ФИЗИОЛОГИЯ МОТОРНЫХ СИСТЕМ
Виды двигательной активности.
Двигательная активность – совокупность двигательных актов.
Двигательная активность человека – одно из необходимых условий поддержания нормального функционального состояния человека, его естественная потребность. Она также может быть использована как действенное лечебное средство, как метод реабилитации больного организма, а также как метод профилактики многих заболеваний сердца, атеросклероза, гипертонической болезни, диабета, хронических обструктивных заболеваний легких, тревожных и депрессивных состояний, стрессорных реакций.
Типы двигательной активности.
1. Локомоция: активное перемещение в пространстве на расстояние большее, чем сам организм (ходьба, бег, галоп на Луне, прыжок, плавание, ныряние, полет, ползанье и т.д.). Езда на автомобиле – не локомоция!
2. Поза: активное поддержание конфигурации сегментов тела относительно друг друга и всего тела в пространстве. Лежание на диване – не поза!
3. Манипуляции: активное оперирование предметами и самой конечностью без перемещения в пространстве (вокруг себя). Письмо, клавиатура, рисование, предметная деятельность.
Роль спинного мозга в двигательной активности.
Спинной мозг занимает всего 2% (27-30 г) от веса головного мозга человека, имеет простое сегментарное строение. В спинном мозгу проходят многочисленные тракты нисходящего и восходящего направлений (белое вещество). Восходящие сенсорные тракты обеспечивают мозг, в том числе и спинной, информацией о внешней и внутренней среде, на основе которой осуществляются рефлексы и программируются сложные многосуставные движения. Нисходящие моторные тракты приносят в спинной мозг организованную команду о параметрах движения. Серое вещество спинного мозга имеет пластинчатую организацию – пластины Рекседа – содержат мотонейроны.
Пластина I (краевая зона) составлена интернейронами, которые переключают афферентную информацию о боли и температуре.
Пластина II (желатинозная субстанция) переключает самые тонкие миелинизированные и немиелинизированные афференты.
Интернейроны следующих четырех пластин (III-VI, собственное ядро) интегрируют информацию от нисходящих супраспинальных, проприоспинальных трактов и сенсорных афферентов.
Пластина VII (ядро Кларка) расположена в боковом роге и переключает информацию о движении и положении конечности к мозжечку.
В пластине VIII расположены интернейроны, управляющие импульсацией мотонейронов, например клетки Реншоу.
Мотонейроны располагаются в IX пластине, в переднем роге серого вещества.
Пластина X окружает центральный канал спинного мозга и состоит из нейроглиальных клеток.
