Справочный материал. Глава 06 – Физиология синапсов. Рис. 6 Межнейронный (аксосоматический) синапс
 Скачать 298 Kb. Скачать 298 Kb.
|

|
Справочный материал по Физиологии. Глава 6 – Физиология синапсов. Передачу возбуждения между нейронами, а также от нейронов к мышечным и секреторным клеткам осуществляют специализированные межклеточные контакты — синапсы (рис. 6–1, 6–2, 6–6, см. также рис. 5–1). 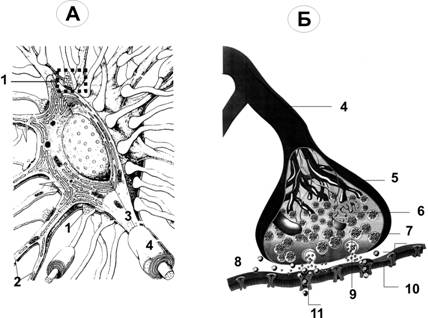 Рис. 6–1. Межнейронный (аксосоматический) синапс. А. На поверхности перикариона заканчивается множество (например, на мотонейронах до 10 000) концевых разветвлений (пресинаптических терминалей) аксонов от других нейронов; это аксосоматические синапсы. Один из них (1) представлен в правой части схемы. 2 — ветвления одного из дендритов. 3 — место отхождения аксона (аксонный холмик). 4 — миелинизированный аксон. Б. Между концевой терминалью аксона (5) пресинаптического нейрона и поверхностью перикариона постсинаптического нейрона (10) находится синаптическая щель (8) с диффундирующими в ней молекулами нейромедиатора (9). Для терминали аксона (пресинаптическая часть синапса, 5) характерны митохондрии (6) и содержащие молекулы нейромедиатора синаптические пузырьки (7). В постсинаптическую мембрану вмонтированы рецепторы для связывания нейромедиатора и многочисленные ионные каналы (11). Помимо аксосоматических, между нейронами образуются аксоаксональные, аксодендритические и дендродендритические синапсы. Большинство межнейронных синапсов относится к аксодендритическим (например, в коре больших полушарий — до 98%). Пресинаптическая клетка выделяет в межклеточное пространство между контактирующими клетками (синаптическая щель) химический посредник — нейромедиатор. Молекулы нейромедиаторов взаимодействуют с их рецепторами на постсинаптической клетке, что приводит к изменениям мембранного потенциала (МП): деполяризации (возбуждающие синапсы) или гиперполяризации (тормозные синапсы). Синаптическая передача обладает пластичностью, т.е. способностью к облегчению, потенциации и депрессии. В синапсах проведение возбуждения всегда происходит в одном направлении — от пресинаптической терминали к постсинаптической клетке. 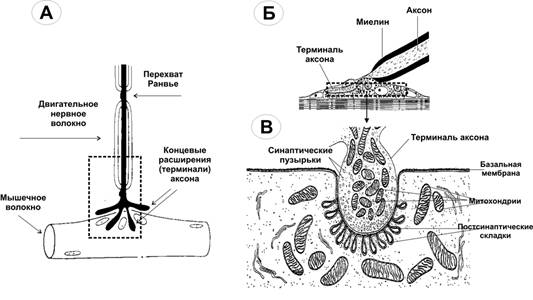 Рис. 6–2. Нервно-мышечный синапс (А и Б — фрагмент выделенной прямоугольником области на А) образуется между концевыми расширениями (терминалями) соматических мотонейронов (пресинаптическая часть синапса) и поверхностью скелетного МВ (постсинаптическая часть). В — фрагмент обозначенной прямоугольником на Б. На пре- и постсинаптической стороне видны многочисленные митохондрии, необходимые для поддержания мембранного электрогенеза, в терминали расположено множество содержащих нейромедиатор (ацетилхолин) синаптических пузырьков, освобождающих ацетилхолин в синаптическую щель и взаимодействующих с никотиновыми холинорецепторами, вмонтированными в постсинаптическую мембрану. [11]. Термин «синапс» (от греч. synapsis — соприкосновение, соединение) предложил в 1897 г. Чарлз Шеррингтон. Отто Лёви, Генри Дейл и А.В. Кибяков доказали, что передача возбуждения в синапсах имеет химическую природу. Помимо химической передачи в синапсах, между нейронами, а также нейронами и глиоцитами возможна и электрическая передача возбуждения — при помощи щелевых контактов, обеспечивающих электротоническое, метаболическое и информационное сообщение между контактирующими клетками. По отношению к таким межклеточным контактам между нервными элементами применяют термин «эфапс» (греч. ephapsis — прикосновение, касание, контакт). Такие электрические синапсы широко распространены в нервной системе беспозвоночных, но изредка встречаются и в ЦНС млекопитающих. Так, в сетчатке глаза горизонтальные клетки связаны между собой щелевыми контактами. Благодаря этому локальные потенциалы, потенциалы действия, а также продукты метаболизма и красители (в эксперименте) могут распространяться от клетки к клетке, оказывая значительное влияние на процесс переработки зрительной информации. Основные отличия между химическими и электрическими синапсами представлены в табл. 6–1. Таблица 6-1. Химические и электрические контакты между нейронами
