реферат. Сравнительная характеристика проведения нервного импульса по миелиновым и безмиелиновым волокнам
 Скачать 254.26 Kb. Скачать 254.26 Kb.
|

|
Министерство здравоохранения Ростовской области Государственное бюджетное профессиональное образовательное Учреждение Ростовской области «Ростовский базовый Медицинский колледж» Сестринское дело Реферат на тему: «Сравнительная характеристика проведения нервного импульса по миелиновым и безмиелиновым волокнам» Выполнила: студентка 1 курса 5 группы отделения Сестринское дело очно-заочная форма обучения Омельченко Е.Л. Преподаватель: Захарова О.Ф. Ростов-на-Дону 2020 год СОДЕРЖАНИЕ Введение 1. Структура нервного волокна. 2. Классификация нервных волокон. 3. Особенности строения миелиновых и безмиелиновых нервных волокон. 4. Механизм проведения возбуждения по нервному волокну. 5. Сравнительная характеристика проведения нервного импульса по миелиновым и безмиелиновым волокнам. 6. Законы проведения возбуждения по нервным волокнам. 7. Особенности проведения возбуждения в нервных волокнах. Заключение Список использованных источников ВВЕДЕНИЕ раздражимость Нервная ткань (textus nervosus) — совокупность клеточных элементов, формирующих органы центральной и периферической нервной системы. Обладая свойством раздражимости, нервная ткань обеспечивает получение, переработку и хранение информации из внешней и внутренней среды, регуляцию и координацию деятельности всех частей организма. В составе нервной ткани имеются две разновидности клеток: нейроны (нейроциты) и глиальные клетки (глиоциты). Первый тип клеток организует сложные рефлекторные системы посредством разнообразных контактов друг с другом и осуществляет генерирование и распространение нервных импульсов. Второй тип клеток выполняет вспомогательные функции, обеспечивая жизнедеятельность нейронов. Нейроны и глиальные клетки образуют глионевральные структурно-функциональные комплексы. Исходя из выше сказанного, понятно что нервная система животного интересна в изучении. Нервная ткань обладает свойствами раздражимости, возбудимости. 1. СТРУКТУРА НЕРВНОГО ВОЛОКНА Нервные волокна представляют собой отростки нейронов, с помощью которых осуществляется связь между нейронами, а также нейронов с исполнительными клетками. В состав нервного волокна входят осевой цилиндр (нервный отросток) и глиальная оболочка. По взаимоотношению осевых цилиндров с глиальными клетками выделяют два типа нервных волокон: безмиелиновые и миелиновые. Оболочку безмиелиновых волокон образуют шванновские клетки (леммоциты). При этом осевые цилиндры прогибают клеточную оболочку леммоцитов и погружаются в них. Клеточная мембрана обычно полностью окружает каждый осевой цилиндр и смыкается над ним, образуя сдвоенную мембрану (мезаксон).  Рис. 1. Схема строения миелинизированного нервного волокна. Оболочку миелиновых волокон образуют в периферической нервной системе также шванновские клетки, а в ЦНС — олигодендроциты. В отличие от безмиелиновых волокон в миелиновых волокнах мезаксон удлиняется и спирально закручивается вокруг осевого цилиндра, образуя слой миелина толщиной от долей мкм до 10 мкм (липидный футляр) вокруг осевого цилиндра. Миелиновая оболочка через равные участки (0,5— 2,0 мм) прерывается, образуя свободные от миелина небольшие участки — узловые перехваты Ранвье. Протяженность перехватов в волокнах периферической нервной системы находится в пределах 0,25—1,0 мкм, в волокнах ЦНС их длина может достигать 14 мкм. Участки волокон между узловыми перехватами называются межузловыми сегментами, они образованы слоем миелина. Основную часть миелина (78 % сухой массы) составляют липиды, в них на долю фосфолипидов приходится 42 %, цереброзидов — 28 %, холестерина — 25 %. Несмотря на метаболическую инертность миелина (особенно по обновлению холестерина), поддержание целостности его структуры требует затраты энергии и нарушение снабжения олигодендроцитов кислородом и питательными веществами быстро вызывает деструкцию миелина. Одной из основных функций глиальной оболочки нервных волокон является изолирующая функция, способствующая лучшему проведению биопотенциалов по отросткам нейронов. 2. КЛАССИФИКАЦИЯ НЕРВНЫХ ВОЛОКОН Наиболее распространена классификация по Дж.Эрлангеру и Х.Гассеру (1937), в которой волокна разделяют на три типа: А, В и С (табл.3). Волокна типа А и В являются миелиновыми, типа С — безмиелиновыми. Волокна А делят на 4 подгруппы: α, β, γ, δ. В периферической нервной системе к волокнам Аα относятся афферентные волокна от механорецепторов кожи, мышечных и сухожильных рецепторов, а также эфферентные волокна к скелетным мышцам. К Аβ принадлежат афферентные волокна от кожных рецепторов прикосновения и давления, от части мышечных и висцеральных рецепторов. Аγ представляют собой эфферентные волокна, через которые регулируется активность мышечных рецепторов. К Аδ относят афферентные волокна от части тактильных, температурных и болевых, а также суставных рецепторов. К волокнам типа В принадлежат преганглионарные волокна вегетативной нервной системы. К волокнам типа С относят постганглионарные волокна вегетативной нервной системы, афферентные волокна от некоторых болевых (вторичная боль), тепловых и висцеральных рецепторов. Таблица1. Типы волокон в нервах млекопитающих (по Эрлангеру—Гассеру)
Из данных, представленных в табл.3, видно, что средний диаметр каждого типа волокна снижается от типа А до С (каждый примерно в 2 раза по отношению к предыдущему). Соответственно этому снижается и скорость проведения возбуждения. Низкая скорость проведения нервного импульса в волокнах типа С связана с особенностями проведения возбуждения в безмиелиновых волокнах. Лабильность также уменьшается от волокон Аα до С и находится в обратной зависимости от продолжительности фазы абсолютной рефрактерности. Возбудимость тоже уменьшается от волокон Аα (наибольшая возбудимость) к волокнам С (наименьшая возбудимость). Например, пороговая сила электрического тока у волокон С в 30—50 раз больше, чем у волокон Аα. Исследование факторов, блокирующих нервную проводимость, показало, что к давлению наиболее чувствительны волокна А, к кислородному голоданию (гипоксии) — волокна В, к местным анестетикам — волокна С. Нервные волокна имеют две основные функции — проведение возбуждения и транспорт веществ, обеспечивающих трофическую функцию. 3. ОСОБЕННОСТИ СТРОЕНИЯ МИЕЛИНОВЫХ И БЕЗМИЕЛИНОВЫХ НЕРВНЫХ ВОЛОКОН Безмиелиновое волокно - представляет собой цепь леммоцитов, в которую вдавлено от 5 до 20 осевых цилиндров. Каждый осевой цилиндр прогибает цитолемму леммоцита и вдавливается в его цитоплазму. Сближенные участки цитолеммы образуют мезаксон (дословно – брыжейка аксона). Находятся они преимущественно в составе вегетативной нервной системы. Клетки олигодендроглии оболочек безмиелиновых нервных волокон, располагаясь плотно, образуют тяжи цитоплазмы, в которых на определенном расстоянии друг от друга лежат овальные ядра. В безмиелиновых нервных волокнах внутренних органов часто в одной такой клетке располагается не один, а несколько (10-20) осевых цилиндров, принадлежащих различным нейронам. Они могут, покидая одно волокно, переходить в смежное. Такие волокна, содержащие несколько осевых цилиндров, называются волокнами кабельного типа. При электронной микроскопии безмиелиновых нервных волокон видно, что по мере погружения осевых цилиндров в тяж леммоцитов последние одевают их как муфта. На поперечном срезе безмиелинового нервного волокна определяются: - в центре волокна – ядро леммоцита - по периферии – поперечные сечения осевых цилиндров, как бы подвешенные на мезаксонах. - по самой периферии волокна – базальная пластинка из соединительной ткани. Безмиелиновые нервные волокна относятся к волнам кабельного типа, они тонкие (5-7- мкм) и проводят нервные импульсы очень медленно (1-2- м/сек). Миелиновые нервные волокна значительно толще безмиелиновых. Диаметр поперечного сечения их колеблется от 1 до 20 мк. Они также состоят из осевого цилиндра, одетого оболочкой из леммоцитов, но диаметр осевых цилиндров этого типа волокон значительно больше, а оболочка сложнее. В сформированном миелиновом волокне принято различать два слоя оболочки: внутренний, более толстый, - миелиновый слой, и наружный, тонкий, состоящий из цитоплазмы леммоцитов и их ядер. Миелиновый слой содержит в своем составе липоиды, а поэтому при обработке волокна осмиевой кислотой он интенсивно закрашивается в темно-коричневый цвет. Все волокно в этом случае