|
Бескаравайная, вопросы 19-24. Типы наследования признаков независимое, сцепленное, аутосомное, сцепленное с полом, голандрическое, моногенное, полигенное. Примеры
Типы наследования признаков — независимое, сцепленное, аутосомное, сцепленное с полом, голандрическое, моногенное, полигенное. Примеры.
Наследование — передача генетической информации (генетических признаков) от одного поколения организмов к другому
Независимое наследование - наследование определенного гена (признака) без влияния иных генетических факторов (др. определенного гена, пола); как правило, говорят о н.н. генов, входящих в разные группы сцепления. (закон Г. Менделя: при дигибридном скрещивании у гибридов второго поколения каждая пара контрастных признаков наследуется независимо от других и дает расщепление 3:1, образуя при этом четыре фенотипические группы в соотношении 9:3:3:1.)
Сцепленное наследование — наследование признаков, гены которых локализованы в одной хромосоме. Сила сцепления между генами зависит от расстояния между ними: чем дальше гены располагаются друг от друга, тем выше частота кроссинговера и наоборот. а) Полное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным. б) Неполное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними.
Моногеннымназывается такой тип наследования, когда наследственный признак контролируется одним геном. (Аутосомное наследование и сцепленное с полом наследование)
Аутосомное наследование — независимое от пола (не сцепленное с полом) наследование какого-либо признака. Характерные черты аутосомного наследования признаков обусловлены тем, что соответствующие гены, расположенные в аутосомах, представлены у всех особей вида в двойном наборе. Это означает, что любой организм получает такие гены от обоих родителей. В соответствии с законом чистоты гамет в ходе гаметогенеза все половые клетки получают по одному гену из каждой аллельной пары. Обоснованием этого закона является расхождение гомологичных хромосом, в которых располагаются аллельные гены, к разным полюсам клетки в анафазе I мейоза.
Наследование, сцепленное с полом — наследование какого-либо гена, находящегося в половых хромосомах. (гемофилия, дальтонизм)
а) Х-сцепленное наследование. Х-хромосома присутствует в кариотипе каждой особи, поэтому признаки, определяемые генами этой хромосомы, формируются у представителей как женского, так и мужского пола. Особи гомогаметного пола получают эти гены от обоих родителей и через свои гаметы передают их всем потомкам. Представители гетерогаметного пола получают единственную Х-хромосому от гомогаметного родителя и передают ее своему гомогаметному потомству.
Голандрическое наследование. Активно функционирующие гены Y-хромосомы, не имеющие аллелей в Х-хромосоме, присутствуют в генотипе только гетерогаметного пола, причем в гемизиготном состоянии. Поэтому они проявляются фенотипически и передаются из поколения в поколение лишь у представителей гетерогаметного пола. Так, у человека признак гипертрихоза ушной раковины («волосатые уши») наблюдается исключительно у мужчин и наследуется от отца к сыну.
Полигенное наследование - тип наследования признаков, обусловленных действием многих генов, каждый из которых оказывает лишь слабое действие. Фенотипически проявление полигенно обусловленного признака зависит от условий внешней среды. У потомков наблюдается непрерывный ряд вариаций количественного проявления подобного признака, а не появление четко различающихся по фенотипу классов. В ряде случаев при блокировании отдельного гена признак не проявляется вообще, несмотря на его полигенную обусловленность. Это свидетельствует о пороговом проявлении признака.
Законы Г. Менделя. Цитологические основы универсальности законов Г. Менделя. Менделирующие признаки человека. Примеры. Промежуточный тип наследования. Анализирующее скрещивание и его значение.
Законы Г. Менделя.
Закон единообразия гибридов первого поколения (первый закон Менделя) — при скрещивании двух гомозиготных организмов, относящихся к разным чистым линиям и отличающихся друг от друга по одной паре альтернативных проявлений признака, всё первое поколение гибридов (F1) окажется единообразным и будет нести проявление признака одного из родителей.
Закон расщепления (второй закон Менделя) — при скрещивании двух гетерозиготных потомков первого поколения между собой во втором поколении наблюдается расщепление в определенном числовом отношении: по фенотипу 3:1, по генотипу 1:2:1.
Закон независимого наследования (третий закон Менделя) — при скрещивании двух особей, отличающихся друг от друга по двум (и более) парам альтернативных признаков, гены и соответствующие им признаки наследуются независимо друг от друга и комбинируются во всех возможных сочетаниях (как и при моногибридном скрещивании).
