Система комплемента. Система комплемента. Функции компонентов системы, роль во врожденном и адаптивном иммунитете
 Скачать 326.5 Kb. Скачать 326.5 Kb.
|

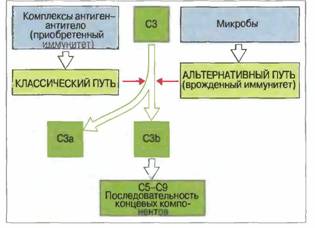
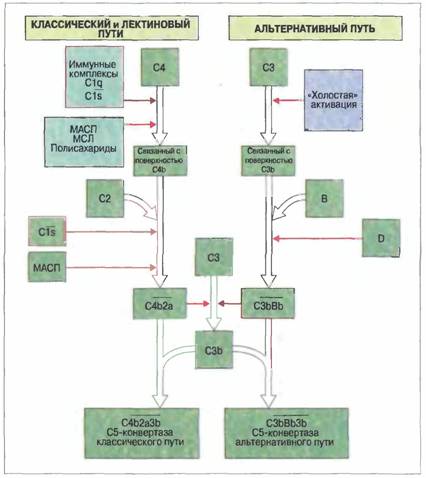
Федеральное Государственное Образовательное УчреждениеВысшего Профессионального Образования«Московская государственная академия ветеринарной медициныи биотехнологий им. К. И. Скрябина»Реферат по дисциплине «Иммунология» на тему: «Система комплемента. Функции компонентов системы, роль во врожденном и адаптивном иммунитете» Выполнила: студентка группы 3 курса дневного отделения ФВМ Москва 2008. План: Введение……………………………………………………………………….…………………………..3 Структура белков комплемента………………………………………………………………..……...….5 Активация комплемента………………………………………………………………………..……..…..6 Рецепторы комплемента…………………………………………………………..........................……..13 Биологические эффекты комплемента……………………………………………..…………………...15 Список используемой литературы……...…………………………………………...………………….20 Введение.Термин «комплемент» первоначально применил Эрлих для описания «дополнительной», присутствующей в сыворотке активности, без которой специфичные антитела не могут лизировать бактерии. Комплемент впервые описал Бухнер в 1889 г. под названием «алексин»— термолабильный фактор, в присутствии которого наблюдается лизис микробов. 1907 г. Феррата, диализуя сыворотку против подкисленной воды, установил, что белки комплемента можно разделить на две фракции: выпадающие в осадок эуглобулины и водорастворимую альбуминовую фракцию (псевдоглобулины). Активность комплемента проявлялась только в присутствии обеих фракций, названных тогда средней и концевой частями, а позднее — С'1 и С'2 компонентами. В последующем Сакс и Омороков обнаружили, что яд кобры инактивирует другой компонент комплемента (С'3), а Гордон установил, что следующий компонент (С'4) разрушается аммиаком. Последовательность открытия перечисленных компонентов комплемента не соответствует очередности их вступления в реакцию активации системы, и этим объясняется кажущаяся нелогичность ее современной номенклатуры. Система комплемента — сложный комплекс белков, представленных главным образом во фракции β-глобулинов, насчитывающий, включая регуляторные, около 20 компонентов, на долю которых приходится 10 % белков сыворотки крови. Номенклатура системы комплемента.Белки классического пути активации и лизирующего мембрану комплекса обозначены каждый своим номером и вступают в реакцию активации в следующем порядке: C1q, C1r, C1s, С4, С2, С3, С5, С6, С7, С8, С9. Среди них много предшественников ферментов — проферментов, которые приобретают активность только после расщепления. Обозначение активного фермента отличается от обозначения его неактивного предшественника надбуквенной чертой. Продукты расщепления обозначаются так же, как исходные компоненты комплемента, но с добавлением строчных букв - обычно для меньшего фрагмента — «а», а для большего — «b», например С3а и С3b. Из этого правила имеется одно исключение: С2b означает меньший, а С2а — больший фрагмент С2. Белки альтернативного пути активации называют факторами и обозначают однобуквенными символами. В тексте слово фактор обычно сокращается до первой буквы F или вовсе опускается. Регуляторные белки чаще всего обозначают аббревиатурами названий их функциональной активности: например, белок, ускоряющий диссоциацию С3-конвертазы классического пути, имеет символ DAF (decay accelerating factor), или, по-русски, ФУД (фактор ускорения диссоциации). Клеточные рецепторы, связывающие компоненты комплемента, названы по аббревиатурам своих лигандов (например, С5а-рецептор) или как маркерные молекулы в номенклатуре CD-системы. Отдельно пронумерованы рецепторы для главных фрагментов С3 как рецепторы комплемента типов 1, 2, 3 и 4 (CR1, CR2, CR3 и CR4). К сожалению, в результате этого некоторые рецепторы в современной литературе имеют по три синонима, например С3b-рецептор = CR1 = CD35. Система комплемента относится к факторам врожденного иммунитета и включает в себя ряд белков, действующих последовательно, т. е. каскадом, в котором каждый фермент катализирует активность следующего. Наиболее важный компонент комплемента — это С3, присутствующий в плазме крови в той же концентрации (1—2 мг/мл), что и некоторые иммуноглобулины. Два главных пути активации комплемента отражают особенности его участия в реакциях врожденного и приобретенного иммунитета. Классический путь связан с приобретенным иммунитетом, поскольку белок C1q взаимодействует с антителами, образовавшими комплекс с антигеном. Альтернативный путь активации комплемента относится к механизмам врожденного иммунитета, начинаясь иммунонеспецифическим связыванием С3b с поверхностью микроорганизма. Активность отдельных компонентов комплемента in vivo можно проиллюстрировать на примерах расстройств, вызванных недостаточностью этих белков. У таких больных наблюдается повышенная восприимчивость к рецидивирующим гнойным бактериальным инфекциям, а также к заболеваниям, для которых характерно повышенное образование аутоантител и иммунных комплексов. Эти наблюдения свидетельствуют о необходимости комплемента как для антибактериальной защиты, так и для устранения иммунных комплексов, которые иначе способны вызывать аутоиммунные заболевания и болезни иммунных комплексов. В результате активации комплемента при воспалении происходят: • опсонизация микроорганизмов и иммунных комплексов; • активация лейкоцитов; • лизис клеток-мишеней. Опсонизация Это стимуляция фагоцитоза в результате прикрепления белков комлемента к поверхности мишеней (микробов, иммунных комплексов и др.). Обладая рецепторами к опсонизирующим белкам, фагоцитарные клетки связывают мишени, что вызывает активацию фагоцитов и эндоцитоз или фагоцитоз мишеней. Активация лейкоцитов Полиморфноядерные гранулоциты и макрофаги обладают специфическими рецепторами к мелким фрагментам белков комплемента, образующимся на поверхности мишеней в результате каскада протеолитических реакций. Диффундируя в окружающую среду, эти фрагменты привлекают фагоциты (направленное движение клеток, или хемотаксис) и, связываясь с ними, вызывают их активацию. Лизис клеток-мишеней Протеолитический каскад комплемента завершается погружением гидрофобного «зонда» в липидный бислой мембраны клетки-мишени и ее последующим осмотическим разрывом и лизисом. Комплемент способен отличать «свое» от «не-своего»Относясь к факторам врожденного иммунитета, комплемент реализует механизмы, позволяющие отличать «свое» от «не-своего». Ключевой момент этой функции заключается в немедленном связывании С3b со всеми чужеродными объектами, будь то микроорганизмы или иммунные комплексы; поверхность собственных клеток организма защищена особыми молекулами, которые весьма эффективно ограничивают отложение С3b. Структура белков комплемента. Комплемент представляет собой систему каскадно-действующих пептид-гидролаз, получивших обозначение от С1 до С9. Установлено, что большая часть компонентов комплемента синтезируется гепатоцитами и другими метками печени (около 90 %, СЗ, С6, С8, фактор В и др.), а также моноцитами/макрофагами (С1, С2, СЗ, С4, С5). Белки системы комплемента относятся к различным суперсемействам. Белки, объединенные в одно суперсемейство — например иммуноглобулинов — имеют много общих структурных и функциональных свойств.