Нф. ТЕОРИЯ Соматосенсорная система полости рта .. Сенсорная функция слизистой оболочки полости рта
 Скачать 0.91 Mb. Скачать 0.91 Mb.
|

|
Сенсорная функция слизистой оболочки полости рта. Слизистая оболочка покрывает все структуры (кроме зубов) полости рта и имеет значительную площадь. В определенных ситуациях она является протезным дожем, на который накладывают зубные протезы. Слизистая оболочка часто травмируется и поражается при стоматологических заболеваниях и заболеваниях внутренних органов, что отражается на ее сенсорной функции. Рецепторы слизистой оболочки полости рта представлены либо свободными нервными окончаниями в виде пластинок, пуговок, спиралей (первично-чувствующие рецепторы), либо специальными образованиями, например, вкусовыми клетками (вторично-чувствующие рецепторы). Тактильная рецепция. Тактильная рецепция слизистой оболочки полости рта обеспечивает возникновение ощущения прикосновения, давления и вибрации. Рецепторный отдел.К главным тактильным рецепторам относятся следующие механорецепторы - рецепторы Меркеля, Мейснера и Пачини (рис. 1). На прикосновение реагируют тельца Мейсснера, расположенные в поверхностных слоях собственно слизистой оболочки. При усилении механического раздражения реагируют диски Меркеля, расположенные в более глубоких слоях эпителиального слоя слизистой. Чувство давления, а также реакция на вибрацию (прерывистое давление) возникают при раздражении телец Пачини, располагающихся в глубоких слоях слизистой. Тактильные рецепторы находятся в строгой функциональной взаимосвязи с механорецепторами пародонта и с проприорецепторами жевательных мышц. Их взаимодействие определяет участие мышц в акте жевания. По функциональным особенностям тактильные рецепторы подразделяют на фазные и статические. 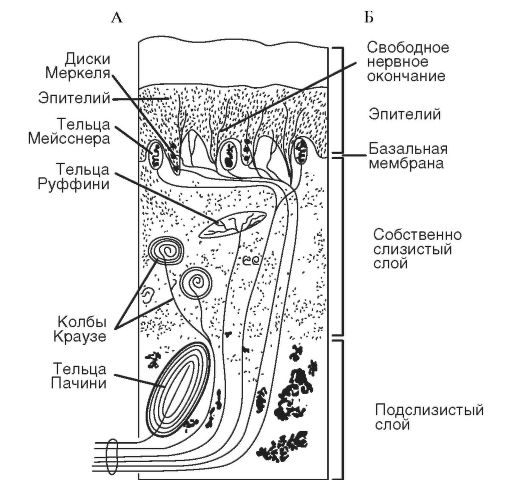 Рис 1. Рецепторы слизистой оболочки полости рта: А - виды рецепторов; Б - строение слизистой. Статические тактильные рецепторы возбуждаются в основном при продолжительном статическом раздражении, например, при наложении съемных протезов. Они менее чувствительны, чем фазные, обладают более длительным латентным периодом, медленно адаптируются. Фазные тактильные рецепторы возбуждаются при динамическом раздражении; они обладают высокой чувствительностью, коротким латентным периодом, быстро адаптируются. Эти рецепторы возбуждаются, например, при недостаточно прочной фиксации съемного зубного протеза. Свойства тактильных рецепторов играют большую роль в процессах адаптации к съемным зубным протезам. На спинке языка функцию осязания обеспечивают нитевидные сосочки. Они имеют вид конусообразных возвышений, плотно прилегают друг к другу, поэтому поверхность языка имеет бархатистый вид. Эпителий, покрывающий нитевидные сосочки, ороговевает. Слущивание поверхностного слоя эпителия нитевидных сосочков является выражением физиологического процесса регенерации. При нарушениях функции органов пищеварения, общих воспалительных и инфекционных заболеваниях отторжение поверхностного слоя эпителия нитевидных сосочков замедляется, и язык приобретает вид «обложенного», Тактильные рецепторы в различных отделах челюстно-лицевой области распределены неравномерно. Наиболее плотно они расположены на кончике языка, слизистой оболочке и красной кайме губ. Вероятно, это обусловлено тем, что данные образования являются первой инстанцией анализа механических свойств веществ, поступающих в полость рта. Верхняя губа (слизистая