Строение и уровни организации ДНК. Строение и уровни организации ДНК предыдущая
 Скачать 1.72 Mb. Скачать 1.72 Mb.
|

|
Строение и уровни организации ДНК Предыдущая: Нуклеиновые кислоты. Общие сведения. Нуклеиновые кислоты - высокомолекулярные вещества, состоящие из мононуклеотидов, которые соединены друг с другом в полимерную цепочку с помощью 3',5'- фосфодиэфирных связей и упакованы в клетках определенным образом. Нуклеиновые кислоты - биополимеры двух разновидностей: рибонуклеиновая кислота (РНК) и дезоксирибонуклеиновая кислота (ДНК). Каждый биополимер состоит из нуклеотидов, различающихся по углеводному остатку (рибозе, дезоксирибозе) и одному из азотистых оснований (урацил, тимин). Соответственно этим различиям нуклеиновые кислоты и получили свое название. 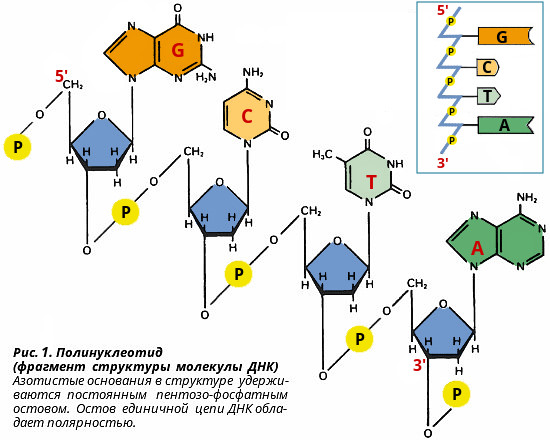 Структура дезоксирибонуклеиновой кислоты Нуклеиновые кислоты имеют первичную, вторичную и третичную структуру. Первичная структура ДНК Первичной структурой ДНК называют линейную полинуклеотидную цепь, в которой мононуклеотиды соединены 3', 5'-фосфодиэфирными связями. Исходным материалом при сборке цепи нуклеиновой кислоты в клетке является нуклеозид 5'-трифосфат, который в результате удаления β и γ остатков фосфорной кислоты способен присоединить 3'-атом углерода другого нуклеозида. Таким образом, 3'-атом углерода одной дезоксирибозы ковалентно связывается с 5'-атомом углерода другой дезоксирибозы посредством одного остатка фосфорной кислоты и образует линейную полинуклеотидную цепь нуклеиновой кислоты. Отсюда и название: 3', 5'-фосфодиэфирные связи. Азотистые основания не принимают участия в соединении нуклеотидов одной цепи (рис. 1.). Такое соединение, между остатком молекулы фосфорной кислоты одного нуклеотида и углеводом другого, приводит к образованию пентозо-фосфатного скелета молекулы полинуклеотида, на котором сбоку один за другим присоединяются азотистые основания. Их последовательность расположения в цепях молекул нуклеиновых кислот строго специфична для клеток разных организмов, т.е. носит видовой характер (правило Чаргаффа). Линейная цепь ДНК, длина которой зависит от числа входящих в цепь нуклеотидов, имеет два конца: один называется 3'-концом и содержит свободный гидроксил, а другой - 5'-концом, содержит остаток фосфорной кислоты. Цепь полярна и может иметь напрвление 5'->3' и 3'->5'. Исключением являются кольцевые ДНК. Генетический "текст" ДНК составлен с помощью кодовых "слов" - триплетов нуклеотидов, называемых кодонами. Участки ДНК, содержащие информацию о первичной структуре всех типов РНК, называют структурными генами. Полинуклеодитные цепочки ДНК достигают гигантских размеров, поэтому в клетке они упакованы определенным образом.
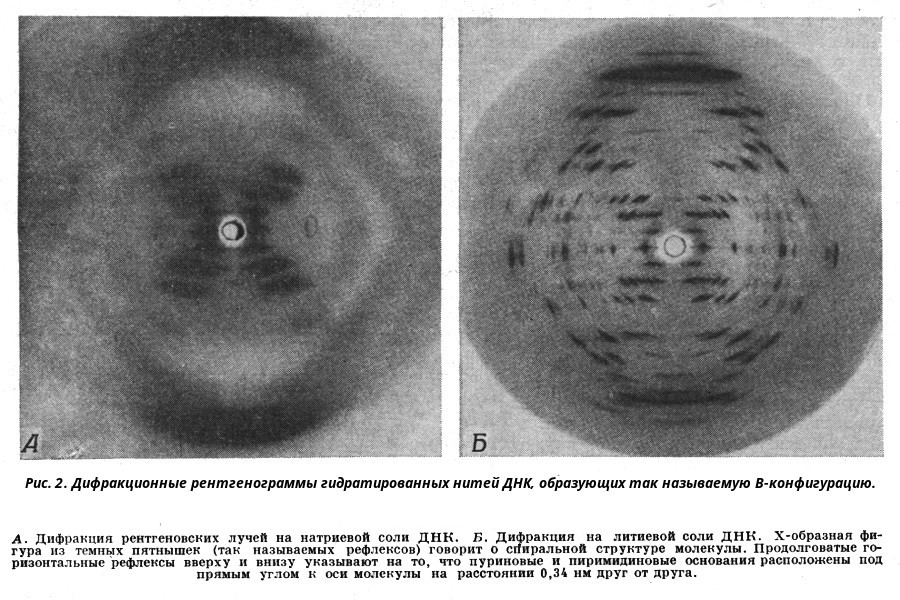 Вторичная структура ДНК Вторичная структура ДНК - это двойная спираль, модель которой была предложена Д.Уотсоном и Ф.Криком в 1953 году. Предпосылки к созданию модели ДНК В результате первоначальных анализов сложилось представление, что ДНК любого происхождения содержит все четыре нуклеотида в равных молярных количествах. Однако в 1940-х годах Э. Чаргафф и его сотрудники в результате анализа ДНК, выделенных из разнообразных организмов, ясно показали, что азотистые основания содержатся в них в различных количественных соотношениях. Чаргафф нашел, что, хотя эти соотношения одинаковы для ДНК из всех клеток одного и того же вида организмов, ДНК от разных видов могут заметно различаться по содержанию тех или иных нуклеотидов. Это наводило на мысль, что различия в соотношении азотистых оснований, возможно, связаны с каким-то биологическим кодом. Хотя соотношение отдельных пуриновых и пиримидиновых оснований в различных образцах ДНК оказалось неодинаковым, при сравнении результатов анализов выявилась определенная закономерность: во всех образцах общее количество пуринов было равно общему количеству пиримидинов (А + Г = Т + Ц), количество аденина - количеству тимина (А = Т), а количество гуанина - количеству цитозина (Г = Ц). ДНК, выделенная из клеток млекопитающих, была в целом богаче аденином и тимином и относительно беднее гуанином и цитозином, тогда как у бактерий ДНК была богаче гуанином и цитозином и относительно беднее аденином и тимином. Эти данные составили важную часть фактического материала, на основе которого позднее была построена модель структуры ДНК Уотсона - Крика. Еще одним важным косвенным указанием на возможную структуру ДНК послужили данные Л. Полинга о строении белковых молекул. Полинг показал, что возможно несколько различных устойчивых конфигураций аминокислотной цепи в белковой молекуле. Одна из распространенных конфигураций пептидной цепи - α-спираль - представляет собой правильную винтообразную структуру. При такой структуре возможно образование водородных связей между аминокислотами, находящимися на смежных витках цепи. Полинг описал α-спиральную конфигурацию полипептидной цепи в 1950 году и высказал предположение, что