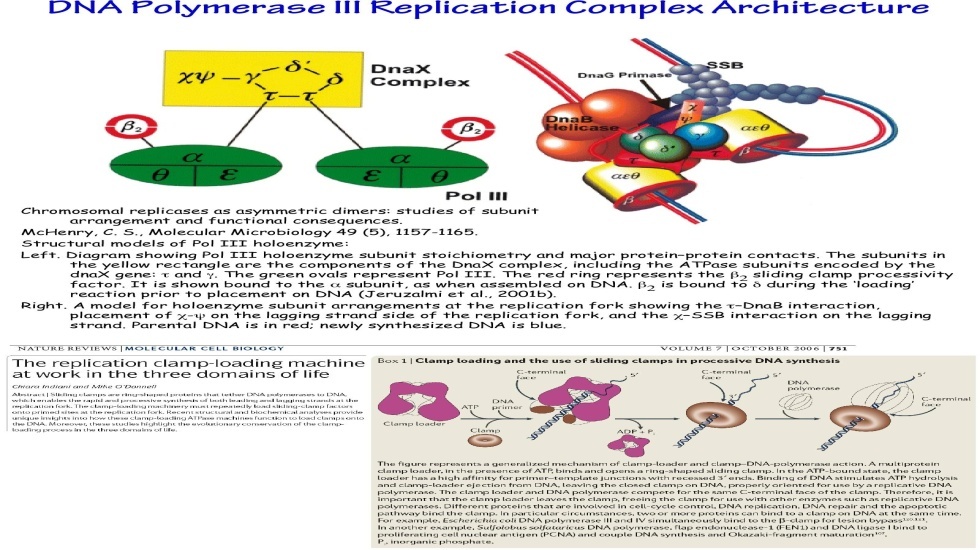 Рис.29.6. Репликационный комплекс E.coli В процессе элонгации происходит наращивание дочерних полинуклеотидных цепей ДНК. Каждая репликативная вилка включает, по крайней мере, две молекулы ДНК-полимеразы III, ассоциированные с несколькими вспомогательными белками. К последним относятся ДНК-топоизомеразы (гиразы), которые раскручивают плотно свернутую двойную спираль ДНК, и хеликазы,которые расплетают двухтяжевую ДНК на две цепи. Ведущая цепь ДНК реплицируется непрерывно в направлении, совпадающем с движением репликативной вилки. Отстающая цепь считывается в направлении, противоположном движению репликативной вилки. Преодоление антипараллельности цепей ДНК при репликации, возможно, достигается путем образования петельной структуры (рис.29.7). Вначале на отстающей цепи синтезируются короткие фрагменты новой цепи ДНК, так называемые фрагменты Оказаки, названные так по имени их первооткрывателя. Каждый фрагмент начинается с короткой РНК-затравки (праймера), необходимой для функционирования ДНК-полимеразы. ДНК-полимераза IIIдостраивает этот праймер до фрагмента ДНК длиной 1000-2000 дезоксинуклеотидных звеньев. 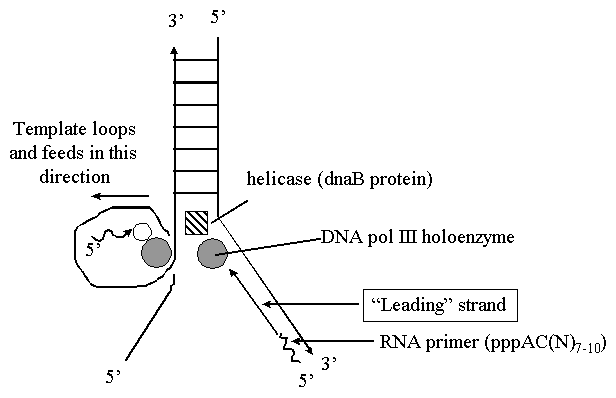 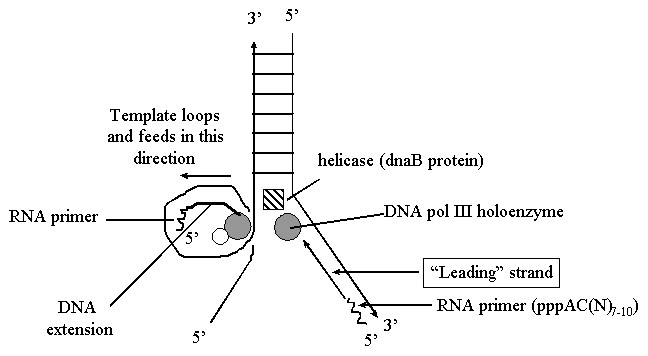 Рис.29.7. Гипотетический механизм преодоления антипараллельностицепей ДНК Кроме полимеризации цепей, которую осуществляет Рol III, в ходе элонгации ДНК происходят следующие события: 1) вырезание РНК-праймеров из лидирующей цепи и из каждого фрагмента Оказаки. Эту функцию выполняет Pol I, благодаря 5ʹ→3ʹ-экзонуклеазной активности; 2) заполнение брешей, оставшихся после удаления праймеров. В этом процессе участвует также ДНК-полимераза I, используя для встраивания нуклеотидов 3ʹ-ОН-группу соседнего фрагмента Оказаки (рис.29.8); 3) соединение фрагментов ДНК в отстающей цепи с помощью фермента ДНК-лигазы; 4) исправление ошибок репликации, благодаря 3ʹ→5ʹ-экзонуклеазной активности, которой обладают как Pol III, так и Pol I. 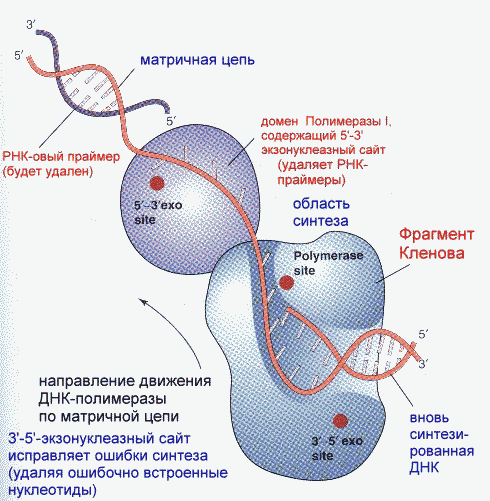 Рис.29.8. Механизм действия ДНК-полимеразы I В целом, последовательность событий, происходящих при репликации ДНК, можно представить в виде следующей схемы (рис.29.9). 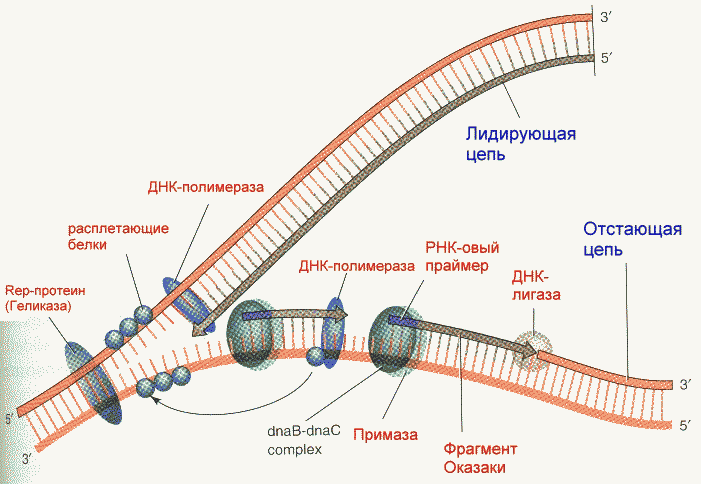 Рис.29.9. Основные участники и события репликации ДНК Терминация синтеза ДНК наступает вследствие исчерпания матрицы. Репликационные «глазки» сливаются, и на каждой матрице образуется дочерняя цепь ДНК. Точность репликацииТочность в процессе репликации очень важна, т.