Серое вещество содержит примерно 13 млн. нейронов, из которых 97% составляют интернейроны (вставочные), основная функция которых заключается в предварительной обработке информации от проводящих трактов. Соответственно, только 3% нейронов составляют мотонейроны.
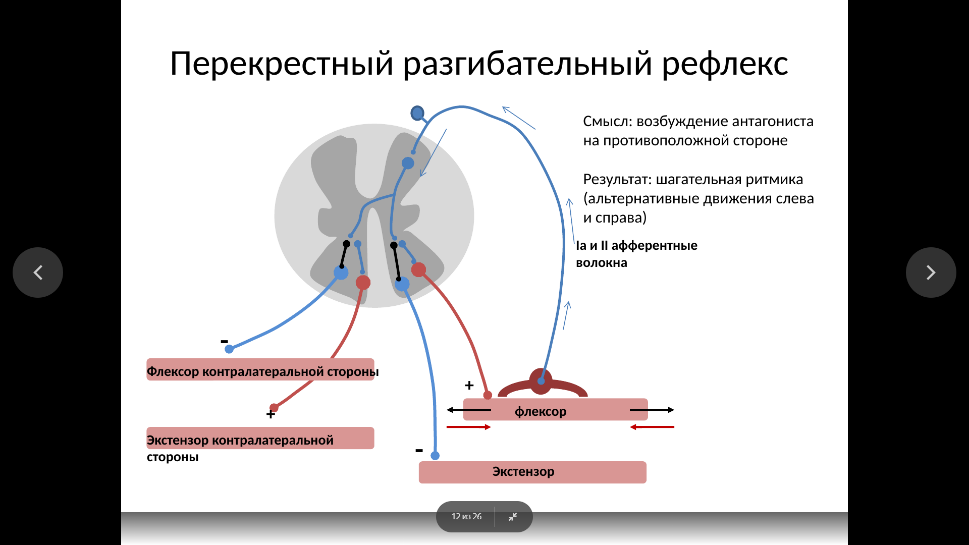
Мышечное веретено имеет длину в несколько миллиметров, одето капсулой и расположено в толще мышцы. Внутри капсулы находится пучок интрафузальных мышечных волокон. Веретена расположены параллельно внешним по отношению к капсуле экстрафузальным волокнам, поэтому при растяжении мышцы нагрузка на веретена увеличивается, а при сокращении – уменьшается. В расслабленной мышце импульсация, идущая от веретен, невелика, но они реагируют повышением частоты разрядов на удлинение мышцы. Таким образом, веретена дают мозгу информацию о длине мышцы и ее изменениях. Импульсация, идущая от веретен, в спинном мозге возбуждает мотонейроны своей мышцы и тормозит мотонейроны мышцы-антагониста, а также возбуждает мотонейроны сгибателей и тормозит мотонейроны разгибателей.
Сухожильные рецепторы Гольджи находятся в зоне соединения мышечных волокон с сухожилием и расположены последовательно по отношению к мышечным волокнам. Они слабо реагируют на растяжение мышцы, но возбуждаются при ее сокращении, причем их импульсация пропорциональна силе сокращения. Поэтому сухожильные рецепторы информируют мозг о силе, развиваемой мышцей. Идущие от этих рецепторов волокна в спинном мозге вызывают торможение мотонейронов собственной мышцы и возбуждение мотонейронов мышцы-антагониста.
Суставные рецепторы реагируют на положение сустава и на изменения суставного угла, участвуя таким образом в системе обратных связей от двигательного аппарата.
Тактильная чувствительность, ощущение, возникающее при действии на кожную поверхность различных механических раздражителей; разновидность осязания. Тактильные рецепторы расположены на поверхности кожи и некоторых слизистых оболочек (полости рта, носа). Они представлены свободными нервными окончаниями, нервными сплетениями вокруг волосяных фолликулов, а также специализированными структурами типа телец Пачини, Мейснера, дисков Меркеля и др., которые являются быстро адаптирующимися рецепторами и возбуждаются при прикосновении и вибрации. Ощущение давления возникает при возбуждении медленно адаптирующихся рецепторов.
α–Мотонейроны осуществляют передачу скелетным мышечным волокнам сигналов, выработанных в спинном мозгу. Аксоны каждого мотонейрона многократно делятся, и, таким образом, каждый из них охватывает своими терминалями до сотни мышечных волокон, образуя в совокупности с ними двигательную единицу. В свою очередь, несколько мотонейронов, иннервирующих, одну и ту же мышцу, образуют мотонейронный пул. В его состав могут входить мотонейроны нескольких соседних сегментов.
