Практика к гистологии. Строение артерий
 Скачать 3.13 Mb. Скачать 3.13 Mb.
|

|
Сердечно-сосудистая система. С.Ю. Виноградов, С.В. Диндяев Строение артерийАртерии эластического типа за счет большого количества эластических волокон и мембран способны растягиваться при систоле сердца и возвращаться в исходное положение во время диастолы. В таких артериях кровь протекает под большим давлением (120-130 мм рт.ст.) и с большой скоростью (0,5-1,3 м/с). В качестве примера артерии эластического типа рассмотрим строение аорты.  Рис. 1. Артерия эластического типа – аорта кролика. Окраска орсеином. Объектив 4. Внутренняя оболочка аорты состоит из следующих элементов: эндотелий, подэндотелиальный слой, сплетение эластических волокон. Эндотелий состоит из крупных (иногда до 500 мкм в длину и 150 мкм в ширину) плоских одноядерных, реже многоядерных, полигональных клеток, расположенных на базальной мембране. В эндотелиальных клетках слабо развита эндоплазматическая сеть, но много митохондрий, микрофиламентов, пиноцитозных пузырьков. Подэндотелиальный слой развит хорошо (15-20 % от толщины стенки). Он образован рыхлой волокнистой неоформленной соединительной тканью, которая содержит тонкие коллагеновые и эластические волокна, много аморфного вещества и малодифференцированных клеток типа гладкомышечных фибробластов, макрофагов. Основное аморфное вещество подэндотелиального слоя, богатое гликозаминогликанами и фосфолипидами, играет большую роль в трофике стенки сосуда. Физико-химическое состояние этого вещества обусловливает степень проницаемости сосудистой стенки. С возрастом в нем накапливается холестерин и жирные кислоты. В этом слое отсутствуют собственные сосуды (vasa vasorum). Сплетение эластических волокон состоит из двух слоев: - внутренний циркулярный, - наружный продольный. Средняя оболочка аорты состоит из 40-50 эластических окончатых мембран, которые связаны между собой эластическими волокнами и образуют вместе с эластическими элементами других оболочек единый эластический каркас. Между мембранами располагаются гладкие миоциты, фибробласты, сосуды сосудов, нервные элементы. Большое количество эластических элементов в стенке аорты смягчает толчки крови, выбрасываемой в сосуд во время сокращения левого желудочка сердца, и обеспечивает поддержание тонуса сосудистой стенки во время диастолы. Наружная оболочка аорты образована рыхлой волокнистой соединительной тканью с большим количеством толстых коллагеновых и эластических волокон, располагающихся в основном в продольном направлении. В этой оболочке также имеются питающие сосуды, нервные элементы и жировые клетки. Артерии мышечного типа Внутренняя оболочка содержит эндотелий с базальной мембраной, подэндотелиальный слой, состоящий из тонких эластических и коллагеновых волокон и малоспециализированных клеток, внутреннюю эластическую мембрану, представляющую собой агрегированные эластические волокна. Иногда мембрана может быть двойной. Средняя оболочка состоит преимущественно из гладких миоцитов, расположенных по пологой спирали. Между ними располагаются соединительнотканные клетки типа фибробластов, коллагеновые и эластические волокна. Спиральное расположение гладких миоцитов обеспечивает при их сокращении уменьшение объема сосуда и проталкивание крови в дистальные отделы. Эластические волокна на границе с внутренней и наружной оболочками сливаются с их эластическими элементами. За счет этого создается единый эластический каркас сосуда, обеспечивающий эластичность при растяжении и упругость при сдавлении, препятствует спадению артерий. На границе средней и наружной оболочек может формироваться наружная эластическая мембрана. Наружная оболочка образована рыхлой волокнистой неоформленной соединительной тканью, в которой волокна располагаются косо и продольно. Необходимо отметить, что по мере уменьшения диаметра артерий толщина всех оболочек уменьшается. Истончаются подэндотелиальный слой и внутренняя эластическая мембрана внутренней оболочки, уменьшается количество гладких миоцитов и эластических волокон в средней, исчезает наружная эластическая мембрана.  Рис. 2. Артерии мышечного (А) и эластического (Б) типов: 1 – внутренняя оболочка (а – эндотелий), 2 – средняя оболочка, 3 – наружная оболочка.  Рис. 3. Артерия (1) и вена (2) мышечного типа кролика. Окраска: гематоксилин-эозин; объектив 10.  Микроциркуляторное руслоМикроциркуляторное русло (МЦР) – это система мелких сосудов, которая обеспечивает регуляцию кровенаполнения органов, транскапиллярный обмен и дренажно-депонирующую функцию. Состав МЦР: артериолы, в т.