и молекулы ДНК, вероятно, имеют спиральную структуру, закрепленную водородными связями. Однако наиболее ценные сведения о строении молекулы ДНК дали результаты рентгеноструктурного анализа. Рентгеновские лучи, проходя сквозь кристалл ДНК, претерпевают дифракцию, т. е. отклоняются в определенных направлениях. Степень и характер отклонения лучей зависят от структуры самих молекул. Дифракционная рентгенограмма (рис. 3) дает опытному глазу ряд косвенных указаний относительно строения молекул исследуемого вещества. Анализ дифракционных рентгенограмм ДНК привел к заключению, что азотистые основания (имеющие плоскую форму) уложены наподобие стопки тарелок. Рентгенограммы позволили выявить в структуре кристаллической ДНК три главных периода: 0,34, 2 и 3,4 нм. 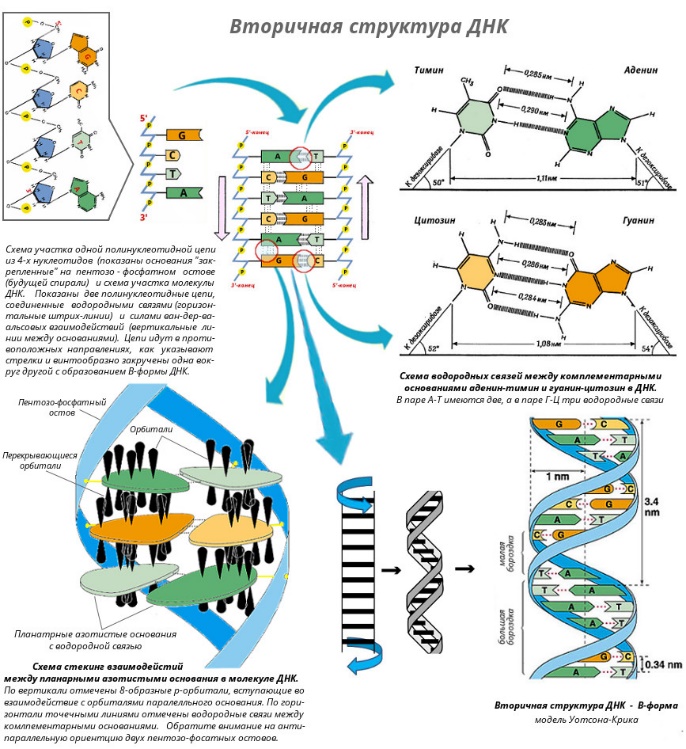 Модель ДНК Уотсона-Крика Исходя из аналитических данных Чаргаффа, рентгенограмм, полученных Уилкинсом и исследований химиков, предоставивших сведения о точных расстояниях между атомами в молекуле, об углах между связями данного атома и о величине атомов, Уотсон и Крик начали строить физические модели отдельных составных частей молекулы ДНК в определенном масштабе и "подгонять" их друг к другу с таким расчетом, чтобы полученная система соответствовала различным экспериментальным данным [показать] . В результате своих исследований, в 1953 году Уотсон и Крик предложили модель строения молекулы ДНК (рис. 3), которая остается актуальной по настоящее время. Согласно модели молекула ДНК состоит из двух комплементарных полинуклеотидных цепей. Каждая цепь ДНК представляет полинуклеотид, состоящий из нескольких десятков тысяч нуклеотидов. В ней соседние нуклеотиды образуют регулярный пентозо-фосфатный остов за счет соединения остатка фосфорной кислоты и дезоксирибозы прочной ковалентной связью. Азотистые основания одной полинуклеотидной цепи при этом располагаются в строго определенном порядке против азотистых оснований другой. Чередование азотистых оснований в полинуклеотидной цепи нерегулярно. Расположение азотистых оснований в цепи ДНК является комплементарным (от греч. "комплемент" - дополнение), т.е. против аденина (А) всегда оказывается тимин (Т), а против гуанина (Г) - только цитозин (Ц). Это объясняется тем, что А и Т, а также Г и Ц строго соответствуют друг другу, т.е. дополняют друг другу. Такое соответствие задается химической структурой оснований, позволяющей образовать водородные связи в паре пурина и пиримидина. Между А и Т возникают две связи, между Г и Ц - три. Эти связи обеспечивают частичную стабилизацию молекулы ДНК в пространстве. Устойчивость двойной спирали при этом прямо пропорциональна числу связей G≡С, являющихся более стабильными по сравнению со связями А=Т. Известная последовательность расположения нуклеотидов в одной цепи ДНК позволяет по принципу комплементарности установить нуклеотиды другой цепи. Кроме того, установлено, что азотистые основания, имеющие ароматическую структуру, в водном растворе располагаются один над другим, формируя как бы стопку монет. Такой процесс формирования стопок из органических молекул называется стекинг. Полинуклеотидные цепи молекулы ДНК рассматриваемой модели Уотсона-Крика имеют аналогичное физико-химическое состояние, их азотистые основания располагаются в виде стопки монет, между плоскостями которых возникают ван-дер-ваальсовы взаимодействия (стекинг-взаимодействия). Водородные связи между комплементарными основаниями (по горизонтали) и стекинг-взаимодействие между плоскостями оснований в полинуклеотидной цепи за счет ван-дер-ваальсовых сил (по вертикали) обеспечивает молекуле ДНК дополнительную стабилизацию в пространстве. Сахарофосфатные остовы обеих цепей обращены наружу, а основания внутрь, навстречу друг другу. Направление цепей в ДНК антипараллельно (одна из них имеет направление 5'->3', другая - 3'->5', т.е. 3'-конец одной цепи расположен напротив 5'-конца другой.). Цепи образуют правые спирали с общей осью. Один виток спирали составляет 10 нуклеотидов, размер витка 3,4 нм, высота каждого нуклеотида 0,34 нм, диаметр спирали – 2,0 нм. В результате вращения одной цепи вокруг другой, образуется большая борозда (диаметром около 20 Å ) и малая борозда (около 12 Å) двойной спирали ДНК. Такая форма двойной спирали Уотсона-Крика в дальнейшем получила название В-формы. В клетках ДНК обычно существует в В-форме, которая является самой стабильной. Функции ДНК Предложенная модель объясняла многие биологические свойства дезоксирибонуклеиновой кислоты, в том числе хранение генетической информации и многообразие генов, обеспечиваемое большим разнообразием последовательных сочетаний 4-х нуклеотидов и фактом существования генетического кода, способность к самовоспроизведению и передаче генетической информации, обеспечиваемое процессом репликации, и реализацию генетической информации в виде белков, а также любых других соединений, образующихся с помощью белков-ферментов. Oсновные функции ДНК.