оболочка и красная кайма) более чувствительна к механическим раздражениям, чем нижняя. Сравнительно высок уровень тактильной чувствительности слизистой оболочки твердого неба. Это имеет особое значение при апробации пищи на съедобность (фаза ориентировочного жевания), а также при формировании пищевого комка и в начале глотания, поскольку информация от расположенных здесь рецепторов входит в состав пусковой аффереитации. Таблица 1. Пороги тактильной чувствительности слизистой оболочки десневых сосочков» мг/мм2
Наименьшей тактильной чувствительностью обладает слизистая оболочка вестибулярной поверхности десен. В области десневых сосочков установлен убывающий градиент чувствительности влево и вправо вглубь от центра альвеолярной дуги (табл. 1), причем с правой стороны чувствительность выше, чем с левой. Наличие асимметрии объясняется особенностями иннервации: наибольшее количество нервных рецепторных образований находится на правой стороне лица. Проводниковый отдел тактильного анализатора. От большинства механорецепторов информация в центральную нервную систему проводится по толстым (диаметр до 8-10 мкм) миелиновым волокнам типа А со скоростью 30 - 70 м/с. Первый нейрон находится в тройничном ганглии V нерва, второй - в спинальном и среднемозговом ядрах V нерва, третий - в ядрах таламуса. Центральный (корковый) отдел тактильной сенсорной системы. Первичная соматосенсорная область коры (SI) располагается в задней центральной извилине.S1 обладает высокой степенью топографической организации и содержит «топографическую карту» распределения тактильных рецепторов, в которой площади проекции отдельных зон кожи пропорциональны плотности тактильных рецепторов в этих зонах. Вторичная соматосенсорная зона коры (SII, филогенетически более древняя, чем зона SI), расположенная на верхней стенке боковой борозды (теменная покрышка), возле слуховой зоны. Сюда поступает афферентная импульсация как от противоположной стороны, так и от «своей» стороны; здесь имеется менее четкая соматотопическая проекция поверхности тела. Исследование тактильной чувствительности (эстезиометрия) проводят методом определения абсолютных порогов с помощью аппарата Фрея или пространственных порогов циркулем Вебера. Ф 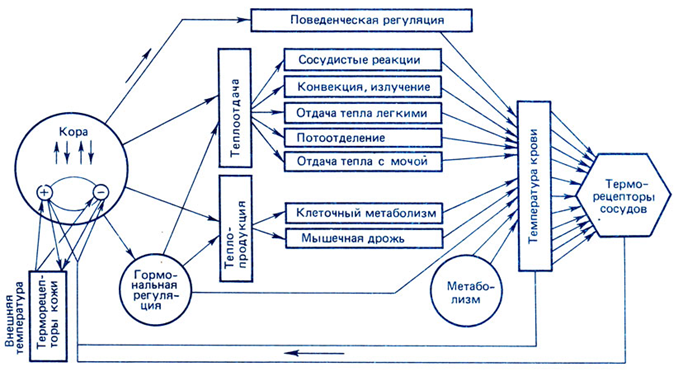 ункциональная система терморегуляции ункциональная система терморегуляцииРис. 2. Функциональная система поддержания температурного гомеостаза. К.В. Судаков, 2008. Системообразующий фактор. Если рассматривать терморегуляцию с позиций функциональной системы (рис. 2), то полезным результатом (системообразующим фактором) является поддержание температуры «ядра» тела (≈ 37оС) и средневзвешенной температуры кожи (≈ 33оС), которые определяют направление переноса тепла между организмом и средой. Температурная сенсорная система как звено афферентного синтеза (рис. 3). Рецепторный отдел. Периферические терморецепторы кожи, кожных и подкожных сосудов позволяют осуществить регуляцию по возмущению ещё до изменения температуры «ядра». Максимальная плотность и чувствительность терморецепторов имеется на главных участках теплоотдачи – лицо, голова, шея, минимальная – на коже нижних конечностей. Специализация рецепторов: холодовые, тепловые и полимодальные. Соотношение холодовых и тепловых рецепторов в коже равно 8:1. Глубина расположения холодовых рецепторов меньше (0,17 мм), чем у тепловых (0,3 мм), латентный период ответа у холодовых короче по сравнению с тепловыми. В