МУ СРСП. Срсп 1 Новые направления цитологии, гистологии и физиологии растений Цель
 Скачать 1.01 Mb. Скачать 1.01 Mb.
|

|
Тема: «Рост, дифференцировка, морфогенез. Соотношение процессов роста и морфогенеза» Цель: Изучить механизмы роста, дифференцировки, морфогенеза. Установить соотношение процессов роста и морфогенеза. Процесс роста складывается из роста клеток, тканей и органов и происходит благодаря деятельности специальных образовательных тканей — меристем, где клетки активно делятся, проходят стадии растяжения и дифференциации. Растение как целостный организм обладает способностью к неограниченному росту (за исключением таких органов, как листья, цветки, плоды). Рост растений неразрывно связан с процессами развития и обусловлен особенностями прохождения растениями различных фаз онтогенеза. Переход к репродуктивной стадии обычно сопровождается усилением процессов старения и постепенным ослаблением активности меристем; поэтому многолетние и позднеспелые растения достигают больших размеров, чем скороспелые. Образование плодов и семян снижает интенсивность роста деревьев (в годы обильных урожаев годичные кольца имеют минимальную ширину), что связано, очевидно, с расходом питательных веществ на репродуктивные органы, а также, возможно, с действием ингибирующих веществ, вырабатываемых созревающими плодами и семенами. У однолетних растений плодоношение сопровождается прекращением роста и их отмиранием. Важная особенность роста растений — его ритмичность — чередование процессов интенсивного и замедленного роста. Существуют ритмы, следующие за изменениями во внешней среде, и ритмы, контролируемые внутренними факторами. В северных широтах процессы роста растений обычно прерываются продолжительными периодами торможения, индуцируемыми укорочением длины дня в конце лета и осенью и понижением температуры. Такое отсутствие видимого роста называется покоем растений. В регуляции роста растений большое значение имеет внутренняя корреляции — взаимовлияния органов. К ним относится явление апикального доминирования, т. е. торможение роста боковых побегов и почек верхушечной почкой. Степень апикального доминирования часто определяет характер роста растений. Так, у большинства хвойных и некоторых лиственных деревьев ствол растёт намного быстрее, чем боковые ветви. В результате этого деревья имеют более или менее конусовидную форму кроны. Апикальное доминирование усиливается при боковом затенении и ослабляется при хорошем освещении. Это можно наблюдать на деревьях, растущих в лесу и на открытом месте. Рост растений регулируется также фитогормонами, стимулирующими (ауксины, гиббереллины и цитокинины) или ингибирующими (абсцизовая кислота) его процессы. Фитогормоны вырабатываются в зонах роста — в верхушечных меристемах побегов, корней, растущих листьях, плодах. Интенсивность ростовых процессов находится в прямой зависимости от снабжения меристем продуктами фотосинтеза — ассимилятами, а, следовательно, и от условий освещения, минерального питания, водного режима, длины дня, температуры. У растений продолжительность жизни различная. Некоторые растения живут всего несколько недель (например, эфемеры), другие могут существовать сотни и тысячи лет. У некоторых растений цветение бывает всего 1 раз в жизни, после чего наступает период плодоношения, затем растение отмирает. Растения, цветущие и плодоносящие 1 раз в жизни, называют монокарпическими. К этой группе относятся все однолетние травянистые растения, жизненный цикл которых с периодом цветения и плодоношения заканчивается в течение 1 вегетационного периода, т.