ч. конечные артериолы (диаметр 50-100 мкм), прекапилляры (диаметр 14-16 мкм), гемокапилляры (кровеносные капилляры) (диаметр 3-40 мкм), посткапилляры (диаметр 8-30 мкм), венулы (диаметр от 30 до 100 мкм), 6) артериоловенулярные анастомозы, 7) лимфатические капилляры. Артериолы – это наиболее мелкие артериальные сосуды мышечного типа, выполняющие следующие функции: транспорт артериальной крови в МЦР, перераспределение крови в МЦР, 3) регуляция кровенаполнения МЦР, 4) регуляция артериального давления. В артериолах сохраняются три оболочки, но выражены они очень слабо. Внутренняя оболочка содержит эндотелий с базальной мембраной, тонкий подэндотелиальный слой и тонкую внутреннюю эластическую мембрану. В базальной мембране эндотелия и во внутренней эластической мембране артериол встречаются перфорации, обеспечивающие транспорт из крови к гладким миоцитам нейромедиаторов, гормонов и др. биологически активных веществ. Средняя оболочка состоит из 1-2-х слоев спирально направленных гладких миоцитов и небольшого количества эластических и коллагеновых волокон. Гладкие миоциты обязательно присутствуют в месте отхождения от артериол прекапилляров. Наружная оболочка тонкая и представлена рыхлой волокнистой неоформленной соединительной тканью. Таким образом, для артериол характерны следующие структурные особенности: мощная мышечная оболочка, толщина стенки превалирует над диаметром просвета → способность к спазмированию, обилие клеточных рецепторов на эндотелии, перфорированная базальная мембрана, тесный контакт эндотелиоцитов и гладких миоцитов. Прекапилляры выполняют следующие функции: транспорт артериальной крови в капилляры ритмичное сокращение сфинктеров регулирует кровенаполнение отдельных групп гемокапилляров Структурные особенности прекапилляров: стенка теряет оболочечный тип строения стенка резко истончается гладкие миоциты расположены поодиночке сфинктеры в местах отхождения прекапилляров от артериол появляются одиночные перициты Кровеносные капилляры Гемокапилляры – наиболее многочисленные (около 40 миллиардов) и тонкие сосуды. Для них характерны следующие основные функции: обмен веществ между кровью и тканями (в т. ч. газовый обмен), транспортировка крови, барьерная (участие в создании гистогематических барьеров), депонирование крови, защитная (участие в воспалительных и иммунных реакциях), трансмуральная миграция лейкоцитов в РВСТ (трансмуральный - это относительное прилагательное, означающее - проходящий и/или действующий через стенку полого органа), транссудация плазмы ((transsudatio; транс- + лат. sudo, sudatum потеть, сочиться) выход жидкой части крови из капилляров и венул в тканевые пространства или полости тела) Строение гемокапилляров В стенке гемокапилляров имеется три слоя (как аналоги трех оболочек рассмотренных ранее сосудов): внутренний слой – представлен эндотелием с базальной мембраной, поверхность эндотелиоцитов, обращенная к току крови, покрыта слоем гликопротеидов (параплазмолеммальный слой); средний слой – содержит перициты, лежащие дискретно (т.е. в определенных участках) в расщеплениях базальной мембраны и являющиеся камбиальными клетками; наружный слой – состоит из адвентициальных клеток, тонких коллагеновых или ретикулярных волокон, аморфного вещества. Классификации гемокапилляров Классификация капилляров по диаметру: узкие – диаметр меньше 7 мкм (находятся в легких, нервах, поперечнополосатых мышцах и др.), средние – диаметром от 7 до 10-11 мкм (характерны для кожи и слизистых оболочек), широкие – диаметр 10-30 мкм (встречаются в некоторых эндокринных органах, печени, кроветворных органах), гигантские – диаметр более 30 мкм. Классификация капилляров по строению: соматический тип (с непрерывным эндотелием и непрерывной базальной мембраной) Локализация: скелетные мышцы, мозг, легкие и др. фенестрированный тип (с фенестрами в эндотелии и непрерывной базальной мембраной) Локализация: эндокринные органы, почки порозный тип (со сквозными отверстиями в эндотелии и базальной мембране) Локализация: печень, кроветворные органы  Рис. 4. Артериолы (1), венулы (2) и Рис. 5. Гемокапилляр: 1 – перицит; 2 – капилляры (3) кролика. Окраска: эндотелиоцит; 3 – адвентициальная клетка. гематоксилин-эозин; объектив 40. Пути трансэндотелиального транспорта капилляров: 1) пассивный транспорт, активный транспорт (пиноцитоз, фагоцитоз), везикулярный транспорт, 4) фенестры, 5) поры. Гистогематический барьер: эндотелиоцит, базальная мембрана, периэндотелильное пространство (перициты, адвентициальные клетки), рабочая клетка. Резервные капилляры – представляют собой плазмолеммальные капилляры, заполненные плазмой. Посткапилляры выполняют функции: отведение венозной крови гематотканевой обмен депонирование крови облегченная миграция лейкоцитов в РВСТ Строение стенки идентично строению стенки гемокапилляра, но имеются некоторые особенности: эндотелий часто фенестрированный появляются отдельные гладкие миоциты Венулы - строение их стенки идентично строению стенки безмышечных и маломышечных вен. Их внутренняя оболочка состоит из эндотелия с базальной мембраной и перицитов в расщеплениях базальной мембраны. Средняя оболочка содержит гладкие миоциты, количество которых увеличивается по мере увеличения диаметра венул (в мышечных венулах они образуют уже 1-2 слоя), тонкие коллагеновые и эластические волокна. Наружная оболочка образована рыхлой волокнистой соединительной тканью. Функции: отведение венозной крови гематотканевой обмен депонирование крови облегченная миграция лейкоцитов в РВСТ Артериоловенулярные анастомозы (АВА) имеются почти во всех органах и обеспечивают соединение артериального русла непосредственно с венозным в обход капилляров. Этим обеспечиваются: перераспределение крови внутри органов, шунтирование крови Классификация: 1) истинные АВА (шунты) – по ним в венозную систему сбрасывается чистая артериальная кровь; подразделяются на две подгруппы: простые АВА – в них регуляция кровотока осуществляется гладкими миоцитами средней оболочки артериолы; АВА со специальными сократительными структурами в виде валиков или подушек в подэндотелиальном слое, образованными гладкими миоцитами. К этой же группе относятся АВА эпителиоидного типа (простые и сложные). В средней оболочке простых АВА имеются овальные светлые клетки (Е-клетки), похожие на эпителиоциты и способные к набуханию, тем самым регулируя просвет сосуда. Сложные, или клубочковые, АВА характеризуются тем, что приносящая артериола делится на 2-4 ветви, которые переходят в венозный сегмент. В стенке могут быть эпителиоподобные клетки. 