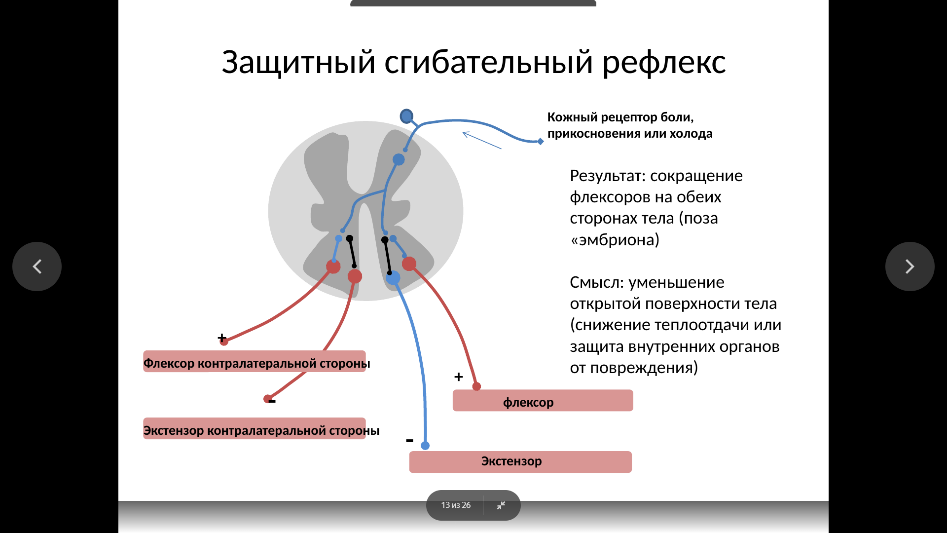
Миотатический рефлекс (рефлекс на растяжение). Это простейший, но в то же время базовый рефлекс, позволяющий автоматически регулировать длину мышцы. Хорошим примером миотатического рефлекса может служить коленный рефлекс. Рефлекс на растяжение начинается с рецепторов длины мышцы (мышечных веретен), которые растягиваются вместе с мышцей и по волокнам типа Ia и II потенциал действия входит в спинной мозг, где моносинаптически, т. е. без дополнительных переключений, образует возбуждающий синапс на α-мотонейроне собственной мышцы. В результате активируются двигательные единицы, которые вызывают сокращение мышцы. Смысл миотатического рефлекса заключается в том, что растяжение мышцы, например камбаловидной мышцы при стоянии, вызывает ее рефлекторное сокращение, что позволяет автоматически поддерживать тонус в позных мышцах и, соответственно, поддерживать позу.
Сухожильные рефлексы – это короткие, быстрые, фазные сокращения скелетных мышц. Они начинаются с рецепторов Гольджи, которые находятся в сухожилиях, связках, суставных сумках и подключены к мышцам последовательно. Для их возбуждения необходимо сильное мышечное сокращение. В данном случае рефлекторная дуга проходит по волокнам Ib, а затем контактирует с тормозным Ib-интернейроном, который тормозит активность гомонимного α-мотонейрона. В то же время Ib-интернейрон (Ib-ИН) тормозит и активность тормозящего интернейрона, управляющего α-мотонейроном мышцы-антагониста. Таким образом, при избыточном напряжении в сухожилии при сильном или быстром сокращении в мышце первичные нервные окончания с сухожильных органов Гольджи препятствуют чрезмерному сокращению и тем самым предотвращают разрыв мышцы или сухожилия. В данном случае рефлекторная дуга проходит по волокнам Ib, а затем контактирует с тормозным Ib-интернейроном, который тормозит активность гомонимного α-мотонейрона. В то же время Ib-интернейрон (Ib-ИН) тормозит и активность тормозящего интернейрона, управляющего α-мотонейроном мышцы-антагониста. Таким образом, при избыточном напряжении в сухожилии при сильном или быстром сокращении в мышце первичные нервные окончания с сухожильных органов Гольджи препятствуют чрезмерному сокращению и тем самым предотвращают разрыв мышцы или сухожилия.
Спинальный шок – это явление, наступающее при перерезке спинного мозга ниже C5 и сопровождающееся выпадением всех рефлексов ниже перерезки.
Точно указать продолжительность спинального шока после травмы невозможно, так как разные рефлексы восстанавливаются с различной скоростью.
У человека при травме шеи на уровне C4-C5 и ниже, выпадают все рефлексы: двигательные, сосудо-двигательные и рефлексы на органы малого таза – человек полностью парализован. У него низкое артериальное давление, отсутствуют дефекация и уринация. Примерно через 2-3 месяца рефлексы восстанавливаются, но они непроизвольные.
При ударе молоточком по сухожилиям наблюдаются сухожильные рефлексы, постепенно восстанавливается артериальное давление и даже становится выше нормы, восстанавливаются уринация и дефекация, но задерживать их больной не может, сфинктер прямой кишки зияет. Центральный паралич скелетных мышц сохраняется.