к. ошибки могут привести к мутациям. ДНК-полимераза III обладает способностью к «редактированию» – исправлению ошибок. По мере продвижения вдоль матричной ДНК-цепи фермент проверяет, нет ли ошибок во вновь синтезированной копии. О наличии ошибок ДНК-полимераза «узнает» по искажению двойной спирали ДНК, которое происходит в том случае, если Т соединится с G или С с А. Обнаружив такой участок, ДНК-полимеразный ферментный комплекс возвращается на один шаг назад и благодаря своей экзонуклеазной активности, вырезает неправильное основание (или их группу) и вставляет то, которое должно быть на этом месте (правильное основание). Скорость точной репликации у бактерий примерно 500 оснований в секунду, а у высших клеток, включая клетки человека, около 50 оснований в секунду. Репликация днк у эукариотДНК хромосом высших клеток намного длиннее, а сами хромосомы устроены намного сложнее, чем маленькие и простые бактериальные геномы. У высших клеток, в отличие от бактерий, ДНК в хромосомах образует комплекс с белками (гистонами), которые участвуют в сворачивании длинных нитей ДНК в серию петель для того, чтобы их можно было упаковать внутри ядра. Репликация ДНК начинается одновременно в нескольких сайтах каждой хромосомы. У эукариот число их может превысить 1000, поэтому большой набор ДНК-последовательностей реплицируется за 5-20 часов. Из каждой такой точки в противоположных направлениях одновременно движутся две репликативные вилки. Репликация продолжается до полного завершения синтеза дочерних цепей и разделения новых дуплексов. У млекопитающих и человека известно пять разных ДНК-полимераз. Репликативный синтез ДНК катализируют две ДНК-полимеразы (a и δ); β- и ε-полимеразы участвуют в репарации ДНК, γ-полимераза – в репликации митохондриальной ДНК. Поскольку эукариотическая ДНК линейна, при репликации из-за вырезания праймеров на концах полинуклеотидных цепей образуются недореплицированные участки. Их репликация осуществляется с помощью особого фермента – теломеразы (рис.29.10). 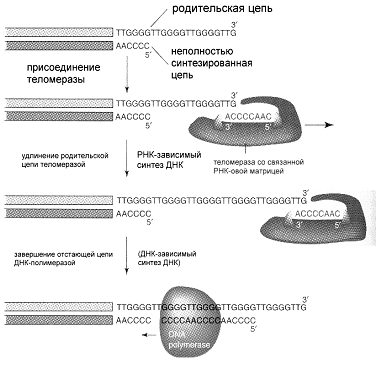 Рис. 29.10. Репликация теломерных участков хромосом Теломераза, содержащая в себе последовательность нуклеотидов, за несколько приемов удлиняет укороченную цепь, создавая пространство для работы праймазы и ДНК-полимеразы, после чего избытки нуклеотидов удаляются: Репаративный синтез днкВоздействие на организм неблагоприятных факторов (химические соединения, ультрафиолетовое