представляется однородным цилиндром, в котором на определенном расстоянии друг от друга располагаются косо ориентированные светлые линии - насечки миелина (incision myelini), ил и насечки Шмидта-Лантермана. Через некоторые интервалы (от нескольких сотен микронов до нескольких миллиметров) волокно резко истончается, образуя сужения - узловые перехваты, или перехваты Ранвье. Перехваты соответствуют границе смежных леммоцитов. Отрезок волокна, заключенный между смежными перехватами, называется межузловым сегментом, а его оболочка представлена одной глиальной клеткой. В процессе развития миелинового волокна осевой цилиндр, погружаясь в леммоцит, прогибает его оболочку, образуя глубокую складку. По мере погружения осевого цилиндра оболочка леммоцита в области щели сближается и ее два листка соединяются друг с другом своей внешней поверхностью, образуя двойную мембрану - мезаксон. При дальнейшем развитии миелинового волокна мезаксон удлиняется и концентрически наслаивается на осевой цилиндр, вытесняя цитоплазму леммоцита и образуя вокруг осевого цилиндра плотную слоистую зону - миелиновый слой. Так как оболочка леммоцита состоит из липидов и белков, а мезаксон представляет собой ее двойной листок, то естественно, что миелиновая оболочка, образованная его завитками, интенсивно окрашивается осмиевой кислотой. В соответствии с этим под электронным микроскопом каждый завиток мезаксона виден как слоистая структура, построенная из белков и липидов, расположение которых типично для мембранных структур клеток. На продольном сечении вблизи перехвата видна область, в которой завитки мезаксона последовательно контактируют с осевым цилиндром. Место прикрепления самых глубоких завитков его наиболее удалено от перехвата, это легко понять, если представить себе, что закручивание мезаксона идет в процессе роста осевого цилиндра и одевающих его леммоцитов. Естественно, что первые завитки мезаксона оказываются короче, чем последние. Длина отростков различна. Переплетаясь между собой, они образуют вокруг осевого цилиндра своеобразный воротничок и попадают на срезах то в поперечном, то в продольном направлении. В толстых волокнах, у которых область перехвата относительно коротка, толщина воротничка из отростков шванновских клеток больше, чем в тонких волокнах. Очевидно, аксон тонких волокон в перехвате более доступен для внешних воздействий. Снаружи миелиновое нервное волокно покрыто базальной мембраной, связанной с плотными тяжами коллагеновых фибрилл, ориентированных продольно и не прерывающихся в перехвате - невралеммой. Функциональное значение оболочек миелинового нервного волокна в проведении нервного импульса в настоящее время недостаточно изучено. Осевой цилиндр нервных волокон состоит из нейроплазмы - бесструктурной цитоплазмы нервной клетки, содержащей продольно ориентированные нейрофиламенты и нейротубулы. В нейроплазме осевого цилиндра лежат митохондрии, которых больше в непосредственной близости к перехватам и особенно много в концевых аппаратах волокна. С поверхности осевой цилиндр покрыт мембраной - аксолеммой, обеспечивающей проведение нервного импульса. Сущность этого процесса сводится к быстрому перемещению локальной деполяризации мембраны осевого цилиндра по длине волокна. Последнее определяется проникновением в осевой цилиндр ионов натрия (Nа+), что меняет знак заряда внутренней поверхности мембраны на положительный. Это, в свою очередь, повышает проходимость ионов натрия в смежном участке и выход ионов калия (К+) на внешнюю поверхность мембраны в деполяризованном участке, в котором восстанавливается при этом исходный уровень разности потенциалов. Скорость движения волны деполяризации поверхностной мембраны осевого цилиндра определяет быстроту передачи нервного импульса. Известно, что волокна с толстым осевым цилиндром проводят раздражение быстрее тонких волокон. Скорость передачи импульса миелиновыми волокнами больше, чем безмиелиновыми. Тонкие волокна, бедные миелином, и безмиелиновые волокна проводят нервный импульс со скоростью 1-2 м/сек, тогда как толстые миелиновые - 5-120 м/сек. Строение миелинового нервного волокна: - осевой цилиндр один, погружен в центральную часть цепи леммоцитов. - мезаксон длинный, закручен вокруг осевого цилиндра, образуя миелиновый слой. - цитоплазма и ядро леммоцита сдвигаются на периферию и образуют неврилемму нервного волокна. - снаружи расположена базальная пластинка. Так как основу плазмолеммы составляет билипидный слой, то миелиновую оболочку образуют наслоения липидных слоев, интенсивно окрашивающихся в чёрный цвет осмиевой кислотой. По ходу миелинового волокна видны границы соседних леммоцитов – узловые перехваты (перехваты Ранвье). Участок между 2 перехватами называется межузловым сегментом. В каждом межузловом сегменте отчётливо видны насечки миелина – прозрачные участки, в которых содержится цитоплазма леммоцита между витками мезаксона. Диаметр миелиновых нервных волокон 12-20 мкм, скорость проведения нервного импульса – большая (10-120 м/с). Высокая скорость проведения импульсов объясняется сальтаторным способом передачи – скачками от 1 перехвата Ранвье к другому. Таблица 2 «Сравнительная характеристика миелиновых и безмиелиновых волокон»
4. МЕХАНИЗМ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ ПО НЕРВНОМУ ВОЛОКНУ Биопотенциалы могут быть локальными (местными), распространяющимися с декрементом (затуханием) на расстояние, не превышающее 1—2 мм, и импульсными (ПД), распространяющимися без декремента по всей длине волокна — на несколько десятков сантиметров, например от мотонейронов спинною мозга по всей длине нервного волокна до мышечных волокон конечностей с учетом длины самой конечности. Распространение локальных потенциалов. Локальные потенциалы (препотенциал, рецепторный потенциал, возбуждающий постсинаптический потенциал — ВПСП) изменяют мембранный потенциал покоя, как правило, в сторону деполяризации в результате входа в клетку Na+ согласно электрохимическому градиенту. В результате этого между участком волокна, в котором возник локальный потенциал, и соседними участками мембраны формируется электрохимический градиент, вызывающий передвижение ионов. В частности, вошедшие в клетку ионы Na+ начинают перемещаться в соседние участки, а ионы Na+ на наружной поверхности клетки движутся в противоположном направлении. В итоге поляризация мембраны соседнего участка уменьшится. Фактически это означает, что локальный потенциал из первичного очага распространился на соседний участок мембраны. Он затухает на расстоянии 1—2 мм от очага первичной деполяризации, что связано с отсутствием ионных управляемых каналов на данном участке мембраны или неактивацией управляемых ионных каналов, продольным сопротивлением цитоплазмы волокна и шунтированием тока во внеклеточную среду через каналы утечки мембраны. 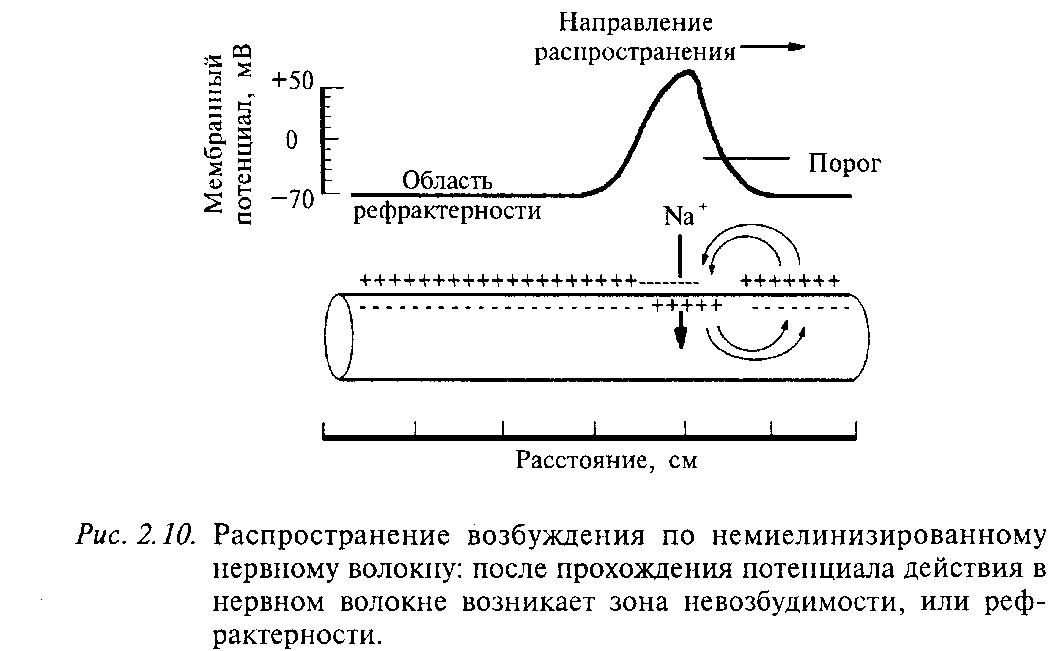 Если возникшая деполяризация мембраны не сопровождается изменением проницаемости потенциалзависимых натриевых, кальциевых и калиевых каналов, такую деполяризацию называют электротонической. Электротоническое распространение возбуждения — физический механизм, оно характерно для тех фрагментов мембран возбудимых клеток, где нет потенциалзависимых ионных каналов. Такими участками являются, например, большая часть мембраны дендритов нервных клеток, межперехватные промежутки в миелиновых нервных волокнах. Если местный потенциал (рецепторный или ВПСП), распространяясь электротонически, достигает участков мембраны, способных генерировать ПД (аксонный холмик, перехваты Ранвье, часть мембраны дендритов и, возможно, сомы), но его амплитуда при этом не достигнет критического уровня деполяризации, то такой потенциал называют препотенциалом. В его возникновении и распространении частично участвуют потенциалзависимые ионные каналы, однако при этом нет регенеративной (самоусиливающейся) деполяризации, характерной для ПД. Поэтому распространение такого потенциала происходит с затуханием амплитуды. Если локальный потенциал достигает участков мембраны, способных генерировать ПД, и его амплитуда выходит на критический уровень деполяризации, формируется ПД, который распространяется по всей длине нервного волокна без затухания. Эффективность электротонического распространения биопотенциалов зависит от физических свойств нервного волокна — сопротивления и емкости его мембраны, сопротивления цитоплазмы. Электротоническое проведение в нервном волокне улучшается при увеличении его диаметра, что