Цитологические основы универсальности законов Г. Менделя.
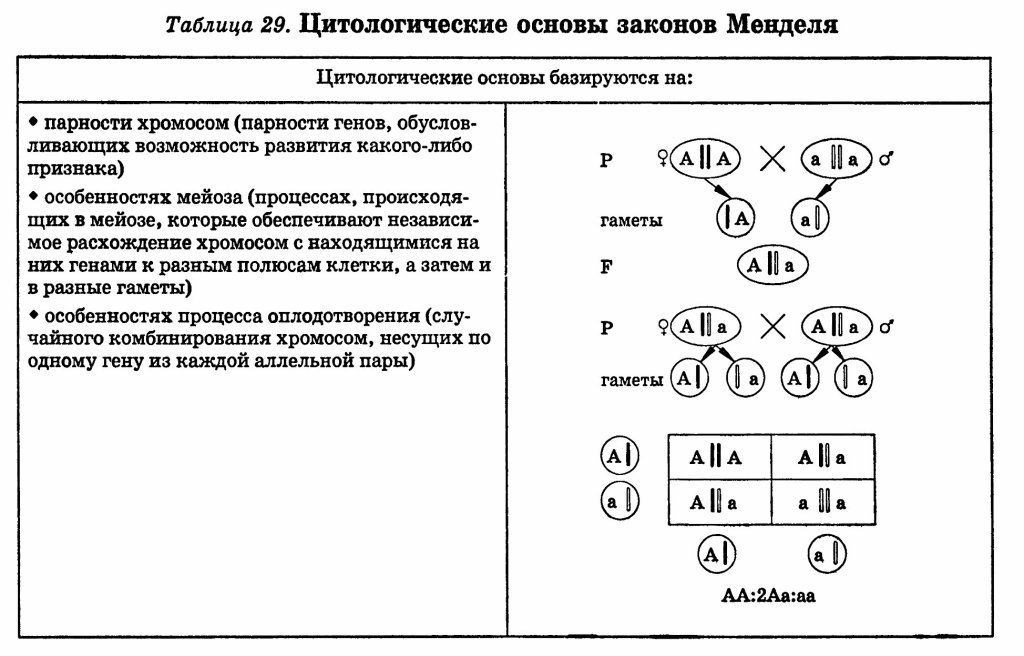
Менделирующие признаки человека. Примеры.
Менделирующими признаками называются те, наследование которых происходит по закономерностям, установленным Г. Менделем. Менделирующие признаки определяются одним геном моногенно, то есть когда проявление признака определяется взаимодействием аллельных генов, один из которых доминирует (подавляет) другой. Менделевские законы справедливы для аутосомных генов с полной пенетрантностью (от лат.penetrans-проникающий, достигающий) и постоянной экспрессивностью (степенью выраженности признака).
Если гены локализованы в половых хромосомах (за исключением гомологичного участка в Х- и У-хромосомах), или в одной хромосоме сцеплено, или в ДНК органоидов, то результаты скрещивания не будут следовать законам Менделя.
Общие законы наследственности одинаковы для всех эукариот. У человека также имеются менделирующие признаки, и для него характерны все типы их наследования: аутосомно-доминантный, аутосомно-рецессивный, сцепленный с половыми хромосомами (с гомологичным участком Х- и У-хромосом).
Примеры.
I. Аутосомно-доминантный тип наследования:
белый локон над лбом;
2) волосы жесткие, прямые (ежик);
3) шерстистые волосы - короткие, легко секущиеся, курчавые, пышные;
4) кожа толстая;
5) габсбургская губа - нижняя челюсть узкая, выступающая вперед, нижняя губа отвислая и полуоткрытый рот;
6) полидактилия (от греч.polus – многочисленный, daktylos- палец) – многопалость, когда имеется от шести и более пальцев;
7) синдактилия (от греч. syn - вместе)-сращение мягких или костных тканей фаланг двух или более пальцев;
8) брахидактилия (короткопалость) – недоразвитие дистальных фаланг пальцев;
9) арахнодактилия (от греч. агаhna – паук ) – сильно удлиненные «паучьи» пальцы
II. Аутосомно-рецессивный тип наследования.
По аутосомно-рецессивному типу наследуются следующие признаки:
1)волосы мягкие, прямые;
2)кожа тонкая;
3)группа крови Rh-;
4)фенилкетонурия – блокируется превращение фенилаланина в тирозин, который превращается в фенилпировиноградную кислоту, являющуюся нейротропным ядом
5)альбинизм.