Механизм спинального шока заключается в отсутствии регуляторных влияний ретикулярной формации ствола мозга на нейроны спинного мозга: в начале шока отсутствуют облегчающие влияния, а затем тормозные.
Роль ствола мозга в двигательной активности.
Ствол мозга включает продолговатый мозг, варолиев мост и средний мозг.
В продолговатом мозгу имеются ядра языкоглоточного, блуждающего, добавочного и подъязычного нервов. В этой связи на уровне продолговатого мозга замыкаются некоторые жизненно важные двигательные рефлексы, осуществляемые мышцами шеи, глотки, языка, жевательными мышцами ─ жевательный рефлекс, сосательный рефлекс, чихание, кашель, глотание и рвота. В продолговатом мозгу также находится бульбарный отдел дыхательного центра, управляющий дыхательной программой.
Варолиев мост содержит ядра черепно-мозговых нервов ─ тройничного, отводящего, лицевого, преддверно-улиткового, которые участвуют в регуляции движения глаз, формировании мимики и регуляции тонуса мышц за счет вестибулоспинального тракта, а также пневмотаксический центр дыхательной программы. Через варолиев мост организуются многие входы в мозжечок.
Средний мозг представляет собой чрезвычайно полифункциональную структуру. Помимо двигательных ядер блокового и глазодвигательного нервов средний мозг содержит крупное образование ─ красное ядро, а также черную субстанцию, покрышку, крышу, серое вещество водопровода, верхние и нижние бугорки, которые в той или иной степени участвуют в регуляции позы и движений. В частности, на уровне среднего мозга замыкаются дуги выпрямительных рефлексов, сторожевых (ориентировочных) и оборонительных рефлексов. Средний мозг регулирует мышечный тонус за счет руброспинального тракта, а также участвует в более сложных двигательных функциях за счет связей черной субстанции с базальными ганглиями, регулирует содружественное движение глаз. В среднем мозгу также располагается локомоторная полоска, необходимая для осуществления шагательной программы. В среднем мозгу, как в варолиевом мосту и продолговатом мозгу, хорошо развита ретикулярная формация, которая участвует в регуляции интенсивности двигательной активности за счет активации бодрствования, в регуляции мышечного тонуса за счет ретикуло-спинальных трактов, а также дыхания.
Ретикулоспинальные тракты. Это наиболее древние двигательные тракты, которые принадлежат к вентромедиальной группе трактов и начинаются от ядер ретикулярной формации – промежуточного (ядро Кахаля), гигантоклеточного, подголубого, вентрального, орального мостового. Волокна ретикулоспинальных трактов контактируют с α- и, особенно, с γ-мотонейронами спинного мозга, которые иннервируют аксиальную мускулатуру, т. е. мускулатуру туловища и проксимальные мышцы конечностей. Волокна медиального ретикулоспинального тракта (из ядер моста) активируют преимущественно γ-мотонейроны мышц-разгибателей, а латерального тракта (из ядер продолговатого мозга) ─ α-мотонейроны мышц-сгибателей. Таким образом, проекция ретикулоспинальных трактов на γ-мотонейроны спинного мозга определяет их участие в регуляции мышечного тонуса.
Вестибулоспинальные тракты (пучок Левенталя). Латеральный вестибулоспинальный тракт начинается от латерального вестибулярного ядра (ядра Дейтерса) и проецируется на α-мотонейроны мышц-разгибателей конечностей. Медиальный вестибулоспинальный тракт проецируется на мотонейроны мышц шеи и верхней половины туловища, обеспечивая сочетанные движения головы, глаз и туловища.
Руброспинальный тракт (пучок Монакова). Филогенетически это молодой тракт, относящийся к дорсолатеральной системе моторных трактов. Руброспинальный тракт проецируется на α-мотонейроны мышц-сгибателей конечностей, преимущественно дистальных мышц. С точки зрения влияния на мышечный тонус этот тракт является антагонистом вестибулоспинального тракта. При перерезке ