Типы ареалов и их классификация Природный фактор формирования ареала
 Скачать 280.44 Kb. Скачать 280.44 Kb.
|

|
Оглавление Введение Типы ареалов и их классификация Природный фактор формирования ареала Заключение Библиографический список Приложения Введение Распределение растений по земной поверхности зависит от биологических особенностей и экологических свойств растений, большей или меньшей способности их к размножению, расселению и приспособляемости их к внешним условиям. Зависит оно также от тех преград, которые растения встречают со стороны различных форм земной поверхности – гор, морей, климата, других живых организмов и человека. Так как все эти факторы не остаются неизменными, а непрерывно меняются, то и распределение видов растений не является застывшим, постоянным, а также подвергается непрерывным изменениям, хотя и медленным, пока в этот процесс не вмешивается человек. Актуальность: Изучение ареалов имеет большое теоретическое и практическое значение. Изучая распространение растений на земном шаре, учёные установили, что различные материки имеют много общих видов, родов и семейств. Это сходство флор удалённых друг от друга территорий объясняется историческими причинами. Изучая ареалы видов, родов, семейств, можно более точно установить родственные связи между ними, проследить процессы формообразования, а также установить участки суши, имеющие один возраст. Цель исследования: Изучить типы ареалов и факторы формирования ареалов. Для достижения поставленной цели нами предусматривались следующие задачи: Рассмотреть типы ареалов и их классификацию; Изучить природные факторы формирования ареалов; Изучить антропогенные факторы формирования ареалов. Типы ареалов и их классификация Изучая ареалы различных видов, родов и других групп растений, мы убеждаемся, что при всем многообразии размеров и очертаний этих ареалов многие их особенности в какой-то степени повторяются. Ареалы растений, принадлежащих к самым различным систематическим группам, могут в большей или меньшей степени уподобиться друг другу. Мы можем, например, отметить весьма значительное сходство ареалов Линнея северной (Linnaea borealis) и Седмичника европейского (Trientalis europaea). Бывает и так, что ареал отдельного вида определенного рода подобен ареалу целой группы видового рода. Например, значительное сходство можно заметить, сравнивая ареалы Подмаренник душистый (Asperula odorata) и группы видов лип, тяготеющей к Липе сердцевидной (Tilia cordata). Внимательно рассматривая соответствующие карты, мы заметим, однако, и определенные различия в распространение растений, ареалы которых нами сравниваются. Как правило, мы имеем дело не с полным совпадением очертаний различных ареалов, но лишь с более или менее значительным сходством. Сходство это подчас настолько велико, что мысль о его обусловленности какими-то общими причинами возникает сама собой. Довольно часто мы замечаем, что, по крайне мере, в некоторой части расположение границ ареалов практически совпадает, так что при нанесении на карту целой серии ареалов границы их сливаются, хотя в других частях очертаний тех же ареалов могут быть различны. Наличие всех отмеченных соотношений подсказывает мысль о плодотворности сравнительного изучения ареалов и известного упорядочения данных о сходствах и различиях в их очертаниях и путем соответствующей классификации ареалов. Последняя должна способствовать ориентировке в многообразии ареалов, облегчая выяснение закономерностей их развития. Упоминая о закономерностях развития ареалов, мы должны подчеркнуть своё принципиальное несогласие со стремлением некоторых исследователей противопоставить сравнительное изучение ареалов стремлением к выяснению причиной обусловленности распространения растений и, в частности, очертаний ареалов, т. е. к объяснению тех соотношений, которые выявляются при непосредственно изучения ареалов как природного явления. Напротив, сравнительное изучение ареалов потому и важно, что оно не только упрощает рассматривание многообразных явлений, но и помогает обнаружению закономерностей распространений растений, обусловленных разным ходом развития ареалов, отражающих взаимодействие на него различных факторов, т. е. помогает разобраться в том, почему определённые растения распространены так, а не иначе. Понятие о типах распространения растений имеет преимущественно значение при рассмотрении не общих закономерностей распределения их по поверхности земного шара как целого, а при анализе флор отдельно взятых его частей. Соответственно этому сравнительное изучение ареалов и построение их классификаций должно преследовать различные частные цеди в зависимости от специальных задач, ставящихся тем иди иным ботанико-географическим исследованием, и от специфики подвергаемого изучению флористического материала. В этом смысле оно должно носить в известной степени прикладной характер. Исключительно многообразно распространение растений, общих Европе и восточным частям Северной Америки, по отсутствующих на западе последней и на северо-востоке Азии. Превосходная сводка Хультена дает яркое представление об ареалах этих амфиатлантических видов. Но, присматриваясь к их ареалам, мы убеждаемся, что единого амфиатлантического типа распространении не существует, что мы имеем перед собой разнотипные ареалы, обладающие определенны ми чертами сходства лишь в одном определенном отношении (наличие географической «связи» между европейской и американской частями ареалов через посредство северной части Атлантического океана). Многообразны (и не только в деталях, но к в существеннейших крупных чертах) ареалы растений, объединяемых в сборную группу арктоальпийских. Они по-разному распространены в Арктике, будучи 70 циркумполярными, то приуроченными к какой-то определенной части околополярного кольца суши. Одни из них общи Арктике с горами Средней Европы, другие сочетают произрастание в Арктике с таковым в горах юга Сибири и Центральной Азии. Широкое распространение в Арктике может сочетаться с узколокализованным в относительно южных горах. У других видов соотношения могут быть противоположными. Попытка классифицировать все это многообразие ареалов в общеземном плане неизбежно поставила бы нас перед вопросом, какие из объединяющих особенностей и из различий в распространении изучаемых видов должны быть отражены классификацией в первую очередь, преимущественно перед другими. Решать этот вопрос в «общетеоретической» постановке едва ли возможно. Но он вполне разрешим (только по-разному!) при анализе флор определенных областей, ставящим перед особо совершенно определенные цели – различные в зависимости от того, какая именно флора изучается и какие конкретные вопросы ее сложения, истории развития должны быть решены. Типы ареалов: 1. Простой ареал имеет сплошную территорию. Установление границ простых ареалов не составляет особых трудностей, если организмы лишены возможности активно перемещаться (растения), либо ведут оседлый образ жизни (некоторые животные). 2. Сложный ареал имеет разорванную территорию. Для птиц это может быть область размножения, зимовок, вместе с миграционным путем. Среди животных много видов, которые в различные периоды жизненного цикла или сезоны меняют область распространения (птицы, млекопитающие). 3. Сплошной – это ареал, все участки которого доступны для особей вида, занимающего его. Перемещение особей с одного участка на другой в пределах сплошного ареала осуществляется при помощи естественных факторов расселения и не носит случайный характер. Внутри сплошного ареала нет непреодолимых географических барьеров, разделяющих его на отдельные участки. Все простые ареалы являются сплошными. Следует отметить, что ареалы также бывают первичные и вторичные, причем первичный ареал, в котором происходит становление вида, является сплошным. 4. Разорванный ареал (дизъюнктивный) – возникает при изменении условий существования вида: появляются горные системы, изменяется климат. а) Гомогенная (однородная) дизъюнкция, когда не граничащие друг с другом участки заселены одной и той же формой вида (русская выхухоль). б) Гетерогенная (разнородная) дизъюнкция, когда разорванные участки заселены разными подвидами одного вида, разными видами одного рода (дуб скальный, дуб монгольский). Или разными родами одного семейства. Гомогенные разрывы ареала в возрастном отношении моложе гетерогенных. 5. Выделяются типы ареалов в зависимости от занимаемой площади. Площадь ареала может быть сопоставима с площадью почти всей суши или ограничиваться небольшим участком. В зависимости от размеров занимаемой территории выделяют различные ареалы узколокальные, локальные, субрегиональные, региональные, полирегиональные, космополитические. а) Узколокальные – характерны для видов, имеющих крайне ограниченную площадь распространения. б) Локальные – по площади несколько больше узколокальных. в) Субрегиональные – больше локальных. г) Региональные – больше субрегиональных. Если вид распространен по всей территории региона, его считают трансрегиональным. Под регионом в данном случае понимают определенную крупную территориальную единицу, как правило на одном материке. д) Полирегиональные – более широкие ареалы, включающие регионы, расположенные на нескольких, но не более чем на трех материках. К полирегиональным относятся ареалы следующих представителей флоры Евразии: евроазиатские виды в пределах умеренной зоны Евразии; евро-сибирские виды, встречающиеся по всей Европе, в западной и Восточной Сибири до Дальнего Востока. Для наименования полирегионального ареала используют названия регионов, в которых обитает вид, род и т.д. е) Космополитические ареалы – характерны для таксономических категорий, обитающих не менее чем на трех материках. Они свойственны обычно водным и болотным растениям, сорным травам. Космополитические ареалы часто ограничиваются широтными пределами и присущи организмам, распространяющимся в определенных физико-географических поясах или природных зонах. Дополнительно космополитические ареалы делят на несколько типов: Циркумконтинентальные ареалы встречаются на суше (циркумумеренные, циркумбореальные). Циркумокеанические ареалы встречаются на океанических и морских акваториях (планктон). Циркумполярные ареалы на суше охватывают зоны арктических пустынь, тундр, лесотундр (песец). Биполярные ареалы характерны для организмов циркумполярных областей в Северных и Южных полушариях. Некоторым организмам свойственны биполярные разрывы ареалов, когда они присутствуют в умеренных холодных широтах обоих полушарий, но отсутствуют в приполярных областях (виды сфагнумов, мхов, буков). Биполярное распространение растений возможно связано с переносом семян и спор перелетными птицами на большие расстояния. Не исключена миграция в Южное полушарие представителей бореальной флоры по горным системам во время четвертичного оледенения, когда снеговая линия в тропических широтах была ниже современной. Природный фактор формирования ареала Формирование ареала связанно с образованием вида и завоеванием им пространства. Новые виды образуются путем изменчивости предковых форм. Естественного обора и сохранения наиболее приспособленных, победивших в борьбе за существования. Процесс увеличения биоразнообразия опирается на теорию монофилитической эволюции Ч. Дарвина. Согласно этой теории многообразие видов в крупной таксономической группе растений или животных происходит от одного корня в результате дивергенции – расхождения признаков. На первых этапах дивергенции осуществляется в рамках микроэволюционного процесса, т.е. изменчивости существующих организационных признаков. Она носит ярко выраженный приспособительный (адаптационный) характер. Виды, попавшие в условия географической изоляции, дают начало ветвям филогенетического древа, все дальше отходящими от основного ствола сначала на уровне микро-, а затем и макроэволюционной дивергенции. Таким образом формируются неповторимые черты биот, отличающиеся не только по составу, но и родов, и семейств. Каждый вид существует определенное время, его популяция растет, увеличивается площадь распространения. Заем в силу разнообразных причин происходит вымирание особей отдельных видов, их ареалы сокращаются, и наконец, они исчезают. В одних случаях исчезновение вида не прерывает хода эволюции, старый вид вытесняется новым; в других – гибель вида обозначает слепую ветвь филогенетического древа. Процесс формирования нового ареала, как правило, возникновение вида происходит в одном месте, откуда он начинает завоевывать пространство. Новый вид происходит от одного предка или в результате гибридизации. Эти положения являются ключевыми для понимания истории формирования ареала. Расселение организмов способствуют различные приспособления их зачатков (диаспор): при половом размножении растений – спор, плодов, семян, при вегетативном – луковиц, клубней, коровищ, ползучих побегов и т.