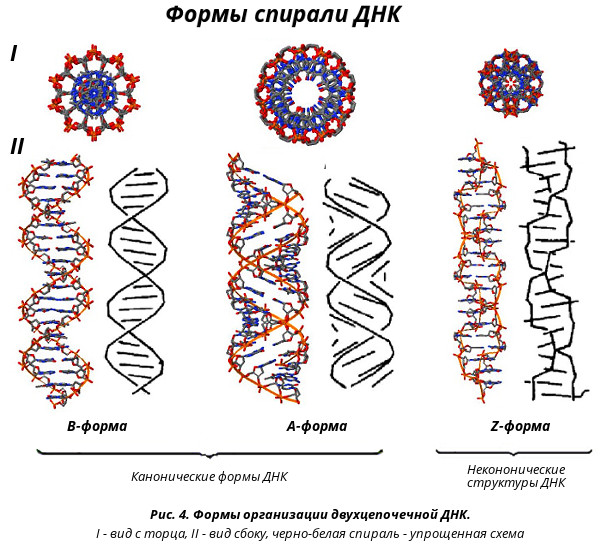 Формы организации двухцепочечной ДНК ДНК может формировать несколько типов двойных спиралей (рис.4). В настоящее время уже известно шесть форм (от А до Е и Z-форма). Структурные формы ДНК, как установила Розалинда Франклин, зависят от насыщения водой молекулы нуклеиновой кислоты. В исследованиях волокон ДНК при помощи рентгеноструктурного анализа было показано, что рентгенограмма радикальным образом зависит от того, при какой относительной влажности, при какой степени насыщения водой этого волокна происходит эксперимент. Если волокно было достаточно насыщено водой, то получалась одна рентгенограмма. При высушивании возникала совершенно другая рентгенограмма, сильно отличающаяся от рентгенограммы волокна высокой влажности. Молекула ДНК высокой влажности получила название В-формы. При физиологических условиях (низкая концентрация соли, высокая степерь гидратации) доминирующим структурным типом ДНК является В-форма (основная форма двухцепочечной ДНК - модель Уотсона-Крика). Шаг спирали такой молекулы равен 3,4 нм. На виток приходится 10 комплементарных пар в виде скрученных стопок "монет" - азотистых оснований. Стопки удерживаются водородными связями между двумя противолежащими "монетами" стопок, и "обмотаны" двумя лентами фосфодиэфирного остова, закрученными в правую спираль. Плоскости азотистых оснований перпендикулярны оси спирали. Соседние комплементарные пары повернуты друг относительно друга на 36°. Диаметр спирали 20Å, причем пуриновый нуклеотид занимает 12Å, а пиримидиновый - 8Å. Молекула ДНК более низкой влажности получила название А-формы. А-форма образуется в условиях менее высокой гидратации и при более высоком содержании ионов Na+ или К+. Эта более широкая правоспиральная конформация имеет 11 пар азотистых оснований на виток. Плоскости азотистых оснований имеют более сильный наклон к оси спирали, они отклонены от нормали к оси спирали на 20°. Отсюда следует наличие внутренней пустоты диаметром 5Å. Расстояние между соседними нуклеотидами составляет 0,23 нм, длина витка – 2,5 нм, диаметр спирали – 2,3 нм. Первоначально считали, что А-форма ДНК менее важна. Однако в дальнейшем выяснилось, что А-форма ДНК, также как и В-форма, имеет огромное биологическое значение. А-форму имеет спираль РНК-ДНК в комплексе матрица-затравка, а также спираль РНК-РНК и шпилечные структуры РНК (2’-гидроксильная группа рибозы не позволяет молекулам РНК образовывать В-форму). А-форма ДНК обнаружена в спорах. Установлено, что А-форма ДНК в 10 раз устойчивее к действию УФ-лучей, чем В-форма. А-форму и В-форму называют каноническими формами ДНК. Формы С-Е также правоспиральные, их образование можно наблюдать только в специальных экспериментах, и, по-видимому, они не существуют in vivo. С-форма ДНК имеет структуру, сходную с В-ДНК. Число пар оснований на виток составляет 9,33, длина витка спирали равна 3,1 нм. Пары оснований наклонены на угол 8 градусов относительно перпендикулярного положения к оси. Желобки по размерам близки к желобкам В-ДНК. При этом главный желобок несколько мельче, а минорный желобок – глубже. В С-форму могут переходить природные и синтетические полинуклеотиды ДНК.