2) атипичные АВА (полушунты) – по ним течет смешанная кровь, т.к. представлены коротким гемокапилляром. СЕРДЦЕЭмбриональные источники развития и их производныеПарные мезенхимальные трубки → эндокард Висцеральный листок мезодермы → миоэпикардиальные пластинки → миокард и эпикард Париетальный листок мезодермы → перикард Мезенхима → коронарные сосуды Нейроэктодерма → ганглиозная пластинка → нервный аппарат Сердце закладывается в начале 3-й недели. Между энтодермой и висцеральным листком спланхнотома из мезенхимы образуются две трубки, выстланные эндотелием. Эти трубки – зачаток эндокарда. Трубки растут и окружаются висцеральным листком спланхнотома. Эти участки спланхнотома утолщаются и дают начало миоэпикардиальным пластинкам. Во время смыкания кишечной трубки происходит сближение и срастание обеих закладок сердца. Теперь зачаток сердца имеет вид двухслойной трубки: эндокардиальный слой и миоэпикардиальный. Позднее из миоэпикардиальной пластинки дифференцируются две части: внутренняя, которая прилежит к мезенхимной трубке, превращается в зачаток миокарда, а наружная – в эпикард. Основные функции сердца: сократительная гемодинамическая эндокринная (секреция предсердного натрий-уретического фактора) генерация потенциала действия создание околосердечного электромагнитного поля Строение стенки сердца Оболочки стенки и их структурный состав: 1. Эндокардэндотелиальный слой (эндотелий на базальной мембране) подэндотелиальный слой (РВСТ) мышечно-эластический слой (гладкие миоциты, РВСТ) наружный соединительнотканный слой (РВСТ, могут быть одиночные кровеносные сосуды) Дубликатурой эндокарда являются клапаны (створчатые и полулунные) 2. Миокард (сократительные, секреторные и проводящие кардиомиоциты, РВСТ, жировая ткань, сосуды, нервный аппарат) 3-4. Эпикард и перикард (имеют сходный структурный состав: мезотелий, РВСТ, жировая ткань, сосуды, нервный аппарат. Между ними находится перикардиальная полость В сердце имеется также фиброзный каркас (фиброзные кольца, треугольники, мембраны). Эти структуры построены из ПВСТ (может встречаться хрящевая ткань).  Рис. 6. Стенка сердца быка: 1 – проводящие кардиомиоциты (волокна Пуркинье) Окраска: гематоксилин-эозин; объектив 10. Проводящая система сердца (ПСС)Общая функция – обеспечение автоматического режима сокращения миокарда по принципу «иерархии соподчинения» Типы кардиомиоцитов ПСС и их функциональное назначениеР-клетки (пейсмекеры, водители ритма первого порядка) → генерация потенциала действия (ПД) с частотой 60-90 имп/мин. Переходные (водители ритма второго порядка) → генерация ПД с частотой 40-50 имп/мин Проводящие (собственно проводящие, клетки Пуркинье, водители ритма третьего и четвертого порядков): малые клетки Пуркинье → генерация ПД с частотой 30-40 имп/мин большие клетки Пуркинье → генерация ПД с частотой 20-30 имп/мин Общие морфологические особенности кардиомиоцитов в ПССОтносительно низкий уровень дифференцировки (по сравнению с сократительными кардиомиоцитами) Мало миофибрилл и они не упорядочены в пространстве Редуцирована Т-система Мало органелл общего назначения Частое отсутствие вставочных дисков Межклеточные соединения с помощью простых контактов Таблица 5 При повышенных систематических функциональных нагрузках общее количество клеток не возрастает, в цитоплазме увеличивается содержание органелл общего назначения и миофибрилл. Увеличивается также размер клеток (происходит функциональная гипертрофия) и возрастает степень плоидности ядер. Гипоталамус Гипоталамус (hypothalamus) является высшим нервным центром регуляции эндокринных функций. Он контролирует и интегрирует все висцеральные функции организма и объединяет эндокринные механизмы регуляции с нервными, будучи мозговым центром симпатического и парасимпатического отделов автономной нервной системы. Субстратом объединения нервной и эндокринной систем являютсянейросекреторные клетки, которые у высших позвоночных и человека располагаются в нейросекреторных ядрах гипоталамуса. Нейроэндокринные трансдукторы (переключатели) и нейрогемальные образования. Медиальное возвышение (eminentia medialis) является нейрогемальным органом гипоталамо-аденогипофизарной системы. Оно образовано эпендимой, отдельные глиальные клетки которой дифференцируются в танициты (tanicyti),отличающиеся разветвленными отростками, контактирующими с клубочками первичной капиллярной сети портальной системы гипоталамо-гипофизарного кровообращения. В гипоталамо-аденогипофизарной системе аккумулируются аденогипофизотропные нейрогормоны - нейротранс-миттеры (либерины и статины), вырабатываемые в мелкоклеточных ядрах среднего и заднего отделов гипоталамуса, которые затем поступают в портальную систему гипофиза. В гипоталамо-нейрогипофизарной системе аналогичным нейрогемальным органом оказывается нейрогипофиз (задняя доля гипофиза), где аккумулируются нонапептидные нейрогормоны (вазопрес-син - антидиуретический гормон и окситоцин), вырабатываемые в крупноклеточных ядрах переднего отдела гипоталамуса, в дальнейшем выделяемые в кровь. Секреторные нейроны (neuronum secretorium) расположены в ядрах серого вещества гипоталамуса. Строение секреторных нейронов описано в главе 10. Нервные ядра (свыше 30 пар) группируются в его переднем, среднем (медиобазальном и туберальном) и заднем отделах. В переднем гипоталамусе располагаются парные супраоптические (nuclei supraoptici) и паравентрикулярные (nuclei paraventriculares) ядра. Супраоптические ядра образованы крупными холинергическими (пептидохолинергическими) нейросекреторными клетками, содержащими как в перикарионах, так и в отростках секреторные гранулы. Аксоны этих клеток проходят через медиальную эминенцию и гипофизарную ножку в заднюю долю гипофиза, где заканчиваются на кровеносных капиллярах утолщенными терминалями. Паравентрикулярные ядра построены более сложно. Их центральная крупноклеточная часть образована такими же крупными холинергическими нейросекреторными клетками, как в супраоптическом ядре, и их аксонами, идущими в заднюю долю гипофиза. В обоих указанных ядрах крупные нейросекреторные клетки продуцируют белковые (нонапептидные) нейрогормоны - вазопрессин, или антидиуретический гормон (АДГ), и окситоцин. У человека выработка антидиуретического гормона совершается преимущественно в супраоптическом ядре, тогда как продукция окситоцина преобладает в крупноклеточной части паравентри-кулярных ядер. Периферическая же часть паравентрикулярного ядра состоит из мелких адренергических нейросекреторных клеток. Аксоны этих клеток направляются в медиальную эминенцию. В мелкоклеточных ядрах среднего (медиобазального и туберального) гипоталамуса их мелкие адренергические (пептидоадренергические) нейро-секреторные клетки вырабатывают аденогипофизотропные нейрогормоны - рилизинг-гормоны, или факторы (от англ. release - освобождать), с помощью которых гипоталамус контролирует деятельность аденогипофиза. Эти нейро-гормоны по своей природе являются низкомолекулярными олигопептидами и разделяются налиберины, стимулирующие выделение и, вероятно, продукцию гормонов передней и средней долей гипофиза, и статины, угнетающие функции аденогипофиза. Среди либеринов различают: соматотропин-рилизинг-фактор - соматолиберин; тиреотропин-рилизинг-фактор - тиролиберин; АКТГ-рилизинг-фактор - кортиколиберин; рилизинг-фактор фолликулости- мулирующего гормона - фоллиберин; рилизинг-фактор лютеинизирующего гормона - люлиберин; пролактин-рилизинг-фактор - пролактолиберин; рилизинг-фактор меланоцитостимулирующего гормона - меланолиберин. Среди статинов выделяют: соматотропин-ингибирующий фактор - сомато-статин; пролактинингибирующий фактор - пролактостатин; ингибирующий фактор меланоцитостимулирующего гормона - меланостатин. Важнейшие ядра среднего гипоталамуса локализуются в сером бугре (tuber cinereum) - аркуатное или инфундибулярное (nucl. arcuatus seu infundibularis), дугообразно охватывающее гипофизарную ножку, вентромедиальное (nucl. ventromedialis) и дорсомедиальное (nucl. dorsomedialis). Основным местом выработки гипотала-мических либеринов и статинов оказываются аркуатные и вентромедиальные ядра, а также мелкие пептидоадренергические клетки мелкоклеточной части паравентрикулярного ядра и аналогичные клетки в сером перивентрикуляр-ном веществе, в преоптической зоне гипоталамуса и в супрахиазматическом ядре. Аксоны мелких пептидоадренергических клеток, равно как и сопровождающих их адренергических нейронов обычного типа, направляются в медиальную эминенцию в составе тубероинфундибулярного дофаминер-гического пучка, берущего начало от дофаминергических нейронов тубе-ральных ядер . Регуляция гипоталамусом периферических эндокринных желез Влияние гипоталамуса на периферические эндокринные железы осуществляется преимущественно гуморально через секрецию рилизинг-факторов, которые попадают в переднюю долю гипофиза. Такой способ передачи называется трансаденогипофизарным. Кроме того, гипоталамус посылает свои эфферентные импульсы к регулируемым эффекторам прямо по симпатическим или парасимпатическим нервам последних, без опосредования гипофизом, т. е. парагипофизарно. Парагипофизарная регуляция эндокринных эффекторов может проявляться способностью эндокринных образований непосредственно реагировать (по принципу отрицательной обратной связи) на собственные гормоны, или иммунологические агенты, поступившие в циркуляцию, или на величину эффекта, вызванного ими в организме. Функции гипоталамуса находятся под контролем головного мозга. В его разных частях находятся нейроэндокринные клетки, которые вырабатывают нейропептиды (более 50). Среди них, например, энкефалин, является интернейрональным медиатором, под воздействием которого по цепочке нейронов происходит выработка нейроаминов - серотонина и норадрена-лина нейронами лимбической системы и норадреналина нейронами ретикулярной формации. Нейроамины влияют на секрецию гипоталамических нейрогормонов. Действие последних стимулирует или тормозит активность аденоцитов гипофиза. Так возникает тесная функциональная связь нервной и эндокринной систем, обеспечивающая контроль, интеграцию и реактивность живых систем.
Гипофиз. Гипофиз (питуитарная железа) вместе с гипоталамусом составляет гипоталамо-гипофизарную нейросекреторную систему. Он является мозговым придатком. В гипофизе различают аденогипофиз (переднюю долю, промежуточную и туберальную части) и нейрогипофиз (заднюю долю, воронку). Развитие. Аденогипофиз развивается из эпителия крыши ротовой полости. На 4-й неделе эмбриогенеза образуется эпителиальное выпячивание в виде гипофизарного кармана (карман Ратке), из которого вначале формируется железа с внешним типом секреции. Затем проксимальный отдел кармана редуцируется, и аденомер становится обособленной эндокринной железой. Нейрогипофиз образуется из материала инфундибулярной части дна Ш-го желудочка мозга и имеет нейральное происхождение. Эти две разные по происхождению части вступают в контакт, образуя гипофиз. Строение. Аденогипофиз состоит из эпителиальных тяжей — трабекул. Между ними проходят синусоидные капилляры. Клетки представлены хромофильными и хромофобными эндокриноцитами. Среди хромофильных эндокриноцитов различают ацидофильные и базофильные эндокриноциты. Ацидофильные эндокриноциты — это клетки средних размеров, округлой или овальной формы, с хорошо развитой гранулярной эндоплазматической сетью. Ядра находятся в центре клеток. Они содержат крупные плотные гранулы, окрашивающиеся кислыми красителями. Лежат эти клетки по периферии трабекул и составляют 30-35% от общего количества аденоцитов в передней доле гипофиза. Имеется две разновидности ацидофильных эндокриноцитов: соматотропоциты, вырабатывающие гормон роста (соматотропин), и лактотропоциты, или маммотропоциты, продуцирующие лактотропный гормон (пролактин). Соматотропин стимулирует процессы роста всех тканей и органов. При гиперфункции соматотропоцитов может развиться акромегалия и гигантизм, а в условиях гипофункции — замедление роста тела, что приводит к гипофизарной карликовости. Лактотропный гормон стимулирует секрецию молока в молочных железах и прогестерона в желтом теле яичника. Базофильные эндокриноциты — это крупные клетки, в цитоплазме которых есть гранулы, окрашивающиеся основными красителями (анилиновым синим). Они составляют 4-10% от общего числа клеток в передней доле гипофиза. В гранулах содержатся гликопротеины. Базофильные эндокриноциты подразделяются на тиротропоциты и гонадотропоциты. Тиротропоциты — это клетки с большим числом плотных мелких гранул, окрашивающихся альдегидфуксином. Они вырабатывают тиреотропный гормон. При недостатке в организме тиреоидных гормонов тиротропоциты трансформируются в клетки тиреоидэктомии с большим числом вакуолей. При этом усиливается продукция тиротропина. Гонадотропоциты — округлые клетки, в которых ядро смешено к периферии. В цитоплазме есть макула — светлое пятно, где расположен комплекс Гольджи. Мелкие секреторные гранулы содержат гонадотропные гормоны. При недостатке в организме половых гормонов в аденогипофизе появляются клетки кастрации, для которых характерна кольцевидная форма из-за наличия крупной вакуоли в цитоплазме. Такая трансформация гонадотропной клетки связана с ее гиперфункцией. Различаются две группы гонадотропоцитов, вырабатывающих либо фолликулостимулирующий, либо лютеинизирующий гормоны. Кортикотропоциты — это клетки неправильной, иногда отростчатой формы. Они разбросаны по всей передней доле гипофиза. В их цитоплазме определяются секреторные гранулы в виде пузырька с плотной сердцевиной, окруженного мембраной. Между мембраной и сердцевиной имеется светлый ободок. Кортикотропоциты вырабатывают АКТГ (адренокортикотропный гормон), или кортикотропин, активирующий клетки пучковой и сетчатой зон коры надпочечников. Хромофобные эндокриноциты составляют 50-60% от общего числа клеток аде-ногипофиза. Они располагаются в середине трабекул, имеют небольшие размеры, гранул не содержат, цитоплазма их слабо окрашивается. Это сборная группа клеток, среди которых находятся молодые хромофильные клетки, еще не накопившие гранул секрета, зрелые хромофильные клетки, уже выделившие секреторные гранулы, и резервные камбиальные клетки. Таким образом, в аденогипофизе обнаруживается система взаимодействующих клеточных дифферонов, формирующих ведущую эпителиальную ткань этой части железы. Средняя (промежуточная) доля гипофиза у человека развита слабо, составляя 2% от общего объема гипофиза. Эпителий в этой доле однородный, клетки богаты мукоидом. Местами есть коллоид. В промежуточной доле эндокриноцитами вырабатываются меланоцитостимулирующий гормон и липотропный гормон. Первый адаптирует сетчатку глаза к зрению в сумерках, а также активизирует кору надпочечников. Липотропный гормон стимулирует жировой обмен. Влияние нейропептидов гипоталамуса на эндокриноциты осуществляется с помощью системы гипоталамо-аденогипофизарного кровообращения (портальной). В первичную капиллярную сеть срединного возвышения секретируются гипоталамические нейропептиды, которые далее по портальной вене попадают в аденогипофиз и в его вторичную капиллярную сеть. Синусоидные капилляры последней расположены между эпителиальными тяжами эндокриноцитов. Так нейропептиды гипоталамуса воздействуют на клетки-мишени аденогипофиза. Нейрогипофиз имеет нейроглиальную природу, не является гормонпродуцирующей железой, а выполняет роль нейрогемального образования, в котором накапливаются гормоны некоторых нейросекреторных ядер переднего гипоталамуса. В задней доле гипофиза идут многочисленные нервные волокна гипоталамогипофизарного тракта. Это нервные отростки нейросекреторных клеток супраоптического и паравентрикулярного ядер гипоталамуса. Неройны этих ядер способны к нейросекреции. Нейросекрет (трансдуктор) транспортируется по нервным отросткам в заднюю долю гипофиза, где выявляется в виде телец Херринга. Аксоны нейросекреторных клеток заканчиваются в нейрогипофизе нейрососудистыми синапсами, посредством которых нейросекрет поступает в кровь. Нейросекрет содержит два гормона: антидиуретический (АДГ), или вазопрессин (он действует на нефроны, регулируя обратное всасывание воды, а также суживает сосуды, повышая кровяное давление); окситоцин, стимулирующий сокращение гладких мышц матки. Лекарственный препарат, полученный из задней доли гипофиза, называется питуитрином и используется для лечения несахарного мочеизнурения. В нейрогипофизе имеются нейроглиальные клетки, называемые питуицитами. Реактивность гипоталамо-гипофизарной системы. Боевые травмы и сопровождающие их стрессы приводят к сложным нарушениям нейроэндокринной регуляции гомеостаза. При этом нейросекреторные клетки гипоталамуса усиливают выработку нейрогормонов. В аденогипофизе уменьшается количество хромофобных эндокриноцитов, что ослабляет репаративные процессы в этом органе. Число базофильных эндокриноцитов нарастает, а в ацидофильных эндокриноцитах появляются крупные вакуоли, свидетельствующие об их напряженном функционировании. При длительном лучевом поражении в железах внутренней секреции возникают деструктивные и  зменения секреторных клеток и угнетение их функции. зменения секреторных клеток и угнетение их функции.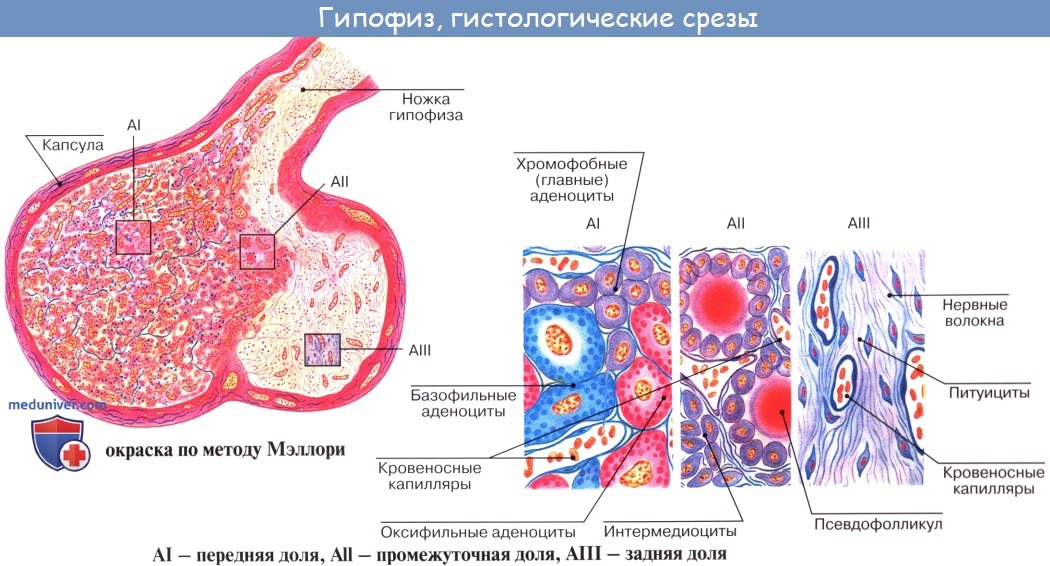 Надпочечники Надпочечники представляют собой парные железы, состоящие из коркового и мозгового вещества. Каждая из этих частей является самостоятельной железой внутренней секреции, вырабатывающей свои гормоны — регуляторы защитно-приспособительных реакций организма. Корковое вещество — это аденогипофиззависимая, а мозговое вещество — аденогипофизнезависимая эндокринные железы. Развитие надпочечников. Корковая часть надпочечников развивается на 5-й неделе эмбриогенеза из участка целомического эпителия в области корня брыжейки у краниального полюса правой и левой первичной почки. Отсюда происходит другое название коры надпочечников — интерреналовое тело. Вначале образуется первичная (или феталь-ная) кора, состоящая из крупных ацидофильных эндокриноцитов. Начиная с 10-й недели эмбриогенеза из того же источника формируется дефинитивная кора надпочечников за счет мелких базофильных эндокриноцитов, окружающих снаружи первичную кору. В эмбриогенезе толщина фетальнои коры значительной превосходит таковую дефинтивной коры. Однако после рождения эндокриноциты фетальнои коры погибают механизмом апоптоза, поэтому толщина коры уменьшается. Полное развитие коры происходит после полового созревания. Источник развития коркового вещества надпочечников находится вблизи от зачатка гонад, с чем связана способность клеток сетчатой зоны коры вырабатывать андрогенный гормон близкий по свойствам тестостерону. Мозговое вещество надпочечников образуется несколько позже (на 6-7-й неделе эмбриогенеза) из общего с симпатическими ганглиями зачатка — нервного гребня. Симпатобласты мигрируют в интерреналовое тело, размножаются, формируя мозговые эндокриноциты хромаффинной ткани. Строение надпочечников. Надпочечники покрыты соединительнотканной капсулой, под которой находится слой малодифференцированных клеток. Корковое вещество состоит из системы эпителиальных тяжей. Между ними по соединительнотканным прослойкам проходят кровеносные капилляры. В коре надпочечника различают три зоны: клубочковую, пучковую и сетчатую. Клубочковая, или наружная, зона располагается под капсулой. Корковые эндокриноциты (адренокортикоциты) образуют здесь аркады, или клубки. В цитоплазме этих клеток хорошо развита агранулярная эндоплазматическая сеть, что вообще характерно для клеток, синтезирующих стероидные гормоны. Эндокриноциты клубочковой зоны вырабатывают минералокортикоиды (альдостерон и другие). Альдостерон регулирует уровень натрия в организме, предотвращая его выведение. Эндокриноциты клубочковой зоны получают сигналы об изменениях уровня натрия в крови через ангиотензиновую систему, и таким образом их деятельность оказывается связанной с функцией юкстагломерулярного гистиона почки. Минералокортикоиды влияют на водно-солевой обмен, усиливают воспаление и образование коллагена. В клубочковой зоне отмечается много митозов эпителиальных клеток. На границе между клубочковой и пучковой зонами расположен суданофобный слой. Здесь также находятся малодифференцированные эпителиоциты.  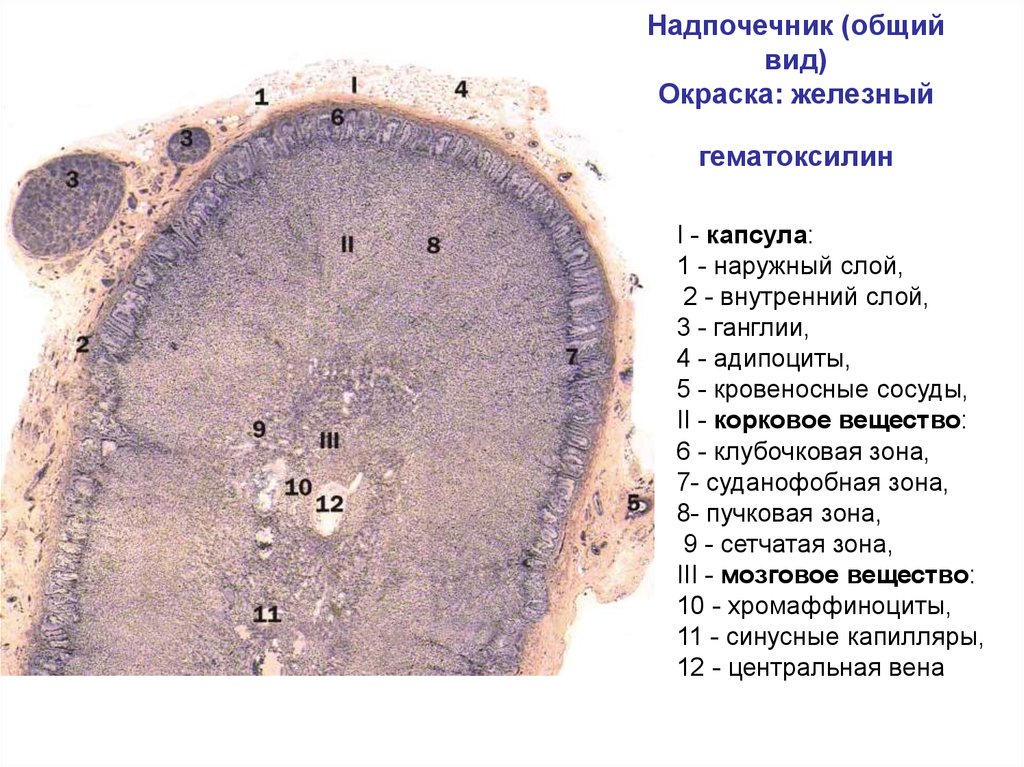 Щитовидная железа Щитовидная железа вырабатывает тиреоидные гормоны тироксин и трийодтиронин, а также тирокальцитонин (кальцитонин). Тироксин и трийодтиронин являются мощными стимуляторами окислительных процессов в клетках, причем трийодтиронин в 5-10 раз активнее тироксина. Эти гормоны усиливают обмен веществ, синтез белков, газообмен, обмен углеводов и жиров. Тиреоидные гормоны оказывают значительное влияние на развитие, рост и дифференцировку клеток и тканей. Они ускоряют развитие костной ткани. Особенно большое влияние гормоны щитовидной железы оказывают на гистогенез нервной ткани. При недостаточности щитовидной железы тормозится дифференцировка клеток и тканей головного мозга, нарушается психическое развитие человека. Тиреоидные гормоны оказывают стимулирующее действие на регенерационные процессы в тканях. В состав тироксина и трийодтиронина в качестве обязательного ингредиента входит йод, поэтому поступление йода с питьевой водой и пищей в организм необходимо для нормальной деятельности щитовидной железы. Не содержащий йода третий гормон щитовидной железы — тирокальцитонин участвует в регуляции кальциевого и фосфорного обмена. Развитие щитовидной железы. Зачаток щитовидной железы появляется на 4-й неделе эмбриогенеза в виде выпячивания вентральной стенки глоточной кишки между 1-й и 2-й парами жаберных карманов. Выпячивание это превращается в эпителиальный тяж с утолщением на конце. Щитовидная железа вначале закладывается как экзокринная железа. Позднее проток, соединяющий железу с глоткой, редуцируется, и от этого протока остается лишь foramen cecum. Эпителиальный тяж раздваивается на конце. В течение 3-го месяца по ходу эпителиальных тяжей образуются перетяжки. В четкообразных сегментах этих тяжей появляются просветы. По мере углубления перетяжек эпителиальные тяжи распадаются на отдельные фолликулы и клеточные островки. В конце 3-го месяца эпителий фолликулов дифференцируется. Клетки его — фолликулярные эндокриноциты (тироциты) — начинают вырабатывать гормоны, которые накапливаются в полости фолликула. Образование новых фолликулов и их развитие происходят гетерохронно. К моменту рождения отмечается наличие фолликулов с коллоидом ("коллоидный тип строения") и без коллоида с десквамацией эпителия ("десквамативный тип строения"). Между фолликулами встречаются межфолликулярные островки клеток. В процессе развития железы наряду с дифференцировкой эпителия происходит разрастание мезенхимы, преобразующейся в соединительную ткань. Формируется строма железы, содержащая густую сеть капилляров. В строму проникают нервные волокна. В зачаток железы врастают также производные 5-й пары жаберных карманов — клетки так называемых ультимобранхиальных телец. Это С-клетки, вырабатывающие кальцитонин. Это клетки нейроэктодермальной природы, и в закладку щитовидной железы они внедряются через ультимобранхиальные зачатки. Таким образом, в формировании структурно-функциональных