д. Поды и семена большинства водных, а равно и значительного количества наземных растений его переноситься текшими водами, чем облегчаются расселения растений вниз по течению рек. Многие плоды и семена могут переноситься морскими течениями. Например, ареал кокосовой пальмы (Cocos nucifera) охватывает берега тропических островов, разбросанных акваториям Тихого океана и Индийского океанов. Кокосовые орехи, попадая в воду, разносятся морскими течениями. Они хорошо приспособлены к длительным и морским путешествиям. Своей плавучести они обязаны заполненной воздухом и непроницаемой для соленой морской воды волокнистой наружной оболочке. Орехи не теряют способности к прорастанию после плавания в море в течение 3–4 месяцев. За это время они могут быть перенесены на расстояние 5000 км. Легкость спор и мелких семян множества видов обеспечивает возможность переноса их на более или менее значительное расстояние даже слабыми токами воздуха. У растений с более тяжелыми семенами возможность переноса по воздуху часов возрастает благодаря «летательным приспособлением» – волосистыми придатками, разного вида хохолками, перепончатыми лопастями. Перенос зачатков растений при помощи ветра может осуществляться не только по воздуху: опавшие на землю плоды и семена, при наличии у них летучек, могут перегоняться ветром по поверхности почвы; семена деревьев, опавшие зимой на затвердевшую поверхность снежного покрова, перегоняются, скользя по ней, особенно легко. Упав на поверхность воды, снабженные летучками смена, долго ре тонут и под напором ветра легко передвигаются по поверхности водоемов. Очень многие виды настолько тесно связанны с определением биоценотическими условиями, что их расселение возможно лишь там, где складываются совершенно определенные соотношения между другими растениями. Так, многие растения темнохвойных, таежных лесов могут расселяться только при наличии древесного полога определенного характера, а, следовательно, расширять свой ареал только за счет пространств, уже заселенных или заселяемых определенными видами деревьев. Такие растения умеренного севра, как встречающиеся у нас оба вида клюквы (Oxycoccus quadripetalus), подбел (Andromeda polifolia), росянки (Drosera rotundifolia), настолько тесно связаны с условиями сфагновых болот, что развитие последних, поселение в данной местности определенных видов мхов из рода Sphagnum является необходимой предпосылкой их расселения. Поскольку подобных, тесно связанных с определенной биоценотической средой растений во многих странах значительное количество и целые группы видов требуют для своего расселения одних и тех же предпосылок, у нас иногда создается впечатление, что происходит переселение целой формации, биоценоза. Расселение видов может регулироваться и более узкими биологическими связями между ними. Паразитное растение, например, зависит от условий, создаваемых присутствием или отсутствием па определенном пространстве того растения-хозяина, с которым паразит связан. Некоторые растения в своем распространении от определенных видов животных, являющихся переносчиками их семян. Обязательной предпосылкой расселения других растений является предшествующее или одновременное расселение насекомых, опыляющих их цветы. Расселение растений, как всякий естественный процесс, требует для своего осуществления некоторого времени. Поэтому при прочих равных условиях широта распространения вида может служить отражением его возраста, или точнее времени, которым он располагал для расселения. Это положение может применяться и в частной форме – для суждения о времени присутствия и расселения вида в какой-то определенной стране. Учет изменяемости условий, определяющих возможности расселения растений, позволяет в ряде случаев устанавливать, к какому времени было приурочено расселение тех пли иных видов, каковы могли быть конкретные условия того пространства, через которое шло их расселение в соответствующее время. С тех пор как первые экспедиции на Огненную Землю и Фолклендские о-ва — Коммерсонеа, Дюмон дТОрвиля и в особенности экспедиция Росса, в которой участвовал Гукер, доставили сведения о произрастании на них арктических и субарктических растений, проблема биполярных ареалов служила предметом неоднократного рассмотрения. Загадочность этого разрыва ареала, в промежутке между частями которого расположена во всю ширину неотропическая зона, в пределах которой эти растения обитать не могут, привлекала внимание многочисленных исследователей, начиная от Дарвина и Гукера. Еще в 1939 г. Стеффеном опубликована (Steffen, 39) новейшая сводка по этому вопросу, которой мы в дальнейшем и воспользуемся. С самого научала необходимо только оговориться, что в этом разделе мы будем рассматривать не биполярные ареалы семейств и таких родов, как Ranupculus, Caltha, Saxifraga, Draba, Lupinus, Gentiana, Veronica, Euphrasia, Pedicularis, Plantago, Valeriana, Senecio и др. Свое: образность эндемичных циклов и форм, которые эти роды образовали в южном полушарии, говорит о давнем, во всяком случае не позднее чем с третичного периода, нахождении их в этих современных обитаниях. Этот разрыв ареалов является результатом широкого распространения родов, еще при своем возникновении мало дифференцированных, в то время, когда материки были связаны в одно целое. В отношении таких родов циркумполярное распространение ничего загадочного не представляет. На соображениях о нахождении этих родов в субантарктической зоне мы остановимся позднее. Сейчас же нашей задачей является объяснение нахождения в южном полушарии тождественных видов или небольших в систематическом отношении уклонений от арктических или субарктических видов. Некоторые систематики считают такие уклоненные формы за. разновидности, другие, основываясь не на морфологических отличиях, а на географической удаленности их, выделяют их в самостоятельные виды. Это дело вкуса и неурегулированности понятия о виде и низших систематических единицах. Во всяком случае, как бы мы их ни называли, несомненно, что степень их уклонений от бореальных видов настолько незначительна, что недавний характер разрыва их ареалов не может вызвать какие-либо сомнения. Мы оставляем в стороне биполярные виды мхов и папоротникообразных, время происхождения которых настолько от нас отдалено, что мы имеем очень мало данных для суждения о происхождении их современного ареала. Также отбрасываем и ряд видов заносного характера и космополитов с очень широкой амплитудой приспособляемости, а остановимся только на тех видах, биполярный разрыв ареалов которых может считаться действительно загадочным. Таких видов[1], согласно Стеффену, 16. Все они являются аркто-аль- пийцами или субаркто-ореофитами. Иначе говоря, мы имеем здесь дело не с чисто арктическими видами, а с ореофитамц, растущими и в альпийских поясах горных хребтов. Это весьма важнее обстоятельство, облегчающее объяснение биполярности этих ареалов. Объяснения, которые давались этим биполярным ареалам, были троякого характера. Одни искали их в заносе этих растений человеком, птицами, ветром, морскими течениями (Grisebaeh, 56). Другие считали, что они являются результатом распространения этих видов по торным системам в четвертичном периоде, когда общее охлаждение климата могло способствовать таким миграциям. Наконец, третьи пытались в этих видах найти доказательство полигонного происхождения видов. В отношении указанных 16 видов нет никакого основания предполагать занос их человеком. Образование многими из них эндемичных разновидностей или превращение их в викарные виды исключает возможность их столь недавнего заноса. Случайные же факторы, как ветер, птицы, морские течения, при переносе диаспор на большие расстояния — а в данном случае речь идет о 8000—9000 км — нельзя серьезно принимать во внимание, как мы это неоднократно старались обосновать (см. ч. I). От этих ничем не подтверждаемых доказательств уже давно пора совсем отказаться точно так же как и от теории политоп- ного происхождений (см. ч. I), не выдерживающей никакой критики. Остается, следовательно, только одно возможное объяснение — это постепенное распространение этих видов по горным системам. Такое объяснение не представляет особых затруднений, в особенности если учесть, как это делает Стеффен (1. с.), перемещение экваториальной зоны, вытекающее из теории перемещения материков Вегенера. Но сейчас же возникает вопрос, в каком направлении шло распространение этих биполярных видов, т. е., иначе говоря, в южном или северном полушарии находятся центры их происхождения. На этот вопрос надо ответить, что оно могло итти в обоих направлениях, но не всегда мы сейчас можем точно установить, откуда — с севера или с юга — шла миграция вида. Примером может служить род Empetrum, представленный в субантарктической Америке видом Е. rubrum, а в арктической и северной умеренных зонах — циркумполярным Е. nigrum. Но, помимо этих двух видов, в Ньюфаундленде, вокруг залива св. Лаврентия, растут еще два описанных Фернальдом вида —E. atropurpureum и E. Eamesii, более близкие к южному Е. rubrum, чем к северному Е. nigrum (Fernald, 25). Согласно Стеффену, несомненно южное антарктическое происхождение имеет Careix microglochin. Этот вид образует промежуточное звено между родом Сагех и распространенным в южном полушарии родом Uncinia. Вместе с тем морфологические признаки, отличающие эти два рода, не дают оснований для того, чтобы вывести Uncinia из Сагех, а скорее наоборот. Но несомненно, что большая часть видов Сагех имеет северное происхождение. Необходимая предпосылка для доказательства возможности этих миграций — существование альпийского пояса горной цепи — в данном случае имеется, так как Анды уже в олигоцене достигали высоты, превышающей границу распространения леса. Распространению этих полярных видов не могло не способствовать перемещение экватора. Экватор в, эоцене на долготе Америки должен был быть расположен на 30°, а в олигоцене — на 25° севернее современного положения, когда Колумбия находилась уже вне тропической зоны. Следовательно, в это время антарктические виды могли беспрепятственно мигрировать по цепи Анд почти до самого севера Ю. Америки. В течение всего третичного периода шло перемещение экватора к югу, причем в начале четвертичного периода он оказался на 20° южнее, чем в данное время. Это обстоятельство и связанное с этим охлаждение климата во всей лежащей к северу от тогдашней экваториальной зоны области явилось, в свою очередь, благоприятным для миграции арктических видов на юг. Возможность этого позднего распространения арктических видов в пределы южного полушария объясняет сохранение их в большем количестве в южном полушарии и малую их измененность. При перемещении экватора с севера на юг, распространившиеся в третичном периоде на север антарктические виды должны были оказаться в неблагоприятных условиях. Тем не менее можно предположить, что часть их сохранилась в верхнем поясе Кордильер, оставшемся если не повсеместно, то частично вне границы леса. В начале четвертичного периода эти виды по хребту Скалистых гор достигли арктической Америки, откуда распространились на запад до гор Азии и Европы. Мигрировавшие с севера виды находились по отношению к перемещавшейся экваториальной зоне в более благоприятных условиях. Их прохождение через нее должно было произойти в четвертичном периоде, когда Анды уже почти достигли современной высоты, давшей возможность этим арктическим видам пересечь тропическую зону при их распространении на юг. Оледенение Анд в ледниковое время, частично сохранившееся до настоящего времени, привело к вымиранию большинства этих видов в промежуточных местообитаниях, в результате чего мы и имеем сейчас такие разорванные биполярные ареалы. Единственное затруднение для объяснения изложенных миграций может представить только Панамский перешеек, имеющий сейчас возвышенности, далеко не достигающие 1000 м и поэтому доверху покрытые лесом. Этот промежуток между северной оконечностью Анд, где они доходят до 3000 