Структурные элементы ДНК (неканонические структуры ДНК) К структурным элементам ДНК можно отнести необычные структуры, ограниченные какими-то специальными последовательностями:
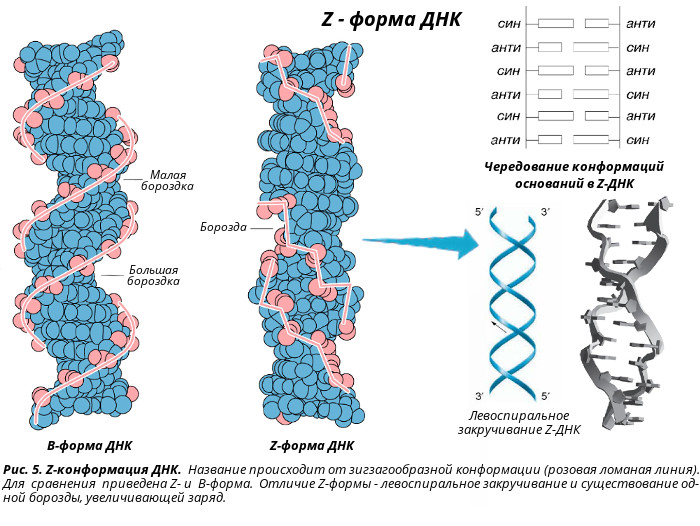 Z-форма ДНК была открыта в 1979 году при изучении гексануклеотида d(CG)3 - [CG-CG-CG]. Ее открыл профессор Массачусетского технологического института Александр Рич с сотрудниками. Z-форма стала одним из важнейших структурных элементов ДНК в связи с тем, что ее образование наблюдалось в участках ДНК, где пурины чередуются с пиримидинами (например, 5’-ГЦГЦГЦ-3’), или в повторах 5’-ЦГЦГЦГ-3’, содержащих метилированный цитозин. Существенным условием образования и стабилизации Z-ДНК являлось присутствие в ней пуриновых нуклеотидов в син-конформации, чередующихся с пиримидиновыми основаниями в анти-конформации. Природные молекулы ДНК в основном существуют в правой В-форме, если они не содержат последовательностей типа (ЦГ)n. Однако, если такие последовательности входят в состав ДНК, то эти участки при изменении ионной силы раствора или катионов, нейтрализующих отрицательный заряд на фосфодиэфирном каркасе, могут переходить в Z-форму, при этом другие участки ДНК в цепи остаются в классической В-форме. Возможность такого перехода указывает на то, что две цепи в двойной спирали ДНК находятся в динамическом состоянии и могут раскручиваться друг относительно друга, переходя из правой формы в левую и наоборот. Биологические следствия такой лабильности, допускающей конформационные превращения структуры ДНК пока не вполне понятны. Полагают, что участки Z-ДНК играют определенную роль в регуляции экспрессии некоторых генов и принимают участие в генетической рекомбинации. Z-форма ДНК - это левозакрученная двойная спираль, в которой фосфодиэфирный остов расположен зигзагообразно вдоль оси молекулы. Отсюда и название молекулы (zigzag)-ДHK. Z-ДНК - наименее скрученная (12 пар оснований на виток) и наиболее тонкая из известных в природе. Расстояние между соседними нуклеотидами составляет 0,38 нм, длина витка – 4,56 нм, диаметр Z-ДНК – 1,8 нм. Кроме того, внешний вид этой молекулы ДНК отличается наличием одной бороздки. Z-форма ДНК была обнаружена в клетках прокариот и эукариот. В настоящее время получены антитела, способные отличать Z-форму от В-формы ДНК. Эти антитела связываются с определенными участками гигантских хромосом клеток слюнных желез дрозофилы (Dr. melanogaster). За реакцией связывания легко следить из-за необычного строения этих хромосом, у которых более плотные участки (диски) контрастируют с менее плотными (междисками). Участки Z-ДНК расположены в междисках. Из этого следует, что Z-форма реально существует в естественных условиях, хотя размеры индивидуальных участков Z-формы пока неизвестны. 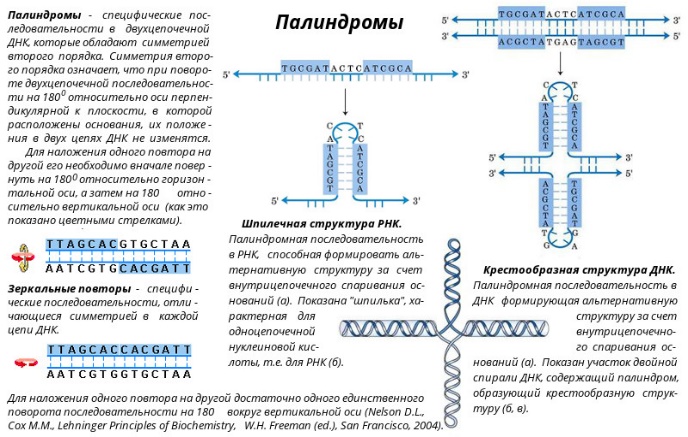 Палиндромы (перевертыши) - наиболее известные и часто встречающиеся в ДНК последовательности оснований. Палиндромом называют слово или фразу, которое читается слева направо и наоборот одинаково. Примерами таких слов или фраз являются: ШАЛАШ, КАЗАК, ПОТОП, А РОЗА УПАЛА НА ЛАПУ АЗОРА. В применении к участкам ДНК данный термин (палиндром) означает одинаковое чередование нуклеотидов вдоль цепи справа налево и слева направо (подобно буквам в слове "шалаш" и пр.). Палиндром характеризуется наличием инвертированных повторов последовательностей оснований имеющих симметрию второго порядка относительно двух цепей ДНК. Такие последовательности, по вполне понятной причине, являются самокомплементарными и имеют склонность к образованию шпилечных или крестообразных структур (рис.). Шпильки помогают регуляторным белкам узнавать место списывания генетического текста ДНК хромосом. В тех случаях, когда инвертированный повтор присутствует в одной и той же цепи ДНК такая последовательность называется зеркальным повтором. Зеркальные повторы не обладают свойствами самокомплементарности и, поэтому не способны