единиц щитовидной железы участвуют следующие клеточные диффероны: это ведущие клетки эпителия фолликулов — фолликулярные эндокриноциты, вырабатывающие тироксин и трийодтиронин; кальцитониноциты, или С-клетки, продуцирующие кальцитонин и другие пептиды (соматостатин, тиролиберин, серотонин и др.). Строение щитовидной железы. Железа, состоящая из двух долей, снаружи покрыта соединительнотканной капсулой, от которой отходят перегородки, подразделяющие паренхиму на дольки. Структурно-функциональной единицей щитовидной железы является фолликул. Средний диаметр фолликулов около 50 мкм. Форма их преимущественно округлая. Фолликулы представляют собой замкнутые пузырьки. Стенка их образована однослойным эпителием, состоящим из фолликулярных эндокриноцитов (тироцитов). Среди этих клеток в виде небольших скоплений находятся С-клетки. Они могут быть и около фолликулов, и между фолликулами. Полость фолликула заполнена продуктом секреции тироцитов — коллоидом, содержащим белки — тироглобулины. Снаружи фолликулы оплетают сети кровеносных и лимфатических капилляров. Между соседними фолликулами встречаются интерфолликулярные островки, состоящие из малодифференцированных эндокриноцитов. Фолликулярные эндокриноциты имеют кубическую форму и округлое ядро. На апикальной поверхности их имеются микроворсинки. В цитоплазме хорошо развиты органеллы, обеспечивающие синтез белков. Много свободных рибосом, образующих полисомы. Соседние эндокриноциты в стенке фолликула соединяются при помощи плотных контактов, десмосом и интердигитаций. Особенностью гистофизиологии щитовидной железы является различно направленное движение секреторных продуктов: вначале в полость фолликула, а затем в противоположном направлении — в кровь. Происходит это благодаря активной деятельности фолликулярных эндокриноцитов. Секреторный цикл этих клеток состоит из следующих фаз: поглощение исходных веществ, синтез секрета, выделение его в полость фолликула в виде коллоида, иодирование коллоида, эндоци-тоз иодированного коллоида и его модификация и выведение гормона через ба-зальную часть клетки в окружающие ткани и кровеносные и лимфатические капилляры. Выработка тиреоидных гормонов начинается с синтеза тиреоглобули-на в базальной части эндокриноцитов. Содержащие тиреоглобулин продукты синтеза из эндоплазматической сети поступают в комплекс Гольджи и далее в виде секреторных гранул путем экзоцитоза выводятся в полость фолликула. К нейодированному тиреоглобулину присоединяется сначала один атом йода, а затем и второй, в результате чего образуются моно- и дийодтиронины. Последующая комплексация дает тетрайодтиронин, или тироксин. При отщеплении одного атома йода от тироксина образуется трийодтиронин. В нормальных условиях процессы образования коллоида и его резорбции уравновешены. Это равновесие, однако, нарушается при гипо- и гиперфункции щитовидной железы. При гипофункции задерживается выведение гормона. При этом фолликулы обычно имеют большие размеры, в полости фолликула коллоида много, он густой, не имеет резорбционных вакуолей, эпителий представлен плоскими эндокриноцитами. При гиперфункции, наоборот, усиливается выведение гормона-коллоида в кровь. Коллоид разжижается, количество его невелико, эпителий стенки фолликула становится высоким призматическим. Парафолликулярные эндокриноциты (С-клетки, или кальцитониноциты) вырабатывают белковый гормон кальцитонин. Он понижает уровень кальция в крови и является антагонистом паратирина. Кальцитонин действует на остеокласты костной ткани, уменьшая их резорбтивную активность. С-клетки — это более светлые и более крупные, чем фолликулярные эндокриноциты, и доля их составляет 0,1%. Обычно они лежат поодиночке или мелкими группами. В цитоплазме парафолликулярных эндокриноцитов много аргирофильных или осмиофильных секреторных гранул, содержащих кальцитонин и другие пептиды. Строма железы состоит из рыхлой волокнистой соединительной ткани, в которой часто обнаруживаются тучные клетки, макрофаги, лимфоциты. Фибробласты щитовидной железы обладают рядом уникальных свойств, что определяется особенностями рецепторно-трансдукторной системы клеток. В частности, они могут участвовать в развитии воспаления наряду с иммунокомпетентными клетками. Хорошо развиты капилляры фенестрированного типа. Нервные волокна содержат пептиды, характерные для С-клеток. | ||||