Промежуточный тип наследования.
Промежуточный характер наследования какого-либо признака у живого организма возникает в том случае, если его фенотип (совокупность всех признаков живого организма, которая обеспечивает его индивидуальность) отличается от фенотипа обоих родителей, то есть возникает неполное доминирование. Один из родительских генов выступает как рецессивный аллель (аллель — за один признак отвечает совокупность генов), который неактивен, а доминантный аллель другого родителя не достаточно усилен для полного проявления признака в чистом виде. Выражение этого признака в фенотипе промежуточное, с уклонением в сторону доминантной аллели.
Таким образом, промежуточный характер наследования возникает при взаимодействии двух совокупностей аллелей, что является проявлением множественного действия генов.
Анализирующее скрещивание и его значение.
Анализирующее скрещивание — скрещивание гибридной особи с особью, гомозиготной по рецессивным аллелям, то есть "анализатором". Смысл анализирующего скрещивания заключается в том, что потомки от анализирующего скрещивания обязательно несут один рецессивный аллель от "анализатора", на фоне которого должны проявиться аллели, полученные от анализируемого организма. Для анализирующего скрещивания (исключая случаи взаимодействия генов) характерно совпадение расщепления по фенотипу с расщеплением по генотипу среди потомков.
Таким образом, анализирующее скрещивание позволяет определить генотип и соотношение гамет разного типа, образуемых анализируемой особью. Мендель, проводя эксперименты по анализирующему скрещиванию растений гороха с белыми цветками (аа) и пурпурных гетерозигот (Аа), получил результат 81 к 85, что почти равно соотношению 1:1.Он определил, что в результате скрещивания и образования гетерозиготы, аллели не смешиваются друг с другом и в дальнейшем проявляются в "чистом виде". В дальнейшем Бэтсон на этой основе сформулировал правило чистоты гамет.
Аллельные гены. Наследование признаков при взаимодействии аллельных генов. Примеры. Множественный аллелизм. Механизм возникновения.
Аллельные гены
У диплоидных организмов за каждый признак отвечают два гена (один от отца, другой от матери). Такие гены называются аллельными, они находятся в гомологичных хромосомах.
В половых клетках (гаплоидных) за каждый признак отвечает только один ген (на этом основано «правило чистоты гамет»).
Аллель – одна из форм одного и того же гена, определяет один из вариантов развития признака.
Наследование признаков при взаимодействии аллельных генов. Примеры.
Полное доминирование: гетерозигота имеет доминантный фенотип. Встречается, когда продукт гена требуется в небольших количествах, например, является регулятором. Например: желтый цвет гороха доминирует над зеленым, праворукость над леворукостью.
Неполное доминирование (промежуточный характер наследования): гетерозигота имеет фенотип, промежуточный между доминантным и рецессивным. Встречается, когда продукт гена является конечным продуктом и требуется в больших количествах. Например: у цветка ночной красавицы гомозиготы имеют красные и белые лепестки, гетерозигота – розовый.
Кодоминирование: оба признака проявляются одновременно, независимо. Например, при наследовании группы крови человека эритроцит IAIA будет иметь на поверхности мембраны антиген (агглютиноген) А, эритроцит IBIBбудет иметь антиген В, эритроцит IAIВ будет иметь оба антигена.
Множественный аллелизм. Механизм возникновения.
Множественный аллелизм — это существование в популяции более двух аллелей данного гена.Множественный аллелизм для генов, контролирующих системы несовместимости, выступает как фактор отбора, препятствующий образованию зигот и организмов определенных зигот. Примером множественного аллелизма являются аллели IA, Iв, I°, определяющие группы крови у человека, двенадцать различных состояний одного локуса у дрозофилы, обусловливающих разнообразие окраски глаз (w — белые, we — эозиновые, wa — абрикосовые, wch — вишневые, wm — пятнистые и т. д.); серия множественных аллелей окраски шерсти у кроликов («сплошная», гималайская, альбинос и т. д.) и т. д.