к формированию шпилечных или крестообразных структур. Последовательности такого типа обнаружены практически во всех крупных молекулах ДНК и могут включать от всего нескольких пар оснований до нескольких тысяч пар оснований. Присутствие палиндромов в виде крестообразных структур в эукариотических клетках не доказано, хотя некоторое количество крестообразных структур обнаружено в условиях in vivo в клетках E. coli. Наличие в составе РНК или одноцепочечных ДНК самокомплементарных последовательностей служит основной причиной сворачивания в растворах нуклеиновой цепи в определенную пространственную структуру, отличающуюся формированием множества "шпилек". 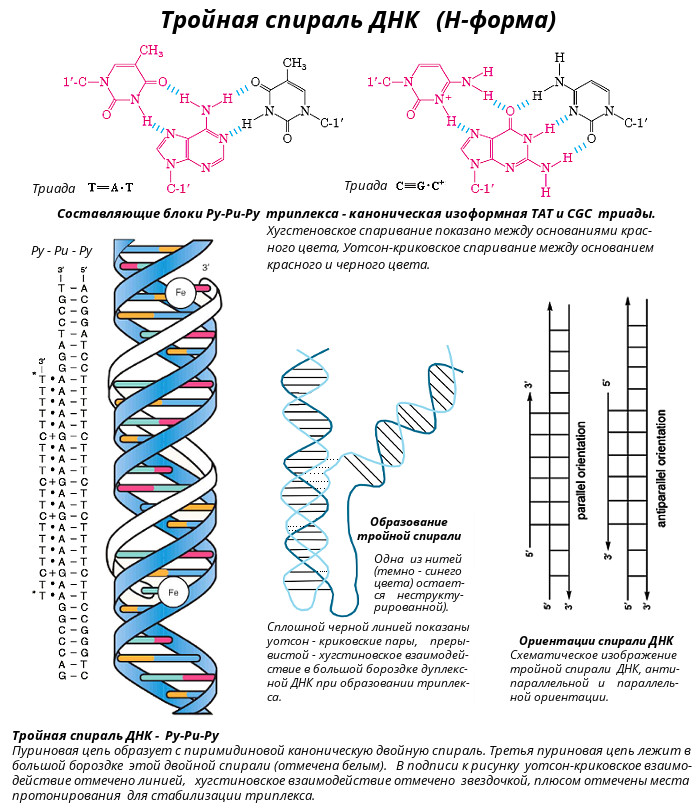 Н-форма ДНК - это спираль, которую образуют три цепи ДНК - тройная спираль ДНК. Представляет собой комплекс уотсон-криковской двойной спирали с третьей одноцепочечной нитью ДНК, которая укладывается в ее большой желобок, с образованием так называемой хугстиновской пары. Образование подобного триплекса происходит в результате сложения двойной спирали ДНК таким образом, что половина ее участка остается в виде двойной спирали, а вторая половина разъединяется. При этом одна из разъединенных спиралей образует новую структуру с первой половиной двойной спирали - тройную спираль, а вторая оказывается неструктурированной, в виде однонитевого участка. Особенностью этого структурного перехода является резкая зависимость от рН среды, протоны которой стабилизируют новую структуру. В силу этой особенности новая структура получила название Н-формы ДНК, образование которой было обнаружено в сверхспирализованных плазмидах, содержащих гомопурин-гомопиримидиновые участки, представляющие собой зеркальный повтор. В дальнейших исследованиях была установлена возможность осуществления структурного перехода некоторых гомопурин-гомопиримидиновых двунитиевых полинуклеотидов с образованием трехнитиевой структуры, содержащей:
Составляющие блоки Py-Pu-Py триплекса - канонические изоморфные CGC+ и TAT триады. Стабилизация триплекса требует протонирования триады CGC+, поэтому эти триплексы зависят от рН раствора.
Составляющие блоки Py-Pu-Pu триплекса - канонические изоморфные CGG и TAA триад. Существенным свойством Py-Pu-Pu триплексов является зависимость их стабильности от присутствия двухзарядных ионов, причем для стабилизации триплексов разной последовательности необходимы различные ионы. Поскольку для образования Py-Pu-Pu триплексов не требуется протонирования входящих в их состав нуклеотидов, такие триплексы могут существовать при нейтральных pH. Прим.: прямое и обратное хугстиновское взаимодействие объясняется симметрией 1-метилтимина: поворот на 180° приводит к тому, что место атома О4 занимает атом О2, при этом система водородных связей сохраняется. Известны два вида тройных спиралей:
Химически гомологичные цепи как в Py-Pu-Pu, так и в Py-Pu-Py триплексах, находятся в антипараллельной ориентации. Это в дальнейшем было подтверждено данными ЯМР спектроскопии. 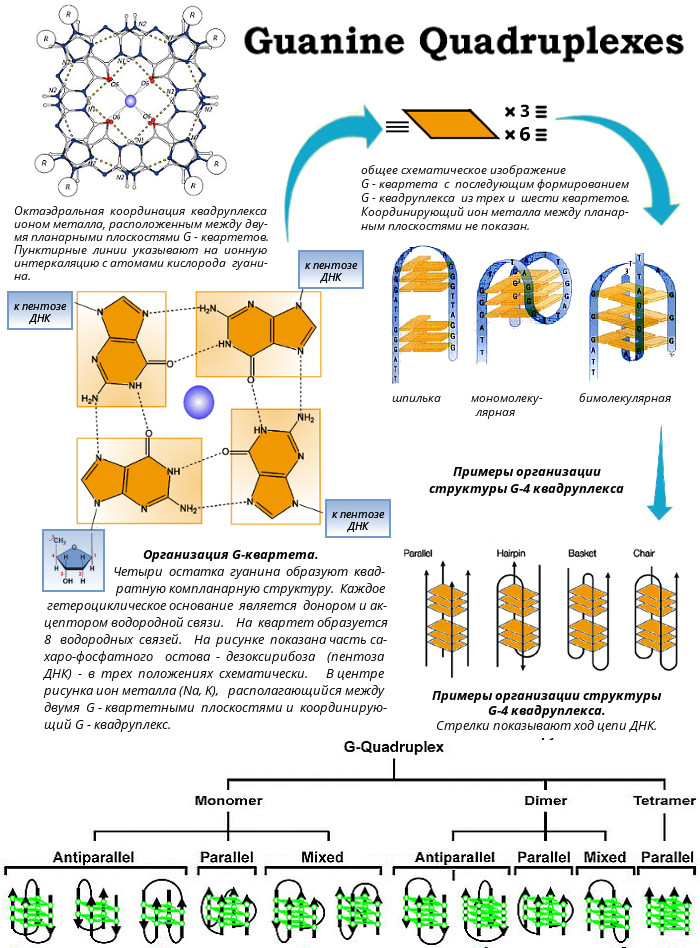 G-квадруплекс - 4-х спиральная ДНК. Такая структура образуется в случае, если имеются четыри гуанина, которые образуют так называемый G-квадруплекс — хоровод из четырех гуанинов. Первые намеки на возможность образования таких структур были получены задолго до прорывной работы Уотсона и Крика — еще в 1910 году. Тогда немецкий химик Ивар Банг обнаружил, что один из компонентов ДНК — гуанозиновая кислота — при высоких концентрациях образует гели, в то время как другие составные части ДНК таким свойством не обладают. В 1962 году с помощью рентгеноструктурного метода удалось установить структуру ячейки этого геля. Она оказалась составлена из четырех остатков гуанина, связывающих друг друга по кругу и образующих характерный квадрат. В центре связь поддерживает ион металла (Na, K, Mg). Такие же структуры могут образовываться и в ДНК, если в ней много гуанина. Эти плоские