Классификация белков комплемента по суперсемействам позволяет лучше понять их структурные и функциональные взаимосвязи.Например, суперсемейство регуляторных белков комплемента, называемых также регуляторами активации комплемента. К ним относятся: • фактор Н — белок плазмы крови с молекулой удлиненной конфигурации; • С4-связывающий белок [Cd-bp (binding protein)] — гептамерный белок плазмы, молекула которого имеет паукообразную форму; • фактор, ускоряющий диссоциацию С3-конвертазы (ФУД, CD55), — белок клеточной мембраны, закрепленный в ней на своеобразной гликофосфолипидной «ножке»; • мембранный кофакторный белок (МКБ, CD46) — трансмембранный белок, действующий как кофактор расщепления С3b: • рецепторы комплемента 1 типа (CR1, CD35) и 2 типа (CR2, CD21) — клеточные рецепторы, имеющие трансмембранные домены. Семейство регуляторных белков комплемента кодирует группа тесно сцепленных генов, расположенных в хромосоме 1. При очевидных различиях структуры все эти белки содержат одинаковый домен, состоящий примерно из 60 аминокислотных остатков и названный коротким общим повтором. Этот домен может много раз встречаться в структуре каждой молекулы, образуя ее каркас и, возможно, определяя специфичность связывания. Синтез этих белков кодируют гомологичные, тандемно расположенные экзоны. Составляющие это семейство шесть белков выполняют также ряд общих функций в активации комплемента: фактор Н, С4-bр, ФУД, МКБ и CR1 подавляют образование комплексов С4b2а и С3bВb, т. е. С3-конвертаз классического и альтернативного путей активации. Некоторые из этих белков имеют и другие общие функции, но не идентичные, а лишь частично перекрывающиеся. Такие функции включают: подавление связывания С2 с С4b и фактора В с С3b, индукцию диссоциации С2а от С4b и Вb от С3b, действие в качестве кофакторов фактора I — фермента, ответственного за катаболизм С3b и С4b. Следует отметить, что короткие общие повторы имеются и в других белках, которые, однако, не взаимодействуют с белками комплемента; это рецептор для ИЛ-2, β2-гликопротеин I и фактор XIII системы свертывания крови. Структура большинства белков комплемента «мозаична». Молекулярная основа родства белков внутри разных семейств становится яснее благодаря клонированию их генов. По современным представлениям, в ходе эволюции происходила многократная дупликация экзонов и их «перетасовка» между различными генами. Находясь одновременно в составе разных генов, эти дуплицированные сегменты ДНК эволюционировали параллельно и во многих случаях сохранили сходные последовательность и функцию, хотя в ряде случаев активность утрачена или приобретена новая.Многие белки комплемента представляют собой «мозаику» из продуктов экзонов, относящихся к генам разных суперсемейств. Так, C1s, фермент классического пути, содержит участки аминокислотной последовательности из сериновой эстеразы и рецептора для липопротеинов низкой плотности, а также короткий общий повтор, встречающийся в суперсемействе регуляторных белков комплемента. Точно так же, С6, С7, С8 и С9 — компоненты лизирующего мембрану комплекса — имеют общие свойства с перфорином цитотоксических Т-лимфоцитов и катионным белком эозинофилов. Активация комплемента. Существует три пути (механизма) активации комплемента: классический, лектиновый и альтернативный. Все они ведут к образованию конвертазы, расщепляющей С3 на С3а и С3b, — центральный момент любого из каскадов комплемента (рис. 1). У позвоночных усложнение системы комплемента шло параллельно с повышением общего уровня организации, тканевой дифференцировки и совершенствования реакций врожденного и приобретенного иммунитета. Однако уже у круглоротых (миног и миксин) - низшего таксона ныне живущих позвоночных - система комплемента представлена альтернативным и лектиновым путями, а у эволюционно более продвинутых хрящевых рыб впервые появляется классический путь активации комплемента. Конвертаза классического и лектинового путей представляет собой комбинацию фрагментов С4 и С2 — С4b2а, тогда как конвертаза альтернативного пути — это комплекс С3 с FB — С3bВb. Фрагмент С3b, который отщепляют от С3 обе конвертазы, связывается с мембраной мишени и становится фокусом дополнительного образования С3b — эта ступень каскада получила название петля усиления. Рис. 1. Сопоставление классического и альтернативного путей активации комплемента
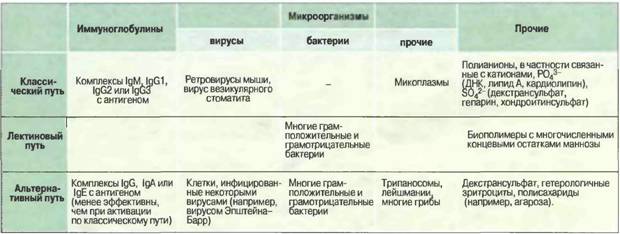
Активация комплемента как по классическому, так и по альтернативному пути приводит к появлению С3 конвертазы, которая превращает С3 в С3b, и эта конверсия - центральное событие всего каскада. В свою очередь, С3b активирует цепочку концевых компонентов комплемента (С5-С9), образующих литический комплекс. При активации по классическому пути сначала антиген связывается со специфическими антителами и только затем происходит фиксация С3. В альтернативной активации антитела не участвуют. Она начинается ковалентным связыванием С3b с гидроксильными группами на цитоплазматической мембране микробной клетки. Активация по альтернативному пути служит механизмом неспецифического врожденного иммунитета, тогда как классический путь представляет собой связующее звено между врожденным и приобретенным иммунитетом, появившееся в филогенезе сравнительно недавно. Присоединяя дополнительно молекулу С3b, обе С3-конвертазы могут превращаться в конвертазу С5, которая функционирует как катализатор на первой ступени каскада, ведущего к образованию лизирующего мембрану комплекса. Классический путь активации комплемента.Классический путь активации комплемента чаще всего запускается иммунными комплексами; роль первого ферментного комплекса в нем выполняет белок С1 (табл.1).Активацию инициирует связывание С1 с антителами в составе иммунных комплексов. Ферментный комплекс С1 состоит из 5 молекул — одной C1q, двух C1r и двух С1s; их соединение зависит от Са2+. Первая ступень каскада активации по классическому пути — это связывание антитела не менее чем с двумя из шести сферических доменов молекулы C1q. С1q состоит из 18 полипептидных цепей трех типов (по 6 цепей А-, В- и С-типов). Все 18 цепей своими коллагеноподобными N-концами (78 остатков аминокислот) образуют канатообразную спирально закрученную структуру, от которой расходятся в разные стороны С-концевые участки цепей (103—108 аминокислотных остатков), завершающиеся глобулярными головками, которые могут взаимодействовать с комплементсвязывающими участками Сн2-доменами (части Fc-областей) агрегированных молекул IgG в составе комплекса с антигеном. Молекулы C1q могут также связываться СнЗ-доменами неагрегированной молекулы IgM, конформация которой изменилась с «плоской» на «сложенную» в результате образования комплекса с антигеном. Предполагается, что многоточечное связывание сферических доменов C1q с входящими в иммунные комплексы молекулами IgG или IgM ведет к изменению конформации всего комплекса С1, вызывая автокаталитическую самоактивацию сначала одной, а затем и другой молекул C1r с превращением их в две молекулы активного фермента C1r, которые расщепляют обе молекулы С1s c образованием соответственно двух молекул C1s, обладающих активностью сериновой эстеразы. Табл. 1. Активаторы комплемента.
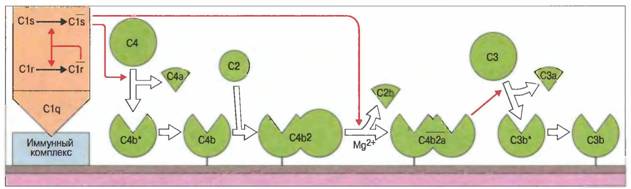
Лектиновый путь активации комплемента.Он почти идентичен классическому, но запускается независимо от антител.Белок C1q относится к семейству кальций-зависимых лектинов, названных коллектинами (коллагеновые лектины). В это же семейство белков входят маннан-связывающий лектин (МСЛ), называемый иначе маннан-связывающим белком (маннозосвязывающий белок, МСБ), конглютинин и легочные поверхностноактивные белки А и D. Сывороточный МСЛ может связываться с концевыми маннановыми группами на поверхности клеток бактерий, приобретая за счет этого способность к взаимодействию с двумя маннан-связывающими лектин-ассоциированными сериновыми протеиназами, МАСП1 и МАСП2, гомологичными по структуре С1r и С1s. Это взаимодействие подобно взаимодействию C1qcC1rnC1sn приводит к независимой от антител активации комплемента по классическому пути. Кроме того, C1q связывается непосредственно, т.е. без участия антител, с некоторыми микробами, в частности с микоплазмами и рядом ретровирусов (но не ВИЧ). Под действием С1 происходит расщепление С4 с образованием активированного С4b Белок С4 комплемента содержит внутреннюю тиоэфирную связь, участок расположения которой высокогомологичен тиоэфир-содержашему участку С3. При расщеплении С4 под действием C1s возникает два фрагмента: С4а. обладающий слабой анафилатоксической активностью, и более крупный (нестабильный, промежуточный), С4b*. (Звездочка указывает на нестабильное состояние молекулы, в которой активирован участок связывания). В течение нескольких миллисекунд С4b* подвергается атаке расположенных в непосредственной близости нуклеофильных групп. Большинство молекул С4b* гидролизуется с образованием инактивированного iC4b. Однако С4b* может образовывать ковалентные связи с амино- или гидроксигруппами молекул клеточной мембраны, превращаясь в связанный на поверхности С4b. Известны два изотипа С4 — С4А и С4В. Их кодируют расположенные тандемно гены главного комплекса гистосовместимости. Активированный С4А взаимодействует преимущественно с аминогруппами, а С4В — с гидроксируппами, образуя соответственно амидные и эфирные связи. Таким образом, С4А связывается в основном с белками, а С4В с углеводами. Связанный на клеточной поверхности С4b становится, в свою очередь, участком связывания для профермента С2 (в присутствии Мg2+) . Связанный С2 служит субстратом для С1s, который расщепляет его с освобождением С2b, при этом более крупный фрагмент, С2а, остается присоединенным к С4b, в результате чего образуется С4b2а — ферментный комплекс, называемый С3-конвертазой классического пути.Полипептид С3 относится к белкам с необычными посттрансляционными изменениями структуры. Расположенные на близком расстоянии остатки цистеина и глутамина образуют за счет элиминации аммиака метастабильную внутреннюю тиоэфирную связь. Электрофильная (акцептирующая электроны) карбонильная группа (—С+=0) этого тиоэфира чувствительна к атаке нуклеофильных групп (доноры электронов), в том числе амино- и гидроксигрупп приближающихся белковых и углеводных молекул. Таким образом, С3 способен ковалентно связываться с этими молекулами. Протеолитическое отщепление С3а от N-конца α-цепи С3 под действием С3-конвертазы приводит к конформационному изменению оставшейся части молекулы (т.е. С3b*), делающему внутреннюю тиоэфирную связь весьма нестабильной. Она становится новым участком связывания внутри С3b*, способным очень активно взаимодействовать с находящимися вблизи нуклеофильными группами. Как и в случае С4b*, большая часть молекул С3b* подвергается гидролизу, однако некоторые молекулы связываются с белками и углеводами, находящимися в непосредственной близости от места активации. Поскольку С3-конвертаза обычно образуется на чужеродной поверхности или на иммунных комплексах, С3b накапливается в основном там же. Затем связанный С3b становится фокусом дальнейшей активации комплемента по так называемой петле усиления альтернативного пути (рис. 2). Рис. 2. Классический путь активации комплемента
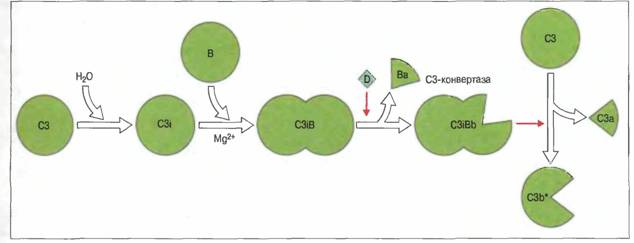