связано с уменьшением сопротивления цитоплазмы, а также при миелинизации волокна, увеличивающей сопротивление мембраны (до 105 Ом/см2) и уменьшающей ее емкость (до 0,005 мкФ/см2). Эффективность электротонического проведения характеризует постоянная длины мембраны (λm). Это расстояние, на которое может электротонически распространиться биопотенциал, пока его амплитуда не уменьшится до 37 % от исходной величины. Постоянная длины для локальных потенциалов реально не превышает 1 мм, и их амплитуда затухает на расстоянии 1—2 мм от места возникновения. Для передачи возбуждения на большие расстояния необходимо формирование ПД. В его распространении, кроме электротонического механизма, существенная роль принадлежит механизму регенеративной деполяризации, позволяющей сохранить амплитуду ПД на всем пути его следования. Проведение потенциала действия. Обязательным условием проведения нервного импульса является наличие на всем протяжении или в ограниченных, но повторяющихся участках волокна потенциалзависимых ионных каналов, ответственных за формирование ПД. В распространении ПД можно выделить два этапа: этап электротонического проведения, обусловленный физическими свойствами нервного волокна, и этап генерации ПД в новом участке на пути его движения, обусловленный реакцией ионных каналов. В зависимости от расположения и концентрации ионных каналов в мембране волокна возможно два типа проведения ПД: непрерывный и сальтаторный (скачкообразный). 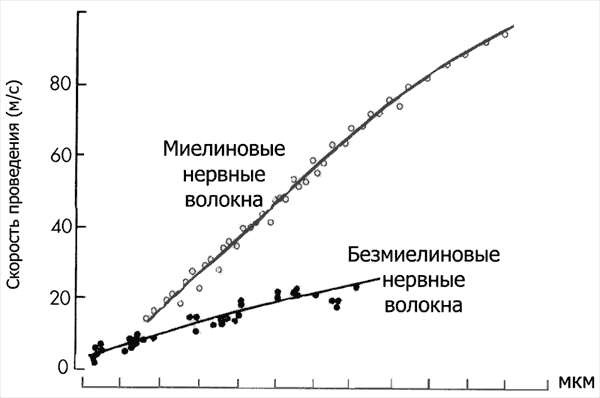 Рис. 3. Скорость проведения возбуждения в миелиновых и безмиелиновых нервных волокн. Непрерывное распространение ПД осуществляется в безмиелиновых волокнах типа С, имеющих равномерное распределение потенциалзависимых ионных каналов, участвующих в генерации ПД. Проведение нервного импульса начинается с этапа электротонического распространения возникшего ПД. Амплитуда ПД нервного волокна (мембранный потенциал + инверсия) составляет около 90 мВ, постоянная длины мембраны (λm) в безмиелиновых волокнах равна 0,1—1,0 мм. Поэтому ПД, распространяясь на этом расстоянии как электротонический потенциал и сохранив как минимум 37 % своей амплитуды, способен деполяризовать мембрану до критического уровня и генерировать на всем протяжении новые ПД (рис.5). При этом на этапе электротонического распространения нервного импульса ионы движутся вдоль волокна между деполяризованным и поляризованным участками, обеспечивая проведение возбуждения в соседние участки волокна. Реально при неповрежденном нервном волокне этап чисто электротонического распространения ПД (вдоль мембраны) предельно мал, так как потенциалзависимые каналы имеются в непосредственной близости друг от друга и, естественно, — от возникшего потенциала действия и наблюдается только до достижения деполяризации, равной 50 % Екр. Далее включается перемещение ионов в клетку (нервное волокно) и из клетки за счет активации ионных каналов. При формировании нового ПД в соседнем участке в фазе деполяризации возникает мощный ток ионов натрия в клетку вследствие активации натриевых каналов, приводящий к регенеративной (самоусиливающейся) деполяризации. Этот ток обеспечивает формирование нового ПД той же амплитуды, представляющий собой, как обычно, сумму двух величин — мембранного потенциала покоя и инверсии. В связи с этим проведение ПД осуществляется без декремента (без снижения амплитуды). Таким образом, непрерывное распространение нервного импульса идет через генерацию новых ПД по эстафете, когда каждый участок мембраны выступает сначала как раздражаемый (при поступлении к нему электротонического потенциала), а затем как раздражающий (после формирования в нем нового ПД). Сальтаторный ( прерывистый, скачкообразный ) тип проведения нервного импульса осуществляется в миелиновых волокнах (типы А и В), для которых характерна концентрация потенциалзависимых ионных каналов только в небольших участках мембраны (в перехватах Ранвье), где их плотность достигает 12 000 на 1 мкм2, что примерно в 100 раз выше, чем в мембранах безмиелиновых волокон. В области миелиновых муфт (межузловых сегментов), обладающих хорошими изолирующими свойствами, потенциалзависимых каналов почти нет, и мембрана осевого цилиндра там практически невозбудима. В этих условиях ПД, возникший в одном перехвате Ранвье, электротонически (вдоль волокна, без участия ионных каналов) распространяется до соседнего перехвата, деполяризуя там мембрану до критического уровня, что приводит к возникновению нового ПД, т.