квадраты (G-квартеты) складываются в стопки, и получаются довольно устойчивые, плотные структуры (G-квадруплексы). В четырехспиральные комплексы могут сплетаться четыре отдельные цепочки ДНК, но это скорее является исключением. Чаще единственная нить нуклеиновой кислоты просто завязывается в узел, образуя характерные утолщения (например, на концах хромосом), либо двуцепочечная ДНК на каком-то богатом гуанином участке образует локальный квадруплекс. Наиболее изучено существование квадруплексов на концах хромосом - на теломерах и в онкопромоторах. Однако до сих пор полное представление о локализации такой ДНК в человеческих хромосомах не известно. Все эти необычные структуры ДНК в линейной форме нестабильны по сравнению с В-формой ДНК. Однако ДНК часто существует в кольцевой форме топологического напряжения, когда у нее имеется так называемая сверхспирализация. В этих условиях легко образуются неканонические структуры ДНК: Z-формы, "кресты" и "шпильки", H-формы, гуаниновые квадруплексы и i-мотив. Формы ДНК
Все три перечисленные формы ДНК легко разделяются при гельэлекрофорезе. Третичная структура ДНК Третичная структура ДНК образуется в результате дополнительного скручивания в пространстве двуспиральной молекулы - ее суперспирализации. Суперспирализации молекулы ДНК в эукариотических клетках в отличие от прокариот осуществляется в форме комплексов с белками. ДНК эукариот почти вся находится в хромосомах ядер, лишь небольшое количество ее содержится в митохондриях, а у растений и в пластидах. Основное вещество хромосом эукариотических клеток (в том числе и хромосом человека) - это хроматин, состоящий из двухцепочечной ДНК, гистоновых и негистоновых белков. Гистоновые белки хроматина Гистоны - простые белки, составляют до 50% хроматина. Во всех изученных клетках животных и растений обнаружено пять основных классов гистонов: H1, H2A, H2B, H3, H4, различающихся по размерам, аминокислотному составу и величине заряда (всегда положительный).
Гистон Н1 млекопитающих состоит из одной полипептидной цепи, содержащей примерно 215 аминокислот; размеры других же гистонов варьируют от 100 до 135 аминокислот. Все они спирализованы и скручены в глобулу диаметром около 2,5 нм, содержат необычно большое количество положительно заряженных аминокислот лизина и аргинина. Гистоны могут быть ацетилированы, метилированы, фосфорилированы, поли(АДФ)-рибозилированы, а гистоны Н2А и Н2В – ковалентно связаны с убиквитином. Какова роль таких модификаций в становлении структуры и выполнении функций гистонами до конца пока не выяснено. Предполагается, что в этом заключается их способность взаимодействовать с ДНК и обеспечивать один из механизмов регуляции действия генов. Гистоны взаимодействуют с ДНК в основном через ионные связи (солевые мостики), образующиеся между отрицательно заряженными фосфатными группами ДНК и положительно заряженными лизиновыми и аргининовыми остатками гистонов. Негистоновые белки хроматина Негистоновые белки в отличие от гистонов очень разнообразны. Выделено до 590 разных фракций ДНК-связывающих негистоновых белков. Их еще называют кислыми белками, так как в их структуре преобладают кислые аминокислоты (они являются полианионами). С разнообразием негистоновых белков связывают специфическую регуляцию активности хроматина. Например ферменты, необходимые для репликации и экспрессии ДНК, могут связываться с хроматином временно. Другие белки, скажем, принимающие участие в различных процессах регуляции, связываются с ДНК только в специфических тканях или на определенных стадиях дифференциации. Каждый белок комплементарен определённой последовательности нуклеотидов ДНК (сайт ДНК). К этой группе относят:
При участии структурных, регуляторных белков и ферментов, участвующих в синтезе ДНК и РНК, нить нуклеосом преобразуется в высококонденсированный комплекс белков и ДНК. Образованная структура в 10 000 раз короче исходной молекулы ДНК.