е. с весны до осени. У поликарпических растений период плодоношения продолжается очень долго и занимает большую часть жизни. Поликарпическими растениями являются плодовые деревья (яблоня). Они зацветают спустя несколько лет после посадки, затем цветут и плодоносят в течение многих лет жизни. Вегетативное развитие растений включает последовательное появление листьев на побеге, а также изменение их размеров и формы (у деревьев — кроны, со всё усложняющейся системой побегов) в первой половине жизни и постепенное отмирание в результате старения — во второй. Наиболее кардинальные изменения в растении начинаются при переходе от вегетативного к генеративному развитию, т. е. цветению и плодоношению. Монокарпические растения требуют для перехода к цветению индуцирующего воздействия сначала низких положительных (2—10° С) температур (яровизация), а затем ритмичной смены света и темноты определенной длительности (фотопериодическая индукция). В результате в растениях вырабатываются гормоны (гиббереллины, цитокинины, этилен и др.), вызывающие изменения в точке роста в направлении репродуктивного развития. Без специфичных условий переход к репродуктивному развитию не происходит, в чём проявляется одна из характерных черт развития растений — его многовариантность. В ходе развития растений выделяют фазы: молодость, или ювенильная фаза (относительно медленный рост, цветение отсутствует); фаза усиленного роста; зрелость (вегетативный рост отсутствует, растение готово к цветению и плодоношению); старость (постепенное отмирание вегетативных органов). Один из внутренних факторов, регулирующих развитие растений, — коррелятивное взаимовлияние органов. Например, верхушечная почка тормозит или полностью подавляет рост боковых почек; удаление её может привести к заложению репродуктивных органов. Это взаимовлияние осуществляется как фитогормонами, так и через обмен метаболитов. Развитие растений тесно связано с ростом, причём взаимоотношения между этими процессами могут изменяться в течение онтогенеза от синергизма до антагонизма. Контрольные вопросы: 1. Опишите рост растений. 2. Опишите морфогенез растений. 3. Как происходит дифференцировка? 4. Опишите растения, цветущие и плодоносящие 1 раз в жизни 5. Опишите растения, которые цветут и плодоносят в течение многих лет жизни. Задания для СРС: 1. Составьте схему роста и развития растений. 2. Проанализируйте соотношение процессов роста, морфогенеза и развития. СРСП 9 Тема: «Онтогенез растительной клетки: деление клеток (митоз и амитоз)» Цель: Изучить разные способы деления клетки. Индивидуальное развитие растительного организма, которое начинается с деления оплодотворенной яйцеклетки и заканчивается образованием зиготы, называется онтогенезом. Под онтогенезом семенных растений понимают период с момента прорастания семени до образования новых, во время которого происходит рост и развитие организма. 1. МИТОЗ - этот способ наиболее распространен. Так делятся клетки в конусах нарастания стеблей и на кончиках корней. При делении ядра путем митоза из одной материнской клетки образуются 2 дочерние, которые приобретают такой же набор хромосом, какой был в материнской клетке до деления. Происходит точное распределение носителей наследственной информации - молекул ДНК - между образовавшимися клетками. В дочерних клетках обеспечивается наследственное сходство с исходной материнской клеткой. Деление путем митоза является непрерывным процессом, который условно делится на 4 фазы (рисунок 9.1): 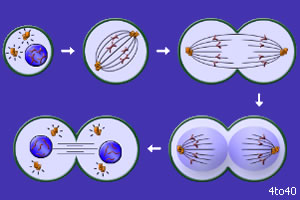 Рисунок 9.1 – Митоз Профаза - ядро увеличивается в объеме, в нем постепенно начинают выявляться хромосомы. К концу профазы исчезает ядрышко, ядерная оболочка растворяется, на противоположных полюсах клетки происходит формирование ахроматинового веретена - веретена деления. Оно представляет собой систему ахроматиновых (неокрашивающихся) нитей, которые тянутся от полюсов клетки. Метафаза - формирование веретена деления заканчивается. Хромосомы приобретают определенную форму, характерную для того или иного вида растений. Хромосомы обычно бывают двуплечими. В месте перегиба называемом центромерой, они соединяются с ахроматиновой нитью веретена. Хромосомы располагаются в экваториальной плоскости клетки. Хорошо заметно, что каждая хромосома состоит из 2 дочерних хроматид, которые постепенно отталкиваются, разъединяются и остаются соединенными лишь в центромерном участке. Анафаза - начинается с деления центромеры - образуются 2 сестринские центромеры. Хроматиды и сестринские центромеры направлены к противоположным полюсам клетки. Хроматиды приобретают самостоятельность. Они увлекаются к противоположным полюсам клетки. Каждая хроматида впоследствии становится самостоятельной хромосомой. Вторая нить строится путем редупликации. У каждого полюса клетки окажется столько же хромосом, сколько их было в материнской клетке. Телофаза - последняя фаза деления ядра, представляет собой фазу обратную профазе. Хромосомы становятся плохо заметными. Восстанавливаются оболочка ядра и ядрышко. Ахроматиновое веретено деления исчезает. В экваториальной плоскости формируется срединная пластинка (фрагмопласт). Фрагмопласт - это зачаток клеточной стенки. Сначала в центральной области веретена деления появляются образующиеся из мембран комплекса Гольджи многочисленные пузырьки, содержащие пектиновые вещества. В результате увеличения их числа и постоянного слияния друг с другом в направлении от центра к периферии клетки возникают длинные плоские мешочки - так называемые мембранные цистерны, которые, сливаясь с плазматической мембраной, делят материнскую клетку на 2 дочерние. Происходит формирование клеточных стенок. Начинается редупликация ДНК по принципу комплементарности азотистые основания ДНК Аденин - Тимин, Гуанин - Цитозин. В РНК вместо тимина - урацил. Таким образом, осуществляется деление клетки - цитокинез. Амитоз - прямое деление, наблюдается у некоторых видов бактерий и грибов, у высших растений лишь в старых и больных клетках. Осуществляется путем простой перетяжки ядра на 2 части с произвольным количеством ядерного вещества без образования веретена деления. В норме так делятся клетки клубней картофеля и эндосперм (рисунок 9.2). 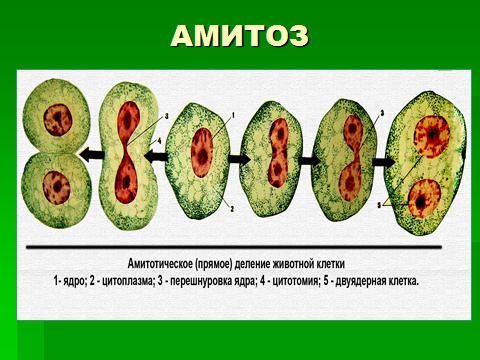 Рисунок 9.2 – Схема амитоза: 1- ядро, 2 – цитоплазма, 3 – перешнуровка ядра; 4 – цитотомия, 5 – двуядерная клетка Контрольные вопросы: Сущность митоза; Как осуществляется амитоз; Как называется зачаток клеточной стенки? Как он образуется? По какому принципу происходит редупликация ДНК? Его сущность; Опишите профазу; Опишите метафазу; Опишите анафазу; Опишите телофазу; Что такое цитокинез? Задания для СРС: Составьте схемы митоза и амитоза. Проанализируйте: в чем заключается значение этих видов деления для растений? Опишите, что произойдет, если будет отсутствовать митоз. СРСП 10 Тема: «Онтогенез растительной клетки: деление клеток (мейоз)» Цель: Изучить разные способы деления клетки. При половом размножении дочерний организм возникает в результате слияния двух половых клеток (гамет) и последующего развития из оплодотворенной яйцеклетки — зиготы. Половые клетки родителей обладают гаплоидным набором (n) хромосом, а в зиготе при объединении двух таких наборов число хромосом становится диплоидным (2n): каждая пара гомологичных хромосом содержит одну отцовскую и одну материнскую хромосому. Гаплоидные клетки образуются из диплоидных в результате особого клеточного деления — мейоза. Мейоз — разновидность митоза, в результате которого из диплоидных (2п) соматических клеток половых желез образуются гаплоидные гаметы (1n). При оплодотворении ядра гаметы сливаются, и восстанавливается диплоидный набор хромосом. Таким образом, мейоз обеспечивает сохранение постоянного для каждого вида набора хромосом и количества ДНК. Мейоз представляет собой непрерывный процесс, состоящий из двух последовательных делений, называемых мейозом I и мейозом II. В каждом делении различают профазу, метафазу, анафазу и телофазу. В результате мейоза I число хромосом уменьшается вдвое (редукционное деление): при мейозе II гаплоидность клеток сохраняется (эквационное деление). Клетки, вступающие в мейоз, содержат генетическую информацию 2n2хр (рис. 1). В профазе мейоза I происходит постепенная спирализация хроматина с образованием хромосом. Гомологичные хромосомы сближаются, образуя общую структуру, состоящую из двух хромосом (бивалент) и четырех хроматид (тетрада). Соприкосновение двух гомологичных хромосом по всей длине называется конъюгацией. Затем между гомологичными хромосомами появляются силы отталкивания, и хромосомы сначала разделяются в области центромер, оставаясь соединенными в области плеч, и образуют перекресты (хиазмы). Расхождение хроматид постепенно увеличивается, и перекресты смещаются к их концам. В процессе конъюгации между некоторыми хроматидами гомологичных хромосом может происходить обмен участками — кроссинговер, приводящий к перекомбинации генетического материала. К концу профазы растворяются ядерная оболочка и ядрышки, формируется ахроматиновое веретено деления. Содержание генетического материала остается прежним (2n2хр). В метафазе мейоза I биваленты хромосом располагаются в экваториальной плоскости клетки. В этот момент спирализация их достигает максимума. Содержание генетического материала не изменяется (2п2хр). В анафазе мейоза I гомологичные хромосомы, состоящие из двух хроматид, окончательно отходят друг от друга и расходятся к полюсам клетки. Следовательно, из каждой пары гомологичных хромосом в дочернюю клетку попадает только одна — число хромосом уменьшается вдвое (происходит редукция). Содержание генетического материала становится 1n2хр у каждого полюса. В телофазе происходит формирование ядер и разделение цитоплазмы — образуются две дочерние клетки. Дочерние клетки содержат гаплоидный набор хромосом, каждая хромосома — две хроматиды (1n2хр). Интеркинез — короткий промежуток между первым и вторым мейотическими делениями. В это время не происходит репликации ДНК, и две дочерние клетки быстро вступают в мейоз II, протекающий по типу митоза. 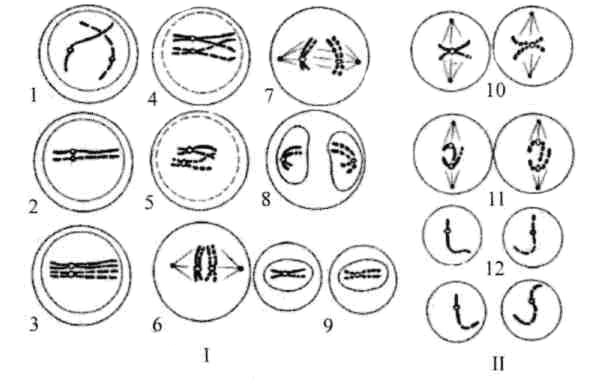 Рисунок 10.1 – Схема мейоза (показана одна пара гомологичных хромосом). Мейоз I: 1, 2, 3. 4. 5 — профаза; 6 —метафаза; 7 — анафаза; 8 — телофаза; 9 — интеркинез. Мейоз II; 10 —метафаза; 11 —анафаза; 12 — дочерние клетки. В профазе мейоза II происходят тс же процессы, что и в профазе митоза. В метафазе хромосомы располагаются в экваториальной плоскости. Изменений содержания генетического материала не происходит (1n2хр). В анафазе мейоза II хроматиды каждой хромосомы отходят к противоположным полюсам клетки, и содержание генетического метериала у каждого полюса становится lnlxp. В телофазе образуются 4 гаплоидные клетки (lnlxp). Таким образом, в результате мейоза из одной диплоидной материнской клетки образуются 4 клетки с гаплоидным набором хромосом. Кроме того, в профазе мейоза I происходит перекомбинация генетического материала (кроссинговер), а в анафазе I и II — случайное отхождение хромосом и хроматид к одному или другому полюсу. Эти процессы являются причиной комбинативной изменчивости. Биологическое значение мейоза: 1) является основным этапом гаметогенеза; 2) обеспечивает передачу генетической информации от организма к организму при половом размножении; 3) дочерние клетки генетически не идентичны материнской и между собой. А так же, биологическое значение мейоза заключается в том, что уменьшение числа хромосом необходимо при образовании половых клеток, поскольку при оплодотворении ядра гамет сливаются. Если бы указанной редукции не происходило, то в зиготе (следовательно, и во всех клетках дочернего организма) хромосом становилось бы вдвое больше. Однако это противоречит правилу постоянства числа хромосом. Благодаря мейозу половые клетки гаплоидны, а при оплодотворении в зиготе восстанавливается диплоидный набор хромосом (рисунок 10.2 и 10.3). 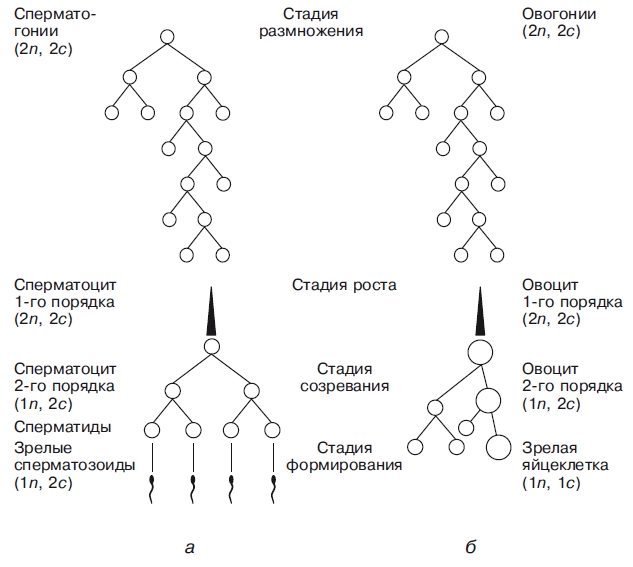 Рисунок 10.2 – Схема гаметогенеза: à — сперматогенез; б — овогенез 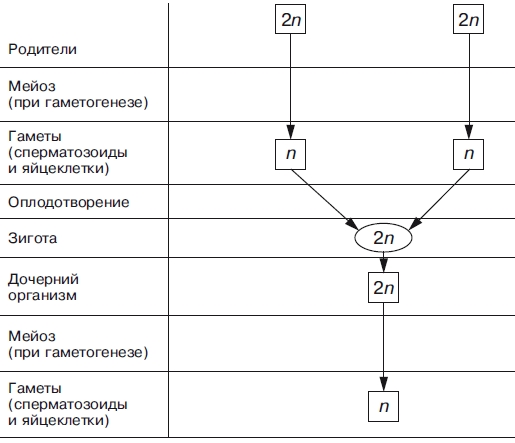 Рисунок 10.3 – Схема, иллюстрирующая механизм сохранения диплоидного набора хромосом при половом размножении Контрольные вопросы: Сущность мейоза; Опишите профазу мейоза 1; Опишите метофазу мейоза 1; Опишите анофазу мейоза 1; Опишите телофазу мейоза 1; Опишите мейоз 2; Биологическое значение мейоза; Задания для СРС: Составьте схему мейоза; Опишите, что произойдет, если будет отсутствовать мейоз. СРСП 11 Клеточный цикл Цель: Ознакомиться с общими и отличительными признаками между растительными и животными клетками Задание: Ответьте на вопросы: что такое интерфаза? Что происходит в пресинтетический период? Характеристика синтетического периода; Особенности постсинтетического периода. Фактор стимуляции митоза; Циклин, его особенности. Функция воспроизведения и передачи генетической информацииобеспечивается в ходе клеточного цикла. Клеточный цикл- совокупность явлений между двумя последовательными делениями клетки или между ее образованием и гибелью. Клеточный цикл включает собственно митотическое делениеи интер-фазу - промежуток между делениями (рисунок 1). Интерфаза Интерфаза значительно более длительна, чем митоз (обычно занимает не менее 90% всего времени клеточного цикла) и подразделяется на три периода: пресинтетический или постмитотический (Gj), синтетический (S) и постсинтетический или премитотический (G2). 1. Пресинтетический или постмитотический (Gj-) период (от англ. gap - промежуток) наступает сразу же после митотического деления клетки и характеризуется активным ростом клетки и синтезом белка и РНК, благодаря чему клетка достигает нормальных размеров и восстанавливает необходимый набор органелл. Gx-период длится от нескольких часов до нескольких дней. В течение этого периода синтезируются особые "запускающие" белки (trigger proteins), или активаторы S-nepuoda. Они обеспечивают достижение клеткой определенного порога (точки R - рестрикции или ограничения), после которого она вступает в S-период. Контроль, осуществляемый на уровне точки R (при переходе из Gi в S), ограничивает возможность нерегулируемого размножения клеток. Проходя эту точку, клетка переключается на последующую регуляцию внутренними факторами клеточного цикла, которая обеспечивает закономерное завершение ее деления. Если клетка не достигает точки R, она выходит из цикла и вступает в период репродуктивного покоя (G0) для того, чтобы (в зависимости от причин остановки): (1) дифференцироваться и выполнять свои специфической функции, (2) выжить в условиях недостаточности питательных веществ или факторов роста, (3) осуществить репарацию поврежденной ДНК. Клетки одних тканей при соответствующей стимуляции вновь способны возвращаться из периода (G0) в клеточный цикл, других - утрачивают эту способность по мере дифференцировки. 