е. возбуждение проводится скачкообразно (рис. 6). Постоянная длина мембраны миелинового волокна достигает 5 мм. Это значит, что ПД, распространяясь электротонически на этом расстоянии, сохраняет 37 % своей амплитуды (около 30 мВ) и может деполяризовать мембрану до критического уровня (пороговый потенциал в перехватах Ранвье равен около 15 мВ). Поэтому в случае повреждения ближайших на пути следования перехватов Ранвье потенциал действия может электротонически возбудить 2—4-й и даже 5-й перехваты. Сальтаторное проведение ПД по миелиновым волокнам является эволюционно более поздним механизмом, возникшим впервые у позвоночных. Оно имеет два важных преимущества по сравнению с непрерывным проведением возбуждения. Во-первых, оно более экономично в энергетическом плане, т.к. возбуждаются только перехваты Ранвье, площадь которых менее 1 % мембраны, и, следовательно, надо меньше энергии для восстановления трансмембранных градиентов Na+ и К+, уменьшающихся в процессе формирования ПД. Во-вторых, возбуждение проводится с большей скоростью (см. табл.3), чем в безмиеликовых волокнах, так как возникший ПД на протяжении миелиновых муфт распространяется электротонически, что в 107 раз быстрее, чем скорость непрерывного проведения ПД в безмиелиновом волокне. 5. СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА ПРОВЕДЕНИЯ НЕРВНОГО ИМПУЛЬСА ПО МИЕЛИНОВЫМ И БЕЗМИЕЛИНОВЫМ ВОЛОКНАМ Механизм распространения возбуждения у различных нервных волокон неодинаков. Механизмы проведения возбуждения в безмиелиновых волокнах. При действии раздражителя пороговой силы на мембрану безмиелинового волокна изменяется ее проницаемость для ионов Nа+, которые мощным потоком устремляются внутрь волокна. В этом месте изменяется заряд мембраны (внутренняя становиться заряженной положительно, а наружная отрицательно). Это ведет к возникновению круговых токов (заряженных частиц) от ʼʼ+ʼʼ к ʼʼ–ʼʼ на протяжении всего волокна. Особенности распространения возбуждения по безмиелиновым волокнам: 1. Возбуждение распространяется непрерывно и всœе волокно сразу охватывается возбуждением. 2. Возбуждение распространяется с небольшой скоростью. 3. Возбуждение распространяется с декриментом (уменьшение силы тока к концу нервного волокна). По безмиелиновым волокнам возбуждение проводится к внутренним органам от нервных центров. При этом низкая скорость распространения возбуждения и его затухание не всегда выгодно организму. По этой причине природой был выработан еще один дополнительный механизм распространения возбуждения. Механизмы проведения возбуждения в миелиновых волокнах. Наличие у миелиновых волокон оболочки, обладающей высоким электрическим сопротивлением, а также участков волокна, лишенных оболочки - перехватов Ранвье создают условия для качественно нового типа проведения возбуждения по миелиновым нервным волокнам. В миелинизированном волокне токи проводятся только в зонах, не покрытых миелином (перехватах Ранвье). В этих участках генерируется очередной ПД. Перехваты длиной 1 мкм расположены через 1000 - 2000 мкм, характеризуются высокой плотностью ионных каналов, высокой электропроводностью и низким сопротивлением. При действии раздражителя пороговой силы на мембрану миелинового волокна в области перехвата Ранвье изменяется проницаемость для ионов Nа+, которые мощным потоком устремляются внутрь волокна. В этом месте изменяется заряд мембраны, что ведет к возникновению круговых токов. Этот ток идет через межтканевую жидкость к сосœеднему перехвату, где происходит смена заряда. Τᴀᴋᴎᴍ ᴏϬᴩᴀᴈᴏᴍ, возбуждение перепрыгивает с одного участка на другой. Обратное движение возбуждения невозможно так как участок, через который оно прошло, находится в фазе абсолютной рефрактерности. Особенности распространения возбуждения по миелиновым волокнам: 1. Распространение ПД в миелинизированных нервных волокнах осуществляется сальтаторно - скачкообразно от перехвата к перехвату, ᴛ.