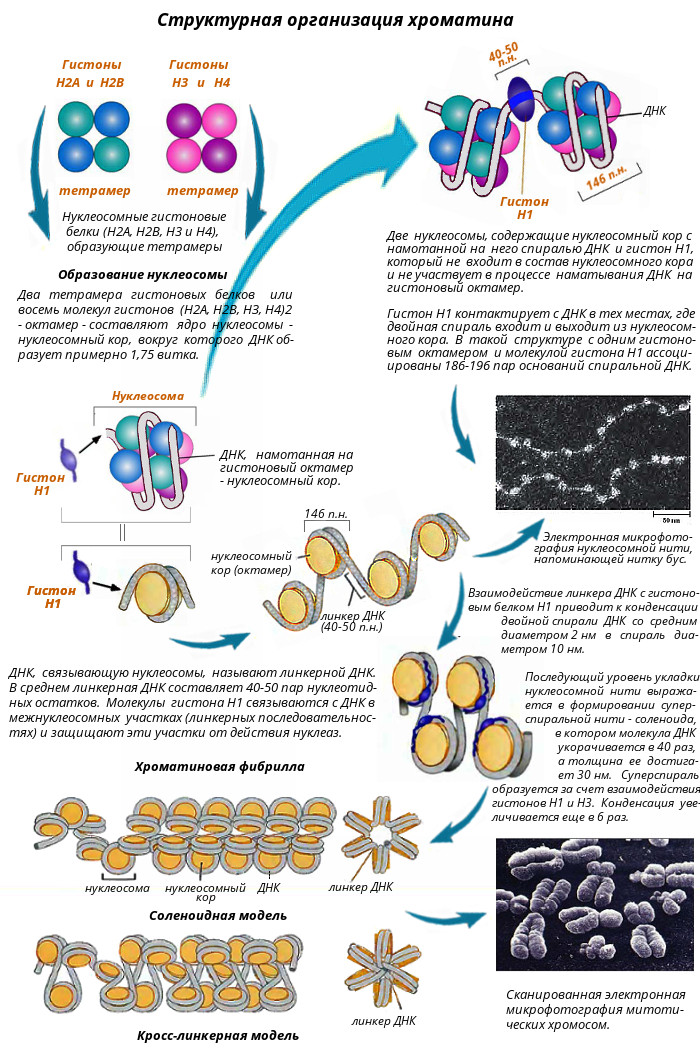 Хроматин Хроматин - это комплекс белков с ядерной ДНК и неорганическими веществами. Основная часть хроматина неактивна. Она содержит плотно упакованную, конденсированную ДНК. Это гетерохроматин. Различают конститутивный, генетически неактивный хроматин (сателлитная ДНК) состоящий из неэкспрессируемых областей, и факультативный - неактивный в ряду поколений, но при определенных обстоятельствах способный эспрессировать. Активный хроматин (эухроматин) неконденсированный, т.е. упакован менее плотно. В разных клетках его содержание составляет от 2 до 11%. В клетках головного мозга его больше всего - 10-11%, в клетках печени - 3-4 и почек - 2-3%. Отмечается активная транскрипция эухроматина. При этом его структурная организация позволяет использовать одну и ту же генетическую информацию ДНК, присущую данному виду организма, по-разному в специализированных клетках. В электронном микроскопе изображение хроматина напоминает бусы: шаровидные утолщения размером около 10 нм, разделенные нитевидными перемычками. Эти шаровидные утолщения названы нуклеосомами. Нуклеосома является структурной единицей хроматина. Каждая нуклеосома содержит сверхспиральный сегмент ДНК длиной 146 пар нуклеотидов, намотанный с образованием 1,75 левых витков на нуклеосомный кор. Нуклеосомный кор – это гистоновый октамер, состоящий из гистонов Н2А, Н2В, Н3 и Н4, по две молекулы каждого вида (рис. 9), который выглядит как диск диаметром 11 нм и толщиной 5,7 нм. Пятый гистон, Н1, не входит в состав нуклеосомного кора и не участвует в процессе наматывания ДНК на гистоновый октамер. Он контактирует с ДНК в тех местах, где двойная спираль входит и выходит из нуклеосомного кора. Это межкоровые (линкерные) участки ДНК, длина которых варьирует в зависимости от типа клеток от 40 до 50 нуклеотидных пар. В результате этого варьирует и длина фрагмента ДНК, входящего в состав нуклеосом (от 186 до 196 нуклеотидных пар). В состав нуклеосом входит примерно 90% ДНК, остальная ее часть приходится на линкер. Считается, что нуклеосомы - это фрагменты "молчащего" хроматина, а линкер - активного. Однако нуклеосомы могут развертываться и переходить в линейную форму. Развернутые нуклеосомы являются уже активным хроматином. Так наглядно проявляется зависимость функции от структуры. Можно считать, что чем больше хроматина находится в составе глобулярных нуклеосом, тем менее он активен. Очевидно, в разных клетках неодинаковая доля покоящегося хроматина связана с количеством таких нуклеосом. На электронно-микроскопических фотографиях в зависимости от условий выделения и степени растяжения хроматин может выглядеть не только как длинная нить с утолщениями – "бусинками" нуклеосом, но и как более короткая и более плотная фибрилла (волокно) диаметром 30 нм, образование которой наблюдается при взаимодействии гистона Н1, связанного с линкерным участком ДНК и гистона Н3, что приводит к дополнительному скручиванию спирали из шести нуклеосом на виток с образованием соленоида диаметром 30 нм. При этом гистоновый белок может препятствовать транскрипции ряда генов и таким образом регулировать их активность. В результате описанных выше взаимодействий ДНК с гистонами сегмент двойной спирали ДНК из 186 пар оснований со средним диаметром 2 нм и длиной 57 нм превращается в спираль диаметром 10 нм и длиной 5 нм. При последующем сжатии этой спирали до волокна диаметром 30 нм степень конденсации увеличивается еще в шесть раз. В конечном итоге упаковка дуплекса ДНК с пятью гистонами приводит к 50-кратной конденсации ДНК. Однако даже столь высокая степень конденсации не может объяснить почти 50 000 - 100 000-кратное уплотнение ДНК в метафазной хромосоме. К сожалению детали дальнейшей упаковки хроматина вплоть до метафазной хромосомы пока не известны, поэтому можно рассматривать лишь общие особенности этого процесса. Уровни компактизации ДНК в хромосомах Каждая молекула ДНК упакована в отдельную хромосому. В диплоидных клетках человека содержится 46 хромосом, которые располагаются в ядре клетки. Общая длина ДНК всех хромосом клетки составляет 1,74 м, однако диаметр ядра, в которое упакованы хромосомы, в миллионы раз меньше. Такая компактная укладка ДНК в хромосомах и хромосом в ядре клетки обеспечивается разнообразными, гистоновыми и негистоновыми белками, взаимодействующими в определенной последовательности с ДНК (см выше). Компактизация ДНК в хромосомах позволяет уменьшить ее линейные размеры примерно в 10 000 раз - условно с 5 см до 5 мкм. Выделяют несколько уровней компактизации (рис. 10). 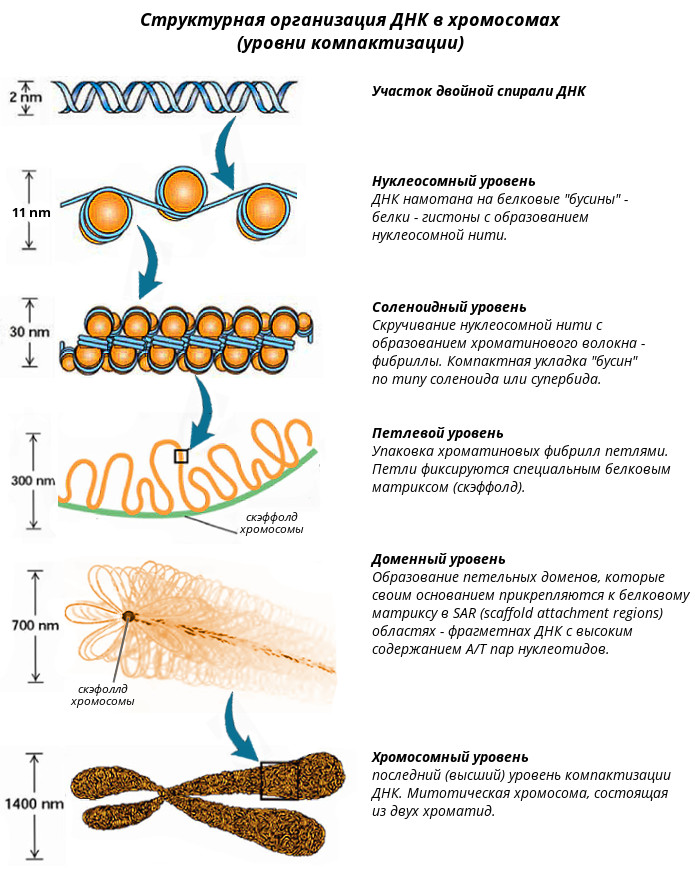 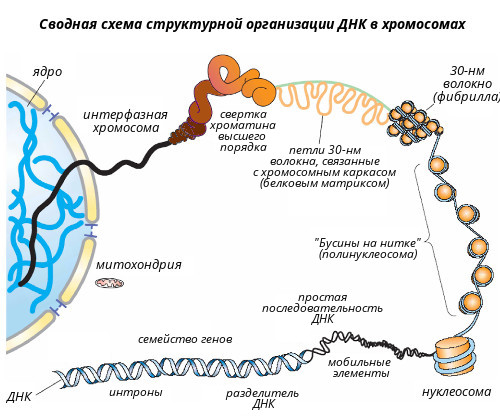