Организация и функция синапса В синапсе различают пресинаптическую часть, постсинаптическую часть и расположенную между клетками синаптическую щель (см. рис 6–1Б и 6–6, а также рис. 6–5 в книге). Пресинаптическая часть Пресинаптическая часть содержит синаптические пузырьки с нейромедиатором, элементы цитоскелета и митохондрии. В пресинаптическую мембрану встроены потенциалозависимые Ca2+ каналы. При поступлении ПД к терминальному расширению мембрана деполяризуется, Ca2+ каналы открываются, ионы Ca2+ входят в терминаль, запуская в активных зонах процесс слияния мембраны синаптического пузырька и пресинаптической мембраны, т.е. секрецию (экзоцитоз) нейромедиатора (рис. 6–6, позиции 2–4). Роль Са2+. Слияние синаптических пузырьков с пресинаптической мембраной происходит при увеличении концентрации Са2+ в цитозоле нервной терминали. Белок синаптического пузырька синаптотагмин связывается с Са2+ и тем самым принимает участие в регуляции экзоцитоза (в том числе путём реорганизации примембранного цитоскелета). Синаптические пузырьки. Молекулы нейромедиатора накапливаются в нервной терминали, находясь внутри синаптических пузырьков вместе с АТФ и некоторыми катионами. В каждом пузырьке находится несколько тысяч молекул нейромедиатора, что составляет квант нейромедиатора. Синтез нейромедиатора. Ферменты, необходимые для образования нейромедиаторов, синтезируются в перикарионе и транспортируются к синаптической терминали по аксонам (рис. 6–4). Типы пузырьков — мелкие (диаметр порядка 50 нм) и крупные (диаметр 100–200 нм). Мелкие синаптические пузырьки содержат «классические» медиаторы (см. ниже). Крупные везикулы содержат нейропептиды. Секреция. Когда ПД достигает нервной терминали, синаптические пузырьки сливаются с пресинаптической мембраной, что приводит к выделению квантов нейромедиатора в синаптическую щель. Незначительное количество квантов нейромедиатора постоянно (спонтанно) секретируется в синаптическую щель. Узнавание. Предшествующий слиянию синаптических пузырьков и плазмолеммы процесс узнавания синаптическим пузырьком пресинаптической мембраны происходит при взаимодействии мембранных белков (синаптобревин, SNAP-25, синтаксин и другие). Влияние токсинов. Синтаксин, SNAP-25 и синаптобревин — мишени ботулинического токсина, необратимо подавляющего слияние синаптических пузырьков с пресинаптической мембраной. Мишень столбнячного токсина — синаптобревин. Активные зоны (рис. 6–3). Секреция нейромедиатора осуществляется в специализированных участках пресинаптического нервного окончания — активных зонах — участках утолщения пресинаптической мембраны. Активная зона состоит из «плотной полоски» на пресинаптической мембране и сгруппированных около неё синаптических пузырьков, потенциалозависимых кальциевых каналов, специальных белков экзоцитоза и элементов цитоскелета. Количество активных зон в нервно-мышечном синапсе достигает 30–40, в межнейронных синапсах — около десятка. Активные зоны расположены против скоплений рецепторов в постсинаптической мембране, что уменьшает задержку в передаче сигнала, связанную с диффузией нейромедиатора в синаптической щели. 