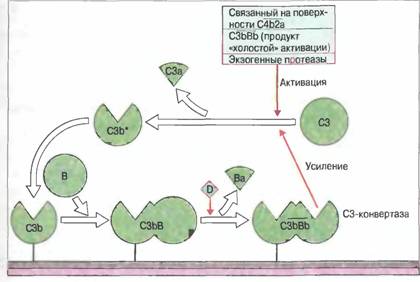
Существует два механизма регуляции классического пути активации комплемента в жидкой фазе. Первый — это действие С1-ингибитора, т.е. ингибитора сериновых протеиназ (серпина), связываюшего и инактивирующего С1r и С1s. Второй механизм состоит в подавлении образования С3-конвертазы классического пути, С4b2а. В жидкой фазе так действуют фактор I и С4-связывающий белок (С4-bр), вместе расщепляющие С4b. Кроме того, С4-bр вызывает диссоциацию С4b2а на С2а и С4b. Активация по классическому пути регулируется также путем подавления взаимодействия комплемента с поверхностью клеток хозяина. Ингибирование осуществляют регуляторные белки комплемента: фактор, ускоряющий диссоциацию С3-конвертазы (ФУД, CD55), рецепторы комплемента 1 типа (CR1, CD35) и мембранный кофакторный белок (МКБ, CD46). Эти белки действуют следующим образом: • подавляют связывание С2 с С4b (ФУД или CR1); • вызывают и ускоряют диссоциацию С4b2а на С2а и С4b (ФУД и CR1); • действуют как кофакторы, стимулируя катаболизм С4b под действием фактора I (МКБ или CR1). Существует самопроизвольная активация комплемента по альтернативному пути. «Холостая» альтернативная активацию комплемента постоянно поддерживает в плазме крови небольшую концентрацию С3b*.Внутренняя тиоэфирная связь в нативной молекуле С3 чувствительна к спонтанному гидролизу с превращением в активированную форму — C3i. (Эта постоянная, происходящая на низком уровне, самопроизвольная активация С3 в плазме крови называется «холостой».) Образующийся C3i связывает фактор В с образованием C3iB (рис. 3). Аналогичным образом, С2 связывается с С4b (рис. 4).Связанный фактор В расщепляется фактором D с высвобождением Ва. Оставшийся комплекс C3iBb представляет собой жидкофазную С3-конвертазу альтернативного пут, расщепляющую С3 на С3а и С3b. Запуск петли усиления альтернативного пути связанным на поверхности аутологичных клеток С3b предотвращают регуляторные белки комплемента.
Поскольку С3-конвертаза альтернативного пути действует в жидкой фазе, большая часть образовавшегося в результате ее активности С3b* гидролизуется и инактивируется водой. Однако в случае контакта с чужеродной поверхностью, в частности с мембраной бактериальной клетки, С3b* ковалентно связывается и инициирует действие петли усиления альтернативного пути. Общая схема взаимодействия компонентов комплемента при активации по классическому, лектиновому и альтернативному механизмам представлена на рис. 4. Как классический, так и альтернативный путь активации комплемента приводит к появлению С3-конвертазы: С4b2а и С3bВb соответственно. Классический путь начинается с активации Cis комплексом антиген-антитело и последующего расщепления активированным С1s компонентов С4 и С2. фрагменты меньшего размера, С4а и С2b, высвобождаются, а более крупные образуют С4b2а. Компоненты С4 и С2 могут быть активированы также МАСП (маннан-связывающей лектин-ассоциированной сериновой протеиназой) - белком лектинового пути, аналогичным Cis, и МСЛ (сывороточным маннан-связывающим лектином). На первых этапах альтернативного пути возникший в результате «холостой» активации и связавшийся с поверхностью белок С3b соединяется с фактором В, от которого фактор D отщепляет меньший фрагмент - Ва. Больший фрагмент В, т.е. Вb, остается связанным с С3b, образуя С3bВb - С3-конвертазу, которая расщепляет дополнительное количество молекул С3 (механизм положительной обратной связи). Поверхность, активирующая комплемент (например, микроорганизмов), стабилизирует С3b, обеспечивая его связывание с фактором В. Это способствует дальнейшей альтернативной активации комплемента. С3-конвертазы классического и альтернативного путей могут дополнительно присоединять С3b, образуя ферментные комплексы, называемые С5-конвертазами (С4b2аЗb и С3bВbЗb соответственно), которые активируют следующий компонент системы комплемента - С5. Поверхности, интенсивно активирующие комплемент, названы защитными, поскольку связанный с ними С3b защищен от протеолиза. Чужеродная поверхность, подобная мембране бактериальной клетки, «защищает» С3, поскольку, связавшись с ней, он проявляет более высокую афферентнность к фактору В, чем к фактору Н, и образует, вероятно, более стабильную конвертазу. Кроме того, на чужеродной поверхности отсутствуют регуляторные белки организма-хозяина, ингибирующие активацию комплемента. Рис.4 Аналогичные этапы активации комплемента по классическому, лектиновому и альтернативному механизмам