Нуклеосомный уровень компактизации обеспечивается специальными белками - гистонами. Восемь положительно заряженных гистоновых доменов образуют кор (сердцевину) нуклеосомы на которую наматывается отрицательно заряженная молекула ДНК. Это дает укорочение в 7 раз, при этом диаметр увеличивается с 2 до 11 нм.
Соленоидный уровень организации хромосом характеризуется скручиванием нуклеосомной нити и образованием из нее более толстых фибрилл 20-35 нм в диаметре - соленоидов или супербидов. Шаг соленоида равен 11 нм, на один виток приходится около 6-10 нуклеосом. Соленоидная упаковка считается наиболее вероятной, чем супербидная, согласно которой фибрилла хроматина диаметром 20-35 нм представляет собой цепь гранул, или супербидов, каждая из которых состоит из восьми нуклеосом. На соленоидном уровне линейный размер ДНК сокращается в 6-10 раз, диаметр увеличивается до 30 нм.
Петлевой уровень обеспечивается негистоновыми сайт-специфическими ДНК-связывающими белками, которые распознают определенные последовательности ДНК и связываются с ними, образуя петли примерно по 30-300 тысяч пар оснований. Петля обеспечивает экспрессию генов, т.е. петля является не только структурным, но и функциональным образованием. Укорочение на этом уровне происходит в 20-30 раз. Диаметр увеличивается до 300 нм. Петлеобразные структуры типа "ламповых щеток" в ооцитах земноводных можно видеть на цитологических препаратах. Эти петли, видимо, суперспирализованы и представляют собой домены ДНК, соответствующие, вероятно, единицам транскрипции и репликации хроматина. Специфические белки фиксируют основания петель и, возможно, некоторые их внутренние участки. Петлеобразная доменная организация способствует укладке хроматина в метафазных хромосомах в спиральные структуры более высоких порядков.
Доменный уровень организации хромосом изучен недостаточно. На данном уровне отмечается образование петлевых доменов - структур из нитей (фибрилл) толщиной 25-30 нм, которые содержат 60% белка, 35% ДНК и 5% РНК, практически не видны во всех фазах клеточного цикла за исключением митоза и несколько беспорядочно распределены по клеточному ядру. Петлеобразные структуры типа "ламповых щеток" в ооцитах земноводных можно видеть на цитологических препаратах. Петлевые домены своим основанием прикрепляются к внутриядерному белковому матриксу в так называемых встроенных местах прикрепления, часто обозначаемых как MAR/SAR-последовательности (MAR, от англ. matrix associated region; SAR, от англ. scaffold attachment regions) – фрагментах ДНК протяженностью в несколько сотен пар оснований, которые характеризуются высоким содержанием (>65%) А/Т пар нуклеотидов. Каждый домен, по-видимому, имеет одну точку начала репликации и функционирует как автономная сверхспиральная единица. Любой петельный домен содержит множество единиц транскрипции, функционирование которых, вероятно, координируется – весь домен находиться либо в активном, либо в неактивном состоянии. На доменном уровне в результате последовательной упаковки хроматина присходит уменьшение линейных размеров ДНК примерно в 200 раз (700 нм).
На хромосомном уровне происходит конденсация профазной хромосомы в метафазную с уплотнением петельных доменов вокруг осевого каркаса негистоновых белков. Эта суперспирализация сопровождается фосфорилированием в клетке всех молекул H1. В результате метафазную хромосому можно изобразить в виде плотно уложенных соленоидных петель, свернутых в тугую спираль. Типичная хромосома человека может содержать до 2600 петель. Толщина такой структуры достигает 1400 нм (две хроматиды), а молекула ДНК при этом укорачивается в 104 раз, т.е. с 5 см растянутой ДНК до 5 мкм. Функции хромосом Во взаимодействии с внехромосомными механизмами хромосомы обеспечивают
Существуют данные, что при активировании участка хроматина, т.е. при транскрипции, с него обратимо удаляются сначала гистон H1, а затем и октет гистонов. Это вызывает деконденсацию хроматина, последовательный переход 30-нанометровой фибриллы хроматина в 10-нанометровую нить и ее дальнейшее разворачивание в участки свободной ДНК, т.е. утрату нуклеосомной структуры. Дополнительно: Решетников Р. В., Копылов А. М., Головин А. В. Классификация G-квадруплексных ДНК по углу вращения квадруплекса и планарности G-квартетов. Журнал Acta Naturae (русскоязычная версия), № 4, том 2, 2010 Иванов В.И. А-ДНК. Биология. Соросовский образовательный журнал, N1, 1998 Тодоров И.Н. Митохондрии: окислительный стресс и мутации митохондриальной ДНК в развитии патологий, процессе старения и апоптозе. Ж.Рос.хим.об-ва им.Д.И.Менделеева, т. LI, N 1, 2007 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||