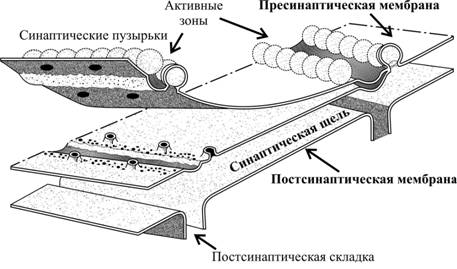 Рис. 6–3. Активные зоны нервно-мышечного синапса расположены напротив постсинаптических складок — участков скоплений холинорецепторов. Пресинаптическая мембрана слева расщеплена на два листка. Жизненный цикл синаптических пузырьков (рис. 6–4). Синаптические везикулы образуются в теле нейрона в эндоплазматическом ретикулуме и комплексе Гольджи (1) и с аксонным транспортом поступают в нервные окончания (2). В нервном окончании мелкие синаптические пузырьки посредством активного транспорта заполняются медиатором (3) и передвигаются к пресинаптической мембране (4). Освобождение медиатора (5) может осуществляться посредством экзоцитоза с полным («классический» механизм) либо неполным (механизм «kiss and run») слиянием. Первый вид экзоцитоза сопровождается встраиванием мембраны везикулы в пресинаптическую, опорожнением пузырька, а затем посредством эндоцитоза образуются покрытые клатрином везикулы (6), которые затем проходят стадию эндосомы (7) и снова заполняются медиатором (3). Второй вид экзоцитоза характеризуется образованием временной поры, соединяющей полость пузырька с синаптической щелью. После выделения медиатора везикула не встраивается в пресинаптическую мембрану, а отпочковывается от неё (8) и повторно заполняется медиатором (3). Крупные синаптические везикулы заполняются медиатором в теле клетки (9), их экзоцитоз происходит в других участках пресинаптической мембраны, а эндоцитоз опорожнённых пузырьков отсутствует (10). 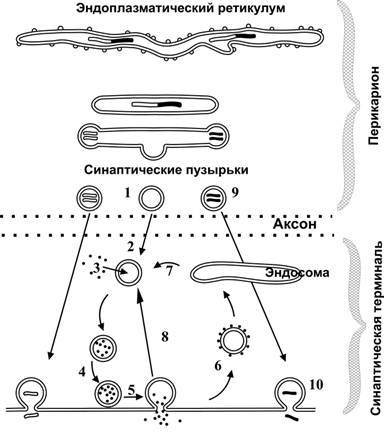 Рис. 6–4. Образование, транспорт и экзоцитоз синаптических пузырьков. Синаптическая щель — промежуток между пре- и постсинаптическими мембранами шириной 20–35 нм. В синаптическую щель из синаптических пузырьков выделяются молекулы нейромедиатора и путём диффузии достигают постсинаптической мембраны. В синаптической щели находятся ферменты, расщепляющие молекулы нейромедиатора (например, ацетилхолинэстераза, гидролизующая ацетилхолин), а в пресинаптическую мембрану вмонтированы переносчики, осуществляющие перенос нейромедиаторов–аминокислот и биогенных аминов (например, глутамата, аспартата, норадреналина) в пресинаптическую терминаль. Таким образом, удаление нейромедиатора из синаптической щели происходит двояко: инактивация ферментом или захват пресинаптической терминалью. Инактивация нейромедиатора. Кратковременность взаимодействия нейромедиатора с рецептором достигается разрушением нейромедиатора специальными ферментами (например, ацетилхолина — ацетилхолинэстеразой). Захват нейромедиатора. В большинстве синапсов передача сигналов прекращается вследствие быстрого захвата нейромедиатора пресинаптической терминалью. Транспортёры. Захват норадреналина осуществляют специфические Na+- и Cl–-транспортирующие белки (например, норадреналин–транспортирующий белок 1) — мишени трициклических антидепрессантов (например, дезипрамин и имипрамин). Система захвата биогенных аминов — точка приложения антидепрессантов и таких препаратов, как кокаин и амфетамины. Дефекты транспортёров норадреналина и серотонина — кандидаты на роль первопричины при психиатрических расстройствах, таких как маниакально-депрессивные состояния. Постсинаптическая часть В постсинаптической мембране находятся рецепторы, чувствительные к нейромедиатору. Взаимодействие нейромедиатора с рецептором приводит к изменению МП постсинаптической мембраны. В зависимости от характера возникающего постсинаптического потенциала (деполяризация или гиперполяризация) различают синапсы возбуждающие и тормозные. Возбуждающие синапсы. Материал этого раздела см. в книге. Тормозные синапсы. При гиперполяризации возбудимость мембраны уменьшается, и ПД не генерируются. Характер электрического ответа постсинаптической стороны и дальнейший физиологический эффект определяются свойствами рецепторов. С точки зрения механизма открытия ионных каналов и последующей де- или гиперполяризации (рис. 6–6) постсинаптические рецепторы подразделяют на ионотропные (от «ион») и метаботропные (от «метаболизм»). Ионотропные рецепторы сами по себе являются ионными каналами. Классический пример — н холинорецепторы (рис. 6–6, слева). Метаботропные рецепторы связаны с ферментами (аденилатциклаза или фосфолипаза C) через G белок. Классический пример — м холинорецепторы (рис. 6–6, справа). 