Хотя не совсем ясно, каковы конкретные структурные характеристики, необходимые для того, чтобы поверхность была защитной, особое значение имеет, по-видимому, ее углеводный состав. Например, присутствие кислых сахаров, в частности сиаловой кислоты, способствует, по-видимому, защите мембраны собственных клеток организма от повышенного отложения С3b. За первоначальным прикреплением одной молекулы С3b к «защитной» поверхности следует стадия амплификации, в результате которой в том же месте фиксируется много дополнительных молекул С3b. Ключевым моментом для быстрого накопления С3b служит образование мембраносвязанной С3-конвертазы. «Петля усиления» Петля усиления - это механизм положительной обратной связи в активации комплемента по альтернативному пути.Связанный с поверхностью С3b присоединяет фактор В. Образовавшийся С3bВ становится субстратом для фактора D — сериновой эстеразы, отщепляющей от фактора В небольшой фрагмент, Ва.. Остающийся на поверхности комплекс С3bВb весьма быстро диссоциирует, если не будет стабилизирован в результате связывания пропердина (Р) с образованием комплекса С3bВbР, представляющего собой связанную с поверхностью С3-конвертазу альтернативного пути. Комплекс С3bВbР расщепляет много все новых молекул С3. Поскольку конвертаза локализована на «защитной» поверхности, образующиеся молекулы С3b* будут связываться именно там, а не в каком-либо ином месте (рис. 5). Петля усиления функционирует и в том случае, когда С3b фиксируется на поверхности в результате классической (зависимой от антител) активации комплемента. Рис.5. «Петля усиления»
Альтернативная активация в жидкой фазе, когда С3b не связан с поверхностью, жестко регулируется белками, подобными или идентичными тем, которые «сдерживают» классическую активацию комплемента. Гомологичный С4-связывающему белку фактор Н, ген которого входит в кластер RCA, вызывает диссоциацию Вb из его комплексов с C3i или С3b, а также действует в качестве кофактора при катаболизировании C3i и С3b с участием фактора I до С3с и С3dg. Регуляция механизма усиления чрезвычайно важна для организма. Если она не функционирует, усиление (как циклический процесс, протекающий по принципу положительной обратной связи) идет до полного расщепления всех молекул С3. (Впервые это наблюдали у больного с наследственным дефицитом регуляторного фермента — фактора I. В отсутствие фактора I петля усиления действует вплоть до превращения всех молекул С3 сыворотки больного в С3b). На мембранах собственных клеток организма как ФУД, так и CR1 ускоряют диссоциацию комплекса С3bВb с высвобождением С3b. И CR1, и МКБ действуют как кофакторы расщепления С3b фактором I. Полностью аналогичным образом ФУД, МКБ и CR1 регулируют активность С4b2а (С3-конвертазы классического пути), когда она связана с клеточными мембранами. Таким образом, судьба связанного на поверхности С3b — это наиболее важный этап в том неспецифическом механизме, с помощью которого система комплемента отличает «свое» от «несвоего». Для связанного С3b существуют две возможности: • Усиление: С3b связывает фактор В с образованием конвертазы, которая вызывает фиксацию все новых молекул С3b на той же самой поверхности. • Подавление: С3b расщепляется фактором I при участии одного из трех кофакторов: фактора Н (из плазмы), CR1 или МКБ (связанного на поверхности). Какая из этих возможностей реализуется, зависит от природы поверхности, связавшей С3b. Присутствие на аутологичной (в частности, клеточной) поверхности собственных молекул, таких как ФУД, CR1 и МКБ, эффективно ограничивает образование С3-конвертаз. Напротив, чужеродная поверхность, например мембрана бактериальной клетки, обеспечивает «защиту» для С3b, так как именно на ней фактор В имеет большую афферентность к С3b, чем фактор Н. В результате фиксация всего нескольких молекул С3b приводит к образованию относительно стабильной С3-конвертазы альтернативного пути — С3bВbР, ферментного комплекса, который вызывает связывание все новых молекул С3b в том же участке. Конечная фаза активации комплемента это образование лизирующего мембрану комплекса.