Причиной множественного аллелизма являются случайные изменения структуры гена (мутации), сохраняемые в процессе естественного отбора в генофонде популяции. Многообразие аллелей, рекомбинирующихся при половом размножении, определяет степень генотипического разнообразия среди представителей данного вида, что имеет большое эволюционное значение, повышая жизнеспособность популяций в меняющихся условиях их существования. Кроме эволюционного и экологического значения аллельное состояние генов оказывает большое влияние на функционирование генетического материала. В диплоидных соматических клетках эукариотических организмов большинство генов представлено двумя аллелями, которые совместно влияют на формирование признаков.
Неаллельные гены. Наследование признаков при взаимодействии неаллельных генов. Примеры.
Неаллельные гены — это гены, расположенные в различных участках хромосом и кодирующие неодинаковые белки. Неаллельные гены также могут взаимодействовать между собой. При этом либо один ген обусловливает развитие нескольких признаков, либо, наоборот, один признак проявляется под действием совокупности нескольких генов.
Наследование признаков при взаимодействии неаллельных генов. Примеры.
Комплементарность. Этот вид взаимодействия генов заключается в том, что при наличии двух доминантных аллелей разных генов появляется новый признак, то есть для появления нового признака у организма должен быть генотип АВ. Так, для развития окраски необходимо, чтобы в организме синтезировались определенные белки и ферменты, превращающие их в пигмент. Классическим примером является наследование окраски цветков у душистого горошка.
Эпистаз. При эпистатическом взаимодействии одна пара генов может подавлять действие другой пары генов. Например, у лошадей масть определяется двумя парами генов. В одной паре генов доминантный аллель А определяет серую окраску (раннее поседение). Этот доминантный ген подавляет действие не только аллельного ему рецессивного гена а, но и подавляет проявление другой пары генов, определяющих масть (вороную, рыжую, гнедую), вне зависимости от того, является эта пара рецессивной или доминантной гомозиготой или гетерозиготой - окраска лошади будет только серой (лошади с генотипами ААвв, Аавв, ААВВ, АаВВ или АаВв).
Полимерия. Многие признаки определяются несколькими парами генов. Это характерно, в основном, для количественных признаков, таких как яйценоскость у кур, жирность молока у коров.
Плейотропное действие гена. При плейотропном действии гена один ген определяет развитие или влияет на проявление нескольких признаков. Это свойство генов было хорошо исследовано на мышах. Из схемы, видно, что ген определяет несколько признаков и признак определяется несколькими генами, поэтому можно сделать вывод, что плейотропное действие гена неразрывно связано с полимерным взаимодействием генов.
Летальные гены. Летальность генов - одна из разновидностей плейотропного действия гена. Так один ген, определяющий какой-либо признак, влияет так же на жизнеспособность в целом.Ярким примером летальности гена служит ген платиновости у лисиц.
Генетические основы существования групп крови в системе ABO. Наследование групп крови. Наследование резус фактора. Резус-конфликт.
Генетические основы существования групп крови в системе ABO.
В основе закономерностей наследования групп крови лежат следующие понятия. В локусе гена АВО возможны три варианта (аллеля) - 0, A и B, которые экспрессируются по аутосомно-кодоминантному типу. Это означает, что у лиц, унаследовавших гены А и В, экспрессируются продукты обоих этих генов, что приводит к образованию фенотипа АВ (IV). Фенотип А (II) может быть у человека, унаследовавшего от родителей или два гена А, или гены А и 0. Соответственно фенотип В (III) - при наследовании или двух генов В, или В и 0. Фенотип 0 (I) проявляется при наследовании двух генов 0. Таким образом, если оба родителя имеют II группу крови (генотипы AА или А0), кто-то из их детей может иметь первую группу (генотип 00). Если у одного из родителей группа крови A(II) с возможным генотипом АА и А0, а у другого B(III) с возможным генотипом BB или В0 - дети могут иметь группы крови 0(I), А(II), B(III) или АВ (IV).
Наследование резус-фактора.
Наследование резус-фактора кодируется тремя парами генов и происходит независимо от наследования группы крови. Наиболее значимый ген обозначается латинской буквой D. Он может быть доминантным - D, либо рецессивным - d. Генотип резус-положительного человека может быть гомозиготным - DD, либо гетерозиготным - Dd. Генотип резус-отрицательного человека может быть - dd.
Резус конфликт.