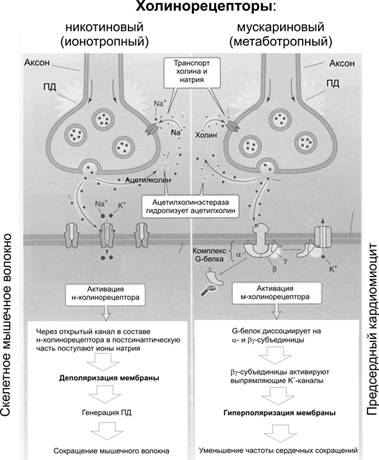 Рис. 6–6. Ионотропные и метаботропные рецепторы в холинергических синапсах [2] Постсинаптические плотности. Область, расположенная с цитоплазматической стороны постсинаптической мембраны, в электронном микроскопе выглядит как зернистая плотная полоска, содержащая множество белков. К ним относятся рецепторы нейромедиаторов, протеинкиназы, структурные и цитоскелетные белки, а также белки, осуществляющие эндоцитоз и гликолиз. Дендритные шипики — характерная особенность 90% возбуждающих синапсов в ЦНС. Эти мелкие (менее 1 мкм длиной) удлинённые структуры, выступающие над поверхностью дендритов и имеющие постсинаптические плотности. Их функция точно не установлена; возможно, они увеличивают поверхность постсинаптической стороны или являются резервуаром для Ca2+. Этапы синаптической передачи Синаптическая передача возможна при реализации ряда последовательных процессов: синтез нейромедиатора, его накопление и хранение в синаптических пузырьках вблизи пресинаптической мембраны, высвобождение нейромедиатора из нервной терминали, кратковременное взаимодействие нейромедиатора с рецептором, встроенным в постсинаптическую мембрану; разрушение нейромедиатора или захват его нервной терминалью. На рис. 6–7 представлены стандартные этапы синаптической передачи. 1. Молекулы нейромедиатора поступают в мембранные синаптические пузырьки, располагающиеся в пресинаптической терминали и концентрирующиеся в активных зонах пресинаптической мембраны. 2. Приходящий по аксону ПД деполяризует пресинаптическую мембрану. 3. Вследствие деполяризации открываются потенциалозависимые Са2+ каналы, и Са2+ поступает в терминаль. 4. Увеличение внутриклеточного [Са2+] запускает слияние синаптических пузырьков с пресинаптической мембраной и выброс нейромедиатора в синаптическую щель (экзоцитоз). 5. Кванты нейромедиатора, поступившие в синаптическую щель, диффундируют в ней. Часть молекул нейромедиатора связывается со специфичными для них рецепторами постсинаптической мембраны. 6. Связавшие нейромедиатор рецепторы активированы, что приводит к изменению поляризации постсинаптической мембраны либо прямо (поступление ионов через ионотропные рецепторы) либо опосредованно — активация ионных каналов через систему G белка (метаботропные рецепторы). 7. Инактивация нейромедиаторов происходит либо путём их ферментной деградации, либо молекулы нейромедиатора захватываются клетками. 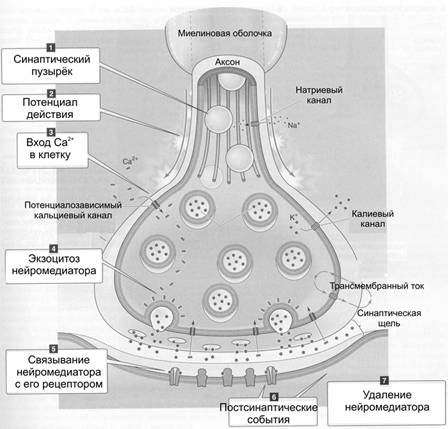 Рис. 6–7. Этапы синаптической передачи [2]. Таким образом, синаптическая передача — сложный каскад событий. Многие неврологические и психиатрические заболевания сопровождаются нарушением синаптической передачи. Различные ЛС влияют на синаптическую передачу, вызывая нежелательный эффект (например, галлюциногены) или, наоборот, корригируя патологический процесс (например, психофармакологические антипсихотические средства). Постсинаптические потенциалы Вызванные и спонтанные постсинаптические потенциалы (ПСП). Нейромедиаторы при связывании с ионотропными рецепторами приводят к возникновению ПСП. Количество квантов медиатора, содержащееся в одном ПСП, определяет его квантовый состав. При возбуждении пресинаптической терминали ПД и секреции множества синаптических пузырьков регистрируются вызванные, или многоквантовые ПСП. Существуют также спонтанные, или миниатюрные ПСП, обусловленные случайным (в