зоне температурного комфорта импульсация от холодовых рецепторов 10 – 12 Гц, от тепловых – 2 – 4 Гц. Максимальная чувствительность и импульсация холодовых рецепторов находится в диапазоне температуры кожи 15 – 34оС, у тепловых – 34 – 43оС, при температуре ниже 20 и выше 44ºС происходит возбуждение болевых рецепторов. Функциональная мобильность терморецепторов является механизмом настройки температурного анализатора: чем ниже температура кожи, тем больше активных холодовых точек, при повышении температуры растет количество тепловых точек. У адаптированных к холоду количество активных холодовых точек меньше, чем у неадаптированных людей. 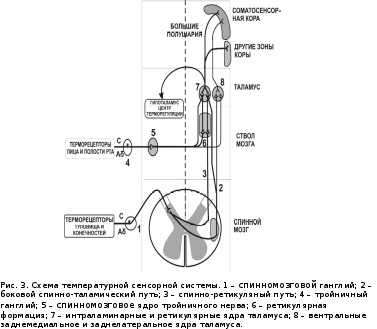 Сигналы от периферических терморецепторов интегрируются преимущественно в переднем гипоталамусе. Терморецепторы внутренних органов обеспечивают регуляцию по отклонению в ответ на изменение температуры «ядра». Терморецепторы ЦНС максимальное их количество содержится в преоптической области гипоталамуса, соотношение тепловых и холодовых 5:1 (дифференциальный порог их чувствительности ≈ 0,011оС), а также в спинном мозге (чувствительность в 4 раза ниже), ретикулярной формации, коре. Терморецепторы сосудов, желудка, верхних дыхательных путей (преобладают холодовые рецепторы). Проводниковый отдел: от холодовых рецепторов идет передача импульсов по волокнам Аδ и С, от тепловых – по волокнам С. От терморецепторов кожи туловища, шеи, конечностей и внутренних органов идут волокна 1‑х нейронов, находящихся в спинномозговых ганглиях. Их аксоны переключаются на 2‑е нейроны в задних столбах спинного мозга и после перекреста образуют на противоположной стороне два главных пути проведения температурной чувствительности. Боковой спинно-таламический путь с преимущественным переключением прямо или через нейроны ретикулярной формации в неспецифических ядрах таламуса часть волокон переключается в специфических (вентральных заднелатеральных) ядрах таламуса и обеспечивает осознание с участием коры постцентральной извилины температурного комфорта или дискомфорта. Спинно-ретикулярный путь дает диффузные окончания в ретикулярную формацию, неспецифические ядра таламуса, гипоталамус (преимущественно передний отдел, где обеспечивается интеграция сигналов от терморецепторов кожи, самого гипоталамуса и других структур). От терморецепторов кожи головы и лица по афферентным волокнам тройничного нерва тройничный ганглий (1-е нейроны) спинномозговое ядро V нерва (2‑е нейроны) специфическое (вентральное заднемедиальное) ядро таламуса (3-и нейроны). Часть импульсов идет в ретикулярную формацию и неспецифические ядра таламуса. Корковый отдел (постцентральная извилина больших полушарий). Обеспечивает восприятие температурных сигналов и осознание действия температурного раздражителя (комфорт или дискомфорт). Участвует в образовании условнорефлекторных реакций на термические раздражения, осуществляет регуляцию по прогнозированию или упреждению влияния на организм измененной температуры среды. Участвует в поведенческих механизмах терморегуляции. Гипоталамус – важнейшая нервная структура, в которой расположено «ядро» центра терморегуляции. В него поступает значительная часть импульсов из проводникового и коркового отделов температурного анализатора. В гипоталамусе формируется «установочная точка» и эфферентная программа, обеспечивающая терморегуляцию. Вместе с тем в центральных механизмах терморегуляции участвуют и другие отделы ЦНС – кора больших полушарий, продолговатый и спинной мозг, в которых также обнаружены термочувствительные нейроны (рис. 4). В гипоталамусе имеется нескольких видов терморегуляторных нейронов. Афферентные нейроны, принимающие информацию от периферических и центральных терморецепторов. Они расположены преимущественно в переднем гипоталамусе и участвуют в сопоставлении реальной температуры тела (и самого гипоталамуса) с установочной точкой, при которой теплопродукция и теплоотдача в организме уравновешены. Термочувствительные нейроны – холодовые и тепловые. Они расположены преимущественно в преоптической области и ядрах переднего гипоталамуса (хотя они обнаружены и в других ядрах гипоталамуса, а также в спинном и продолговатом мозге, коре больших полушарий). Тепловых нейронов в 4–6 раз больше, чем холодовых. Для термочувствительных нейронов характерна высокая чувствительность к изменению температуры (дифференциальный порог – 0,011°С) и высокая зависимость от температуры их импульсации и проводимости ионных каналов,  Повышение температуры активирует Nа+-каналы плазмолеммы тепловых нейронов, входящий Nа+-ток вызывает деполяризацию, достигающую КУД –нейрон приобретает пейсмекерные свойства. Снижение температуры инактивирует К+-каналы плазмолеммы холодовых нейронов, уменьшение выходящего из клетки К+-тока приводит к деполяризации мембраны, достигающей КУД. Эфферентные нейроны,реализующие эфферентную программу терморегуляции (теплопродукцию и теплоотдачу) расположены соответственно в заднем и переднем гипоталамусе. Раздражители гипоталамических нейронов. Афферентная импульсация от центральных и периферических терморецепторов. Гуморальные сигналы – гормоны щитовидной железы (тироксин, трийодтиронин), прогестерон, адреналин и др. Пирогены: экзогенные (бактериальные, вирусные) и эндогенные (цитокины – интерлейкины 1 и 6, ФНО и интерфероны β и , простагландины Е1) действуют на «холодовые» нейроны гипоталамуса и стимулируют теплопродукцию, одновременно угнетая теплоотдачу. Центр теплопродукции расположен в заднем гипоталамусе (вентролатеральные, промежуточные и вентромедиальные ядра). Его раздражение вызывает повышение температуры тела в результате усиления окисления глюкозы и жирных кислот, важную роль при этом играют ядра гипоталамуса, секретирующие тиролиберин (тиролиберин ТТГ тироидные гормоны). В дорсомедиальной части заднего гипоталамуса у стенки III желудочка обнаружен моторный центр дрожи, связанный с моторными центрами продолговатого и спинного мозга и вызывающий сначала повышение тонуса мышц, а затем – дрожь. Он возбуждается при снижении температуры гипоталамуса даже на доли градуса. Разрушение центра теплопродукции приводит к потере способности поддерживать температуру тела на холоде (пойкилотермия). Центр теплоотдачи расположен в переднем гипоталамусе (паравентрикулярные, супрахиазматические и супраоптические ядра, медиальная преоптическая область). Раздражение этих структур вызывает увеличение теплоотдачи в результате расширения сосудов кожи и повышения температуры кожи, потоотделения и тепловой одышки. Разрушение приводит к неспособности выдерживать тепловую нагрузку (перегрев). Эфферентные нейроны гипоталамуса управляют процессами теплопродукции и теплоотдачи: - от тепловых рецепторов возбуждаются эфферентные нейроны переднего гипоталамуса и активируются механизмы, обеспечивающие рост теплоотдачи - от холодовых рецепторов возбуждаются эфферентные нейроны заднего гипоталамуса и усиливается теплопродукция - между этими нейронами имеются перекрестные тормозные связи (реципрокное торможение). Эфферентные (исполнительные) механизмы терморегуляции изложены выше. Если дисбаланс теплопродукции и теплоотдачи небольшой, то он устраняется путем сосудистых механизмов регуляции теплоотдачи. При недостаточности этих механизмов включаются механизмы регуляции потоотделения. При более выраженном дисбалансе, связанном с охлаждением организма, используются механизмы усиления теплопродукции (несократительный и сократительный термогенез). Наиболее мощными средствами предупреждения и ликвидации возникшего дисбаланса являются поведенческие механизмы терморегуляции. Температурная рецепция полости рта. Рецепторный отдел. Тепловые рецепторы гистологически представлены тельцами (гроздьями) Руффини, холодовые - колбами Краузе. Рецепторы холода расположены в эпителии или непосредственно под ним, рецепторы тепла - преимущественно в нижнем и верхнем слоях собственно слизистой оболочки. Эти особенности обусловливают более высокую чувствительность слизистой оболочки к холоду, нежели к характерно теплу. Для тепловой чувствительности наличие возрастающего градиента от передних отделов полости рта к задним отделам, а для холодовой чувствительности -убывающего градиента чувствительности в этом же направлении (рис. 5). Преобладание холодовых рецепторов в передних отделах полости рта, а тепловых - в задних обусловлено специфичностью их функций и значимостью в процессах терморегуляции организма. Холодовая сенсорная система, являясь ведущей в терморегуляции организма, быстрее и адекватнее откликается на изменение температуры внешней среды, в то время как тепловая афферентация сигнализирует в основном о температурном гомеостазе самого организма. Слизистая оболочка щек малочувствительна к холоду и еще меньше - к теплу. Восприятие тепла полностью отсутствует в центре твердого неба, а центральная часть задней поверхности языка не воспринимает ни холодовые, ни тепловые воздействия. Высокой чувствительностью к температурным раздражениям обладают кончик языка и красная кайма губ. При приеме пищи в первую очередь раздражаются именно эти области. Информация о температуре веществ от этих областей в случае необходимости может включать соответствующие защитные реакции. 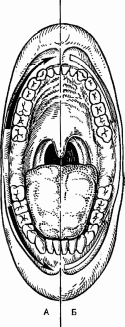 Рис. 5. Распределение температурной чувствительности слизистой оболочки преддверия полости рта. Стрелками обозначены: А — градиент тепловой чувствительности; Б — градиент холодовой чувствительности Проводниковый отделы температурного анализатора.От рецепторов холода отходят тонкие миелиновые волокна типа Аδ диаметром 1 - 4 мкм и скоростью проведения возбуждения 5 - 15 м/с, а от рецепторов тепла — безмиелиновые волокна типа С диаметром 0,5 - 1 мкм и скоростью проведения 0,5 - 2 м/с. Чувствительные ганглии, где расположен I нейрон, относятся к соответствующим нервам, которые иннервируют различные участки слизистой оболочки полости рта. Локализация II, III, а также IV нейрона (корковый отдел) температурного анализатора соответствует тактильному анализатору, поскольку оба они относятся к соматосенсорному виду чувствительности. Исследование температурной чувствительности проводят методом холодовой или тепловой термоэстезиометрии путем определения дифференциальных порогов, а также методом функциональной мобильности. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||