ᴇ. возбуждение (ПД) как бы «перепрыгивает» через участки нервного волокна, покрытые миелином, от одного перехвата к другому и все волокно сразу не охватывается возбуждением. 2. Возбуждение распространяется с большой скоростью. 3. Возбуждение распространяется без декримента. По миелиновым волокнам возбуждение распространяется от анализаторов к ЦНС, к скелетным мышцам, ᴛ.ᴇ. там, где требуется высокая скорость ответной реакции. 6. ЗАКОНЫ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ ПО НЕРВНЫМ ВОЛОКНАМ Закон двустороннего проведения возбуждения. Прямые доказательства этой закономерности были получены во второй половине XIX в. А.И.Бабухиным и Э.Дюбуа-Реймоном. Если стимул действует на средний участок изолированного нерва (Дюбуа-Реймон), то распространение возбуждения регистрируется как в проксимальном, так и в дистальном участках нерва. В опытах на электрическом органе у рыб, иннервируемом разветвлениями аксона одного нейрона, было показано (А.И.Бабухин), что при раздражении перерезанной веточки аксона возбуждение распространяется в необычном центростремительном направлении, передается на другие разветвления аксона, по которым идет в центробежном направлении (так называемый аксон-рефлекс). В условиях организма двустороннее проведение показано в аксонном холмике; возникший в этом месте ПД распространяется не только в аксон, но и в тело нейрона. На уровне целого организма аксоны нервных клеток проводят возбуждение только в одном направлении: от рецепторного отдела рефлекторной дуги к исполнительному органу (эффектору). Роль выпрямителя в рефлекторной дуге выполняют химические синапсы. − Закон изолированного проведения возбуждения. В обычных условиях деятельности нервного ствола (возбуждение только части нервных волокон, асинхронное распространение в них ПД) проведение возбуждения в составляющих его волокнах происходит практически изолированно. Это обусловлено тем, что петли тока в межклеточной жидкости ствола, имеющей низкое сопротивление, почти не проникают в невозбужденные волокна нерва из-за большого сопротивления их оболочек. Изолированное проведение импульсов по нервным волокнам обеспечивает точное афферентное и эфферентное влияния функционально разнородных волокон нерва. Однако при одновременном раздражении значительного количества волокон в межклеточной жидкости ствола возникает достаточно сильный внешний ток, способный возбудить неактивные (прежде всего высоковозбудимые) волокна и таким образом увеличить количество функционирующих нервных волокон в нерве, его эфферентное или афферентное влияние. − Закон физиологической непрерывности нерва.Обязательным условием проведения возбуждения по нервному волокну является анатомическая и функциональная целость возбудимой мембраны осевого цилиндра. Поэтому не только перерезка нерва, но и любое воздействие, нарушающее целость мембраны осевого цилиндра, например перевязка нерва, чрезмерное натяжение нервных волокон, создают непроводимость. Возможность функционального блока проведения возбуждения возможна при морфологической целостности волокон.Непроводимость наступает при воздействиях, нарушающих генерацию нервного импульса. Так, чрезмерное охлаждение или согревание, прекращение кровоснабжения, различные химические агенты, в частности местные обезболивающие — новокаин, кокаин, дикаин, прекращают проведение по нерву. Н.Е.Введенский (1901) показал, что при действии различных факторов на нерв (кокаина, хлороформа, фенола, хлористого калия, сильного фарадического тока) в нем сначала возникает трансформация ритма проводимого возбуждения (блокируется проведение высокочастотных потенциалов действия, и проводятся только низкочастотные ПД), а в дальнейшем может возникать полный блок проведения нервных импульсов — участок парабиоза. В этом участке возникает длительная деполяризация мембраны волокон, которая в результате закрытия инактивационных h-ворот в натриевых каналах сначала затрудняет генерацию ПД (уменьшается его амплитуда, увеличивается длительность, затягивается фаза абсолютной рефрактерности), а в дальнейшем, если инактивация натриевых каналов превысит 50 %, приводит к полной невозбудимости этого участка нервного волокна. Для возникновения блока в проведении возбуждения протяженность парабиотического участка должна превысить постоянную длину мембраны (λm), иначе ПД может распространиться через этот участок электротонически. Нарушение физиологической непрерывности нервных волокон возникает при действии анестетиков, электрического тока, при гипоксии, воспалении, охлаждении. После прекращения действия этих факторов проведение возбуждения по волокнам нерва восстанавливается. Однако, при углублении и усилении действия вызвавшего парабиоз агента обратимые изменения могут переходить в необратимое нарушение жизнедеятельности — смерть. 