отсутствии ПД) экзоцитозом медиатора в синаптическую щель. Эти сигналы обычно одноквантовые и незначительны по амплитуде. Синаптическая задержка. Между моментом поступления ПД к пресинаптическому нервному окончанию и временем возникновения ПСП существует временной интервал в 0,5–1 мс, получивший название синаптической задержки. Она объясняется прежде всего временем, необходимым для выделения медиатора и его действия на постсинаптическую мембрану. Наиболее длительным при этом является процесс запуска секреции медиатора. Возбуждающие и тормозные ПСП. ПСП могут быть либо возбуждающими (деполяризующими), либо тормозными (гиперполяризующими). Возбуждающие ПСП вызваны возрастанием проводимости мембраны для Na+. Они деполяризуют постсинаптическую мембрану, повышают возбудимость клетки, а при достижении критического уровня деполяризации приводят к возникновению ПД. Так, активация н холинорецепторов и глутаматных (ионотропных) рецепторов приводит к возникновению возбуждающих ПСП. Пора (канал) этих рецепторов имеет относительно большой диаметр, несет отрицательный заряд и проницаема для катионов (Na+, К+, Са2+), но через пору внутрь клетки в основном проходят ионы Na+ в силу гораздо большего электрохимического градиента. Тормозные ПСП вызваны повышением проводимости мембраны для K+ и Cl–. Они гиперполяризуют постсинаптическую мембрану, понижают возбудимость клетки и препятствуют генерации ПД. Этот процесс получил название постсинаптического торможения. Так, активация глициновых рецепторов и рецепторов ГАМК типа А приводит к возникновению тормозных ПСП. Эти рецепторы пропускают внутрь клетки ионы Cl–. Возникновение потенциала действия в постсинаптическом нейроне. Постоянное взаимодействие возбуждающей и тормозящей активности на постсинаптическом нейроне приводит к флуктуации МП, представляющего собой алгебраическую сумму деполяризующих и гиперполяризующих влияний. В этом отношении дендритная зона (дендриты и перикарион, см. рис. 5–1) действует как определённого вида интегратор. Участком нервной клетки с самым низким порогом для генерации ПД является аксонный холмик, содержащий в своей мембране в 7 раз больше потенциалозависимых Na+ каналов, чем дендритная зона, и соответственно может генерировать ПД с большей лёгкостью. Возникший ПД распространяется в двух направлениях: вдоль аксона и по телу нейрона. В теле нейрона и дендритах ретроградного направления ПД затухает, так как в этих отделах клетки мало потенциалозависимых Na+ каналов. В терминальном расширении аксона ПД приводит к выделению медиатора в синаптическую щель. Характеристика отдельных нейромедиаторов Ацетилхолин секретируется из терминалей соматических мотонейронов (нервно-мышечные синапсы), преганглионарных волокон, постганглионарных холинергических (парасимпатических) волокон вегетативной нервной системы и разветвлений аксонов многих нейронов ЦНС (базальные ганглии, двигательная кора). Синтезируется из холина и ацетил-КоА при помощи холинацетилтрансферазы, взаимодействует с холинорецепторами нескольких типов. Кратковременное взаимодействие лиганда с рецептором прекращает ацетилхолинэстераза, гидролизующая ацетилхолин на холин и ацетат. Ботулинический токсин Clostridium botulinum угнетает секрецию ацетилхолина. Фосфорорганические соединения (ФОС) ингибируют ацетилхолинэстеразу, что приводит к увеличению количества ацетилхолина в синаптической щели. При отравлении ФОС пралидоксим способствует отделению ФОС от фермента, атропин защищает холинорецепторы от взаимодействия с избыточным количеством нейромедиатора. Токсины бледной поганки Amanita phalloides не только ингибируют активность ацетилхолинэстеразы, но и блокируют холинорецепторы. Дофамин — нейромедиатор в окончаниях некоторых аксонов периферических нервов и многих нейронов ЦНС (чёрное вещество, средний мозг, гипоталамус). После секреции и взаимодействия с рецепторами дофамин активно захватывается пресинаптической терминалью, где его расщепляет моноаминооксидаза. Дофамин метаболизирует с образованием ряда веществ, в т.