Каскад реакций активации комплемента завершается образованием литического комплекса (лизируюший, или атакующий, мембрану комплекс, ЛМК) в результате ферментативного расщепления С5 — белка, гомологичного С3 и С4, но не содержащего внутренней тиоэфирной связи. Прежде чем подвергнуться расщеплению С5-конвертазой, С5 избирательно связывается с С3b в ее составе. С5-конвертаза классического пути — это трехмолекулярный комплекс, С4b2аЗb, в котором С3b, ковалентно присоединенный к С4b, обладает более высокой константой связывания с С5, чем С3b, связанный с другими молекулами клеточной поверхности. С5-конвертаза альтернативного пути представляет собой также трехмолекулярный комплекс — С3bВbЗb, в котором один С3b ковалентно связан с другим. При расщеплении С5 высвобождается небольшой пептидный фрагмент С5а — высокоактивный анафилатоксин. Лизирующий мембрану комплекс образуется путем неферментативной сборки С5b-9 Последующее формирование ЛМК происходит без участия ферментов. Компонент С5b связывается с С6 с образованием С5b6, который взаимодействует с С7, образуя комплекс С5b67. В результате связывания С7 гидрофильный С5b6 превращается в гидрофобный комплекс С5b67, способный преимущественно встраиваться в липидный бислой. К этому комплексу присоединяется С8 и затем последовательно до 14 мономеров С9. В результате формируется литический «зонд», или порообразующая молекула, первые электронные микрофотографии которой получили Хамфри и Дурмашкин. Хотя после присоединения С8 к С5b67 комплекс уже проявляет незначительную литическую активность, полное ее развитие зависит от полимеризованного С9. Сформированный гидрофобный комплекс С5b67 способен самопроизвольно внедряться в мембраны других клеток, расположенных вблизи от клеточной поверхности, где происходит первичная активация комплемента. Этот процесс «реактивного лизиса» в отсутствие регуляции способен принести вред собственным тканям организма. Ряд белков может угнетать «реактивный лизис» путем связывания с С5b67 в жидкой фазе, прежде чем произойдет его фиксация на мембранах собственных клеток организма. Из таких белков в плазме крови в наибольшей концентрации содержится S-белок, или витронектин. Образуемый им комплекс SC5b67 лишен способности внедряться в двойной липидный слой; такая способность отсутствует также и у комплекса С5b678, поскольку он связывается с липопротеинами низкой плотности (ЛПНП), если присоединение С8 к С5b67 происходит в жидкой фазе. Мембрана клеток организма-хозяина содержит белки, защищающие ее от лизиса под действием ЛМК.Эритроциты, как в свое время было установлено, легко лизируются гетерологичным комплементом и труднее — гомологичным. Основа такой видовой рестрикции стала понятной после открытия особых мембранных белков, которые защищают клетки собственного организма от лизиса под действием ЛМК. Два таких белка уже подробно изучены. Первый из них — это CD59, заякоренный гликофосфолипидной «ножкой» в мембранах многих клеток белок. Он связывается с С8 в составе комплекса С5b—8 и подавляет погружение и развертывание С9 в клеточной мембране. Второй белок — фактор гомологичной рестрикции (ФГР), проявляющий ту же активность, что и CD59, но при этом более слабый ингибитор внедрения в мембрану С9. ФГР (мол. масса 65 кДа) также связан с мембраной гликофосфолипидом; его аминокислотная последовательность пока не установлена. Примечательно, что клетки, имеющие ядро, в частности клетки собственной иммунной системы организма, более эритроцитов устойчивы к комплемент-зависимому лизису благодаря способности активно удалять ЛМК путем эндоцитоза и экзоцитоза тех фрагментов мембраны, в которые он проник. |