излучение и др.) приводит к постоянному накоплению ошибок в геноме, которые в конечном итоге вызывают появление патологии, в частности, – невыясненный до сих пор механизм раковых заболеваний. Пока лишь существуют только предположения о том, что причиной раковых заболеваний являются дефекты в носителях информации – ДНК. Возможны, как минимум, четыре типа повреждений ДНК: 1) повреждение одиночных оснований (жезаминирование цитозина в урацил, аденина в гипоксантин; алкилирование оснований; включение аналогов оснований, инсерции и делеции нуклеотидов и др.); 2) повреждение пары оснований, например, индуцированное ультрафиолетовым излучением образование тиминовых димеров; 3) разрывы цепей при действии ионизирующей радиации; 4) образование перекрестных связей между основаниями, а также между ДНК и белками, например, гистонами. На рис. 29.11 показано образование пиримидиновых димеров при воздействии ультрафиолетовых лучей.. 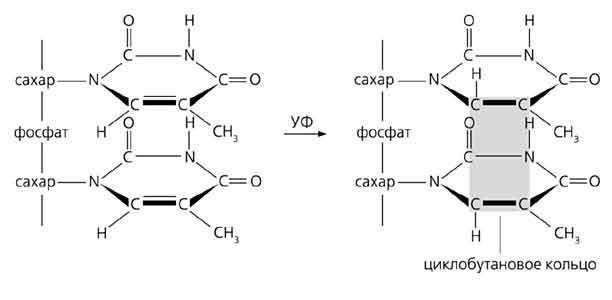 Рис.29.11. Образование тиминовых димеров в ДНК при ультрафиолетовом облучении ДНК с таким нарушением уже не может нормально работать. С нее теперь невозможно считать информацию, необходимую для производства белков, или снять копию. Следовательно, процессы жизнедеятельности клетки нарушаются, а деление ее останавливается. Ферменты, ответственные за копирование ДНК и считывание с нее информации, дойдя до тиминового димера, либо «перепрыгнут» через него, что приведет к разрыву синтезируемого белка на две части, либо вовсе остановятся. В случае такого повреждения репарация начинается с того, что специальный белок, УФ-эндонуклеаза, находит тиминовый димер и рядом с ним разрывает цепь ДНК (рис. 29.12). 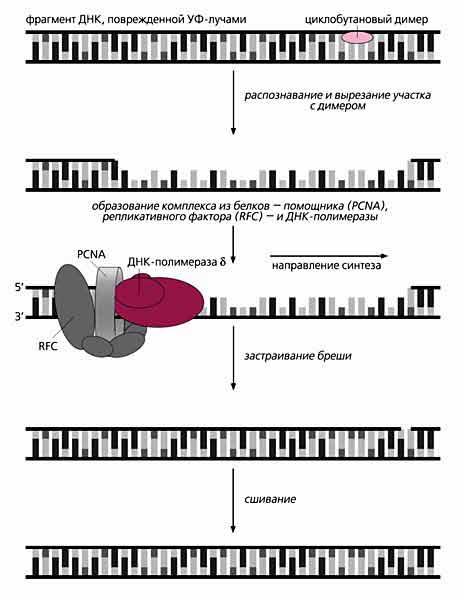 Рис.29.12. Репарация тиминовых димеров При этом вторая цепь остается целой. Затем в работу включается другой белок – экзонуклеаза. Она, отщепляя по одному, удаляет по обе стороны от разрыва несколько сотен нуклеотидов. В результате на цепи ДНК (там, где был обнаружен тиминовый димер) возникает брешь длиной в несколько тысяч нуклеотидов. Эту брешь теперь быстро заделывает третий белок – ДНК-полимераза I (у прокариот) или ДНК-полимераза δ (у эукариот). По принципу комплементарности и на основе информации о второй, нетронутой цепочке ДНК, полимераза встраивает нуклеотиды в первую цепочку. Завершающий этап осуществляет ДНК-лигаза. Она устраняет разрывы на репарированной цепочке ДНК. В результате структура ДНК полностью восстанавливается. |