ч. гомованилиновой кислоты. Шизофрения. При этом заболевании наблюдается увеличение количества D2-рецепторов дофамина. Антипсихотические средства снижают активность дофаминергической системы до нормального уровня. Хорея наследственная — нарушение функции нейронов коры и полосатого тела — также сопровождается повышенной реактивностью дофаминергической системы. Болезнь Паркинсона — патологическое уменьшение количества нейронов в чёрном веществе и других областях мозга с уменьшением уровня дофамина и метионин–энкефалина, преобладанием эффектов холинергической системы. Применение L- ДОФА увеличивает уровень дофамина, амантадин стимулирует секрецию дофамина, бромокриптин активирует рецепторы дофамина. Антихолинергические препараты уменьшают активность холинергической системы мозга. Норадреналин секретируется из большинства постганглионарных симпатических волокон и является нейромедиатором между многими нейронами ЦНС (например, гипоталамус, голубоватое место). Образуется из дофамина путём гидролиза при помощи дофамин--гидроксилазы. Норадреналин хранится в синаптических пузырьках, после высвобождения взаимодействует с адренорецепторами, реакция прекращается в результате захвата норадреналина пресинаптической частью. Уровень норадреналина определяется активностью тирозин гидроксилазы и моноаминооксидазы. Моноаминооксидаза и катехол-О-метилтрансфераза переводят норадреналин в неактивные метаболиты (норметанефрин, 3-метокси-4-гидрокси-фенилэтиленгликоль, 3-метокси-4-гидроксиминдальная кислота). Норадреналин — мощный вазоконстриктор, эффект происходит при взаимодействии нейромедиатора с ГМК стенки кровеносных сосудов. Серотонин (5-гидрокситриптамин) — нейромедиатор многих центральных нейронов (например, ядра шва, нейроны восходящей ретикулярной активирующей системы). Предшественником служит триптофан, гидроксилируемый триптофангидроксилазой до 5-гидрокситриптофана с последующим декарбоксилированием декарбоксилазой L-аминокислот. Расщепляется моноаминооксидазой с образованием 5-гидроксииндолуксусной кислоты. Депрессия характеризуется снижением количества двух нейромедиаторов (норадреналина и серотонина) и увеличением экспрессии их рецепторов. Антидепрессанты уменьшают число этих рецепторов. Маниакальный синдром. При этом состоянии увеличивается уровень норадреналина на фоне снижения количества серотонина и адренорецепторов. Литий снижает секрецию норадреналина, образование вторых посредников и увеличивает экспрессию адренорецепторов. -Аминомасляная кислота — тормозный нейромедиатор в ЦНС (базальные ганглии, мозжечок). Образуется из глутаминовой кислоты под действием декарбоксилазы глутаминовой кислоты, захватывается из межклеточного пространства пресинаптической частью и деградирует под влиянием трансаминазы ГАМК. Эпилепсия — внезапные синхронные вспышки активности групп нейронов в разных областях мозга, связывают со снижением тормозного действия -аминомасляной кислоты. Фенитоин стабилизирует плазмолемму нейронов и снижает избыточную секрецию нейромедиатора, фенобарбитал повышает связывание ГАМК с рецепторами, вальпроевая кислота увеличивает содержание нейромедиатора. Состояние тревоги — психотическая реакция, связанная с уменьшением тормозного эффекта ГАМК. Бензодиазепины стимулируют взаимодействие нейромедиатора с рецептором и поддерживают ингибиторное действие -аминомасляной кислоты. -Эндорфин — нейромедиатор полипептидной природы многих нейронов ЦНС (гипоталамус, миндалина мозжечка, таламус, голубоватое