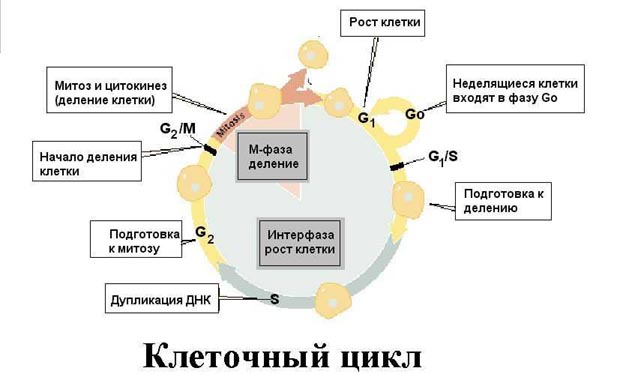 1 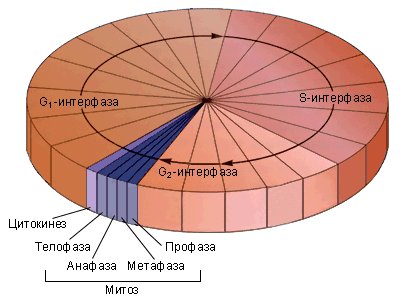 2 Рисунок 1- Клеточный цикл: 1 – схема; 2 - графическое соотношение фаз клеточного цикла. 2. Синтетический (S-) период характеризуется удвоением содержания (репликацией) ДНК и синтезом белков, в частности, гасто-нов, которые поступают в ядро из цитоплазмы и обеспечивают нуклео-сомную упаковку вновь синтезированной ДНК. В результате происходит удвоение числа хромосом. Одновременно удваивается число центрио-лей. S-период длится у большинства клеток 8-12 часов. 3. Постсинтетический или премитотический (G2-) период следует за S-периодом и продолжается вплоть до митоза (часто обозначаемого буквой М). В течение этого периода клетка осуществляет непосредственную подготовку к делению. Происходит созревание центриолей, запасается энергия, синтезируются РНК и белки (в частности, тубулин), необходимые для процесса деления. Длительность С2-периода составляет 2-4 часа. Возможность выхода клетки из С2-периода в Go-период с последующим возвращением в 02-период в настоящее время большинством авторов отрицается. Контроль вступления клетки в митоз осуществляется двумя специальными факторами с противоположно направленными эффектами: митоз тормозится до момента завершения репликации ДНК М-задерживающим фактором и индуцируется М-стимулирующим фактором. Действие последнего проявляется лишь в присутствии других белков - циклите (синтезируются на протяжении всего цикла и распадаются в середине митоза). Регуляция клеточного цикла Клеточный цикл представляет собой однонаправленный процесс, где клетка последовательно проходит разные его периоды, без их пропуска или возврата к предыдущим стадиям. Вступив в клеточный цикл, клетка его заканчивает синтезом ДНК и делением клетки. В цитоплазме митотической клетки есть фактор (или факторы), стимулирующие митоз (ФСМ или MPF – mitosis promoting factor). Этот фактор вызывает не только конденсацию хромосом, но и приводит к распаду ядерной оболочки, т.е. переводит интерфазную клетку, даже без синтеза ДНК, в митотическое состояние (конечно, дальше появления конденсированных хромосом развитие митоза не идет). В ооците на стадии метафазы II деления в цитоплазме существует фактор (или факторы), стимулирующие созревание яйцеклетки (ФСС или MPF – maturation promoting factor). Оказалось, что этот фактор (будем называть его MPF) присутствует в клетках только во время митотического состояния. Он обнаруживается также и во время дробления яйцеклетки. Таким образом, уровень MPF в интерфазных клетках низкий, а в митотических высокий. Далее было найдено, что при инъекции цитоплазмы из митотических клеток культуры ткани в нестимулированный ооцит X. laevis, происходит созревание ооцита. Следовательно, фактор, стимулирующий митоз и фактор, стимулирующий созревание ооцитов – одно и тоже. Этот фактор, MPF, был выделен и охарактеризован. Это гетеродимерный комплекс, состоящий из белка циклина и зависимой от циклина протеинкиназы (Cyclin dependent kinase – Сdk), фермента, относящегося к фосфорилазам, который модифицирует белки, перенося фосфатную группу от АТФ на аминокислоты серин и треонин. Следовательно, MPF состоит из двух субъединиц: каталитической (Сdk) и регуляторной (циклин). Циклин постоянно синтезируется в течение эмбрионального клеточного цикла и резко разрушается при вступлении в анафазу. СРСП №12 |