Гемолитическая болезнь плода и новорожденного это состояние, возникающее в результате несовместимости крови матери и плода по некоторым антигенам. Наиболее часто гемолитическая болезнь новорожденного развивается вследствие резус-конфликта. При этом у беременной женщины резус-отрицательная кровь, а у плода резус-положительная. Во время беременности резус-фактор с эритроцитами резус-положительного плода попадает в кровь резус-отрицательной матери и вызывает в ее крови образование антител к резус-фактору (безвредных для нее, но вызывающих разрушение эритроцитов плода). Распад эритроцитов приводит к повреждению печени, почек, головного мозга плода, развитию гемолитической болезни плода и новорожденного. В большинстве случаев заболевание быстро развивается после рождения, чему способствует поступление большого количества антител в кровь ребенка при нарушении целостности сосудов плаценты.
Генотип как целое. Ядерная наследственность. Закономерности наследования внеядерных генов. Цитоплазматическая наследственность у про- и эукариот.
Генотип - генетическая (наследственная) конституция организма, совокупность всех его генов. В современной генетике рассматривается не как механический набор независимо функционирующих генов, а как единая система, в которой любой ген может находиться в сложном взаимодействии с остальными генами.
Большинство генов может существовать в нескольких модификациях аллелях, а поскольку число генов составляет десятки тысяч, то практически все люди различаются по генотипам. Исключение представляют однояйцевые монозиготные близнецы, имеющие совершенно одинаковые генотипы. Далеко не все гены проявляют своё действие либо находятся между собой в сложных взаимосвязях и взаимодействиях.
Патологические гены, которые обусловливают наследственные болезни и аномалии развития, также разнородны. Одни из них — доминантные — проявляют своё действие при наличии на гомологичной хромосоме нормального гена. В этих случаях болезнь передаётся из поколения в поколение и заболевают в среднем до 50% детей больного. Другие гены — рецессивные — проявляют своё действие лишь в тех случаях, когда ребёнок наследует патологический ген от каждого из клинически здоровых родителей. В таких семьях случаев аналогичного заболевания у других родственников, как правило, нет. Вероятность повторного рождения больного ребёнка в такой семье — 25%. Существуют и другие варианты действия патологических генов.
Ядерная наследственность.
Установлено, что некоторые мутации пластид вызываются ядерными генами, контролирующими отчасти и функционирование пластид. Показано также, что количество ДНК в митохондриях недостаточно для того, чтобы нести всю информацию об их функциях и строении; т. о., и структура митохондрий, по крайней мере частично, определяется геномом. Ядерные и внеядерные гены могут взаимодействовать и при реализации фенотипа.
Закономерности наследования внеядерных генов. Цитоплазматическая наследственность у про- и эукариот.
Наличие некоторого количества наследственного материала в цитоплазме в виде кольцевых молекул ДНК митохондрий и пластид, а также других внеядерных генетических элементов дает основание специально остановиться на их участии в формировании фенотипа в процессе индивидуального развития. Цитоплазматические гены не подчиняются менделевским закономерностям наследования, которые определяются поведением хромосом при митозе, мейозе и оплодотворении. В связи с тем, что организм, образуемый вследствие оплодотворения, получает цитоплазматические структуры главным образом с яйцеклеткой, цитоплазматическое наследование признаков осуществляется по материнской линии. Такой тип наследования был впервые описан в 1908 г. К. Корренсом в отношении признака пестрых листьев у некоторых растений (рис. 6.21).
Как было установлено позднее, развитие этого признака обусловлено мутацией, возникающей в ДНК хлоропластов и нарушающей синтез хлорофилла в них. Размножение в клетках нормальных (зеленых) и мутантных (бесцветных) пластид и последующее случайное распределение их между дочерними клетками приводят к появлению отдельных клеток, совершенно лишенных нормальных пластид. Потомство этих клеток образует обесцвеченные участки на листьях. Фенотип потомства, таким образом, зависит от фенотипа материнского растения. У растения с зелеными листьями потомство абсолютно нормально. У растения с бесцветными листьями потомство имеет такой же фенотип. У материнского растения с пестрыми листьями потомки могут иметь все описанные фенотипы по данному признаку. При этом внешний вид потомства не зависит от признака отцовского растения. |
|
|
 Скачать 204 Kb.
Скачать 204 Kb.