место). Проопиомеланокортин транспортируется по аксонам и расщепляется пептидазами на фрагменты, одним из которых является -эндорфин. Нейромедиатор секретируется в синапсе, взаимодействует с рецепторами на постсинаптической мембране, а затем гидролизуется пептидазами. Метионин–энкефалин и лейцин–энкефалин — небольшие пептиды (5 аминокислотных остатков), присутствующие во многих нейронах ЦНС (бледный шар, таламус, хвостатое ядро, центральное серое вещество). Как и эндорфин, образуются из проопиомеланокортина. После секреции взаимодействуют с пептидергическими (опиоидными) рецепторами. Динорфины. Эта группа нейромедиаторов состоит из 7 пептидов близкой аминокислотной последовательности, которые присутствуют в нейронах тех же анатомических областей, что и энкефалинергические нейроны. Образуются из продинорфина, инактивируются путём гидролиза. Вещество Р — нейромедиатор пептидной природы в нейронах центральной и периферической нервной системы (базальные ганглии, гипоталамус, спинномозговые узлы). Передача болевых стимулов реализуется при помощи вещества Р и опиоидных пептидов. Глицин, глутаминовая и аспарагиновая кислоты. Эти аминокислоты в некоторых синапсах являются нейромедиаторами (глицин во вставочных нейронах спинного мозга, глутаминовая кислота — в нейронах мозжечка и спинного мозга, аспарагиновая кислота — в нейронах коры). Глутаминовая и аспарагиновая кислоты вызывают возбуждающие ответы, а глицин — тормозные. синапсы в нейронных сетях Выше была рассмотрена физиология единичного синапса — важного элемента взаимодействия между нейронами, но в условиях целостного организма основная задача нервной системы — передача и переработка информации — не может быть сведена к работе отдельных синапсов. Напротив, функции нервной системы выполняются только при условии взаимодействия посредством синапсов множества нервных клеток — нейронных цепочек и сетей. При этом в нейронных сетях проявляются такие важные свойства как торможение, утомление, суммация, окклюзия, облегчение, депрессия и потенциация. Пластичность синапсов В ходе функционирования синапсы подвергаются функциональным и морфологическим перестройкам. Этот процесс назван синаптической пластичностью. Наиболее ярко такие изменения проявляются при высокочастотной, или тетанической активности, являющейся естественным условием функционирования синапсов in vivo. Например, частота импульсации вставочных нейронов в ЦНС достигает 1000 Гц. Пластичность (рис. 6–10) может проявляться либо в увеличении (облегчении, потенциации), либо уменьшении (депрессии) эффективности синаптической передачи. Выделяют кратковременные (длятся секунды и минуты) и долговременные (длятся часы, месяцы, годы) формы синаптической пластичности. Последние интересны тем, что они имеют отношение к процессам научения и памяти. 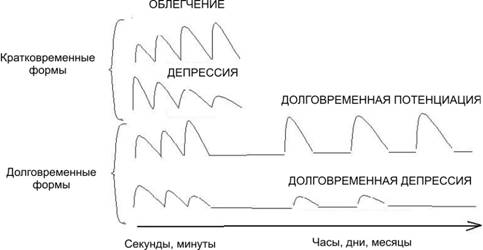 Рис. 6–10. Формы синаптической пластичности Кратковременные формы синаптической пластичности. К ним относятся облегчение, потенциация, депрессия и привыкание. Облегчение. В процессе активности в синапсах с исходно низким уровнем секреции нередко происходит увеличение амплитуды постсинаптического потенциала (ПСП). Этот процесс — облегчение — имеет пресинаптическую природу и объясняется теорией «остаточного кальция». Согласно этой теории, в процессе высокочастотной активности в пресинаптической терминали наблюдается повышение концентрации Са2+, вследствие чего происходит увеличение вероятности освобождения квантов нейромедиатора. Потенциация, посттетаническая потенциация (сенситизация). Увеличение ПСП при высокочастотной активности может иметь и постсинаптическую природу. Такой вид пластичности связан с повышением чувствительности постсинаптических рецепторов к нейромедиатору и