7. ОСОБЕННОСТИ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ В НЕРВНЫХ ВОЛОКНАХ − Большая скорость проведения возбуждения. Скорость проведения ПД в различных типах волокон нерва равна 0,5—120 м/с. Она значительно выше в миелиновых волокнах в связи с сальтаторным типом проведения ПД, а среди миелиновых волокон прямо пропорциональна диаметру волокна. Скорость проведения возбуждения в миелиновых нервных волокнах значительно выше, чем в других удлиненных возбудимых структурах, — в гладких миоцитах (0,02—0,10 м/с), рабочих кардиомиоцитах (около 1 м/с), и только в миоцитах проводящей системы сердца и скелетных миоцитах скорость проведения ПД (2—5 м/с) достигает величин распространения ПД в низкоскоростных нервных волокнах (тип С и В). Передача возбуждения по нервным волокнам является наиболее скоростным из известных способов передачи информации на значительные расстояния в организме. Для сравнения отметим, что скорость передачи гуморальных влияний ограничена скоростью кровотока, которая равна от 0,5 мм/с в капиллярах до 0,25 м/с в аорте (средняя скорость). − Малая утомляемость нервного волокна. При нормальном кровоснабжении (доставке кислорода и питательных веществ) проводящий возбуждение нерв практически неутомляемость. «Изумительно долгая неутомляемость нерва» впервые была показана Н.Е.Введенским (1883): в его опытах нерв сохранял способность к проведению возбуждения в течение 6—8 ч непрерывного раздражения несильными токами в условиях наличия кислорода в окружающей среде и поддержания влажного состояния нерва. Это обусловлено тем, что при проведении ПД по нервным волокнам используется всего лишь одна миллионная часть запасов трансмембранных ионных градиентов и, следовательно, нужны небольшие количества АТФ для восстановления (например, посредством Nа/К-насоса) ионных градиентов. Об энергетической экономности проведения возбуждения свидетельствует и низкая величина теплопродукции в работающем нерве, отражающая степень окислительного фосфорилирования в митохондриях. Ее величина в нерве (0,06 кал/г ткани в течение 1 ч) примерно в 16 раз меньше, чем на соответствующую единицу массы в целом организме в условиях основного обмена, и в миллион раз меньше, чем в работающей мышце. ЗАКЛЮЧЕНИЕ Исходя из вышесказанного, понятно что нервная ткань — ткань эктодермального происхождения, представляющая собой систему специализированных структур, образующих основу нервной системы и создающих условия для реализации её функций. Нервная ткань осуществляет связь организма с окружающей средой, восприятие и преобразование раздражителей в нервный импульс и передачу его к эффектору. Нервная ткань обеспечивает взаимодействие тканей, органов и систем организма и их регуляцию. Механизм распространения возбуждения у различных нервных волокон неодинаков. СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ 1. Альбертс Б., Брей Д. Молекулярная биология клетки М.: Мир, 2004. Т.3. С. 287-374. 2. Антипчук Ю.П. Гистология с основами эмбриологии М.: Просвещение, 2008. 240 с. 3. Березов Т.Т., Коровкин Б.Ф. Биологическая химия М.: Медицина, 2008. С.625-645. 4. Елисеев В.Г., Афанасьев Ю.И. Гистология М.: Медицина, 2002. С.224-249. 5. Заварзин А.А., Щелкунов С.И. Руководство по гистологии Л.: МедГИз, 2009. С.295-314. 6. Корочкин Л.И., Михайлов А.Т. Введение в нейрогенетику М.: Наука, 2000. С.7-98. 7. Николлс Дж.Г., Мартин А.Т. От нейрона к мозгу М.: УРСС, 2003. С.31-33, 143-163, 522-601. 8. Рябов К.П. Гистология с основами эмбриологии Минск: Высшая школа, 2000.255 с. 9. Хухо Ф. Нейрохимия М.: Мир, 2011. С.91-107, 187-237. 10. Ченцов Ю.С. Введение в клеточную биологию М.: Наука, 2005.443 с. 11. Шульговский В.В. Основы нейрофизиологии М.: Аспект пресс, 2000. С.14-35. |