м высоты, и Кордильерами де Сантьяго в Центральной Америке составляет около 600 км. Имели ли Панамские горы в первой половине четвертичного периода ту же высоту, что и сейчас, или они снижены эрозией, доходил ли лес во время ледникового периода до самых их вершин, как сейчас, или существовал безлесный пояс или безлесные участки, давшие возможность рассматриваемым видам преодолеть это препятствие на своем пути, — этого мы сейчас сказать не можем. Но что означенные миграции действительно осуществлялись, мы, к счастью, имеем два свидетельства. Одним из них является то обстоятельство, что не все виды вымерли в промежуточных местонахождениях между арктической и антарктической частями своего ареала. Ряд видов[2] сохранился в изолированных местообитаниях в Андах и Скалистых горах. Второе же свидетельство о существовании таких миграций — это виды[3], в своем распространении достигшие Анд, но, очевидно, не дошедшие до Патагонии и Огненной Земли. . Заключение Обычно выделяют простые, сложные, сплошные и дизъюнктивные ареалы. Сплошной ареал характеризуется сплошным, без пропусков покрытием видом всей населяемой площади, на которой заняты все пригодные для жизни вида местообитания. Примером сплошных ареалов служат области распространения ели сибирской, линнеи северной и т. д. Ареал, состоящий из двух или нескольких частей, разобщенных друг с другом называется дизъюнктивным (разорванным). Дизъюнктивные ареалы образуются из некогда сплошных ареалов. Дизъюнктивные ареалы могут возникать, когда живые организмы сумели преодолеть ту или иную преграду. Во второй главе мы рассмотрели факторы формирования ареалов: природный и антропогенные факторы. Процессы расселения растений осуществляется в природе по-разному не только в зависимости от разнохарактерных биологических особенностей различных видов, в частности от их средств расселения, но и от внешних условий, благоприятствующих реализации имеющихся у вида возможностей или затрудняющих ее. Огромное влияние на формирование ареалов растений и изменения их границ оказывает хозяйственная деятельность человека. Вырубая леса, распахивая степи, осушая болота, человек уничтожает местообитания тех или иных видов, что приводит к сокращению их ареала. Но, с другой стороны, деятельность человека способствует расширению ареалов некоторых видов растений. По итогам работы следует отметить, что ареал вида представляет собой сочетание видовых требований с определённой суммой экологических условий на обширной территории или акватории. Однако эта эколого-географическая сопряжённость видов осложняется изменениями физико-географических условий в прошлом – климата, растительного покрова, формы земной поверхности и пр. Поэтому ареал вида является суммированным эффектом современных и предшествовавших условий. В результате, в географическом распределении видов растений наблюдается ряд неравномерностей и свои закономерности. Исходя из проделанной работы, можно прийти к выводу, что цель и задачи, поставленные в данной работе, были выполнены. Обоснована необходимость изучение типов ареала и их классификации. Библиографический список Абдурахманов Г.М., Лопатин И.К., Исмаилов Ш.И. Основы зоологии и зоогеографии. - М.: Академия, 2001. - 496 с. Вавилов Н.И. Центры происхождения культурных растений / Пять континентов. - М.: Мысль, 1987. - 348 с. Вальтер Г. Общая геоботаника. - М.: Мир, 1982. - 255 с. Вернадский В.И. Биосфера. - Л., 1926. - 146 с. Воронов А.Г. Биогеография. / А.Г. Воронов – М.: МГУ. 1987. – 264 с. Воронов А.Г., Дроздов Н.Н., Криволуцкий Д.А., Мяло Е.Г. Биогеография с основами экологии. - М.: Изд-во МГУ, 1999. - 392 с. Воронов А.Г., Дроздов Н.Н., Мяло Е.Г. Биогеография мира. - М.: Высшая школа, 1985. - 271 с. Второв П.П., Дроздов Н.Н. Биогеография. - М.: Изд-во ВЛАДОС-ПРЕСС, 2001.-304 с. Второв П.П. Биогеография материков. / П.П. Второв, Н.Н. Дроздов – М.: Просвещение. 1979. – 320 с. Гордеева Т.Н. Практический курс географии растений / Т.Н. Гордеева, О.С. Стрелкова. — М.: Высш. шк., 1968. — 335 с. Приложения Приложение 1 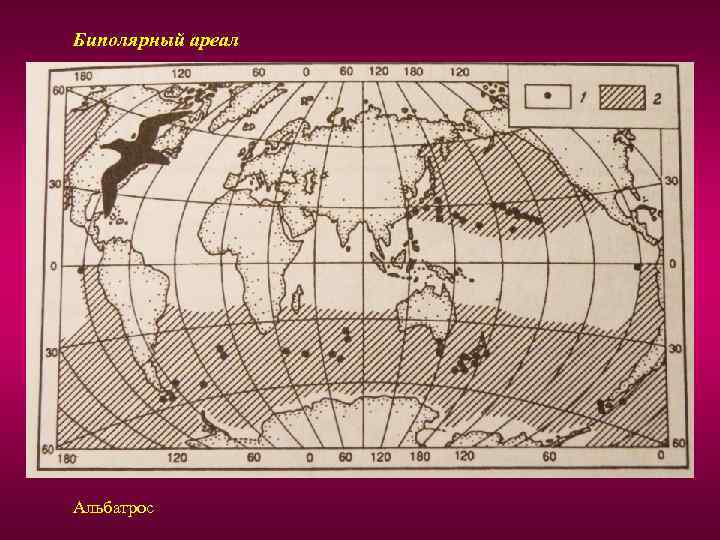 Приложение 2 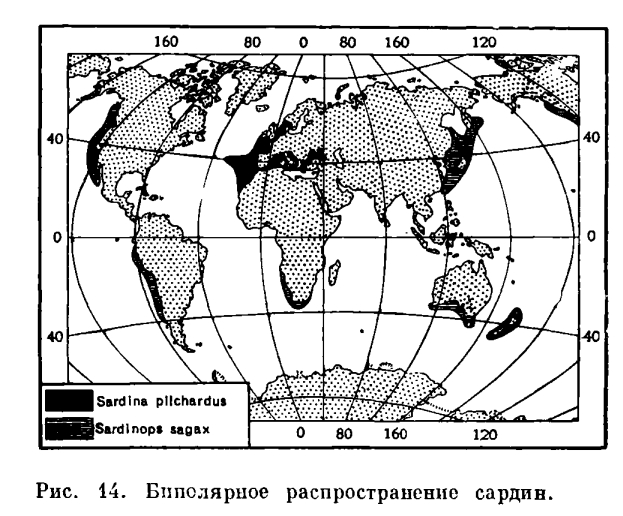 |