называется потенциацией. Величина ПСП может некоторое время (секунды и минуты) оставаться повышенной и после окончания тетанической активности. Это посттетаническая потенциация (в ЦНС — сенситизация). Депрессия и привыкание (габитуация). В синапсах с исходно высоким уровнем секреции высокочастотная активность может приводить к уменьшению величины ПСП. Этот процесс — депрессия — связан преимущественно с истощением запаса нейромедиатора в пресинаптическом нервном окончании. Депрессия является одним из механизмов привыкания (габитуации). Долговременные формы синаптической пластичности Долговременная потенциация — быстро развивающееся устойчивое усиление синаптической передачи в ответ на высокочастотное раздражение. Этот вид пластичности может продолжаться дни и месяцы (рис. 6–11). Долговременная потенциация наблюдается во всех отделах ЦНС, но наиболее полно изучена на глутаматергических синапсах в гиппокампе. Существует три основных подтипа ионотропных глутаматных рецепторов: NMDA (чувствительны к N-метил-D-аспартату), AMPA (связываются с -амино-3-гидрокси-5-изоксазолпропионовой кислотой) и каинатные рецепторы. NMDA- и AMPA–рецепторы играют ключевую роль в возникновении и проявлении долговременной потенциации. NMDA–рецепторы обладают значительной проницаемостью для ионов Са, однако при нормальном уровне МПП они заблокированы ионами Mg. Магниевый блок снимается при сильной деполяризации мембраны. AMPA рецепторы проницаемы для ионов Na+ и К+, и ответственны за возникновение ПСП в глутаматергических синапсах. Механизм долговременной потенциации При высокочастотной стимуляции нейронов гиппокампа выделяется большое количество глутамата, деполяризуется постсинаптическая мембрана и происходит активация NMDA–рецепторов. Значительный кальциевый ток через эти каналы приводит к повышению концентрации ионов Са2+ в постсинаптическом нейроне. Ионы Са2+ связываются с внутриклеточным белком — кальмодулином. Образовавшийся комплекс активирует фермент — Са2+-кальмодулинзависимую протеинкиназу II. Са2+-кальмодулинзависимая протеинкиназа II фосфорилирует AMPA–рецепторы. После фосфорилирования возрастает их ионная проводимость, что приводит к увеличению постсинаптического ответа на каждый квант нейромедиатора. Кроме того, данный фермент мобилизует дополнительные AMPA–рецепторы из цитоплазмы в постсинаптическую мембрану, что приводит к увеличению квантового состава ПСП. Долговременная депрессия также возникает в ответ на высокочастотное раздражение и проявляется в виде длительного ослабления синаптической передачи. Этот вид пластичности имеет сходный механизм с долговременной потенциацией, но развивается при более низкой внутриклеточной концентрации ионов Са2+. В заключение приведём данные о продолжительности различных событий, происходящих в синапсах на их постинаптической стороне: из рис. 6–11 видно, что продолжительность процессов имеет широкий разброс — от 1 мсек (деполяризация постсинаптической мембраны за счёт ионотропных рецепторов) до дней (модуляция синаптической передачи). 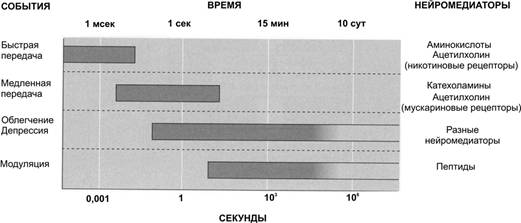 Рис. 6–11. Сравнительная продолжительность различных событий в синапсах [2]. Логарифмическая шкала. Влияние ацидоза, алкалоза и гипоксии на синаптическую передачу Алкалоз резко повышает возбудимость нейронов. Так, повышение рН артериальной крови с нормальных значений 7,4 до 7,7 может вызвать эпилептический приступ. Ацидоз значительно подавляет активность нейронов. Снижение рН с 7,4 до значений ниже 7,2 вызывает коматозное состояние (например, при тяжёлом сахарном диабете или уремическом ацидозе). Гипоксия может привести к полной потере возбудимости некоторых нейронов. Например, когда мозговой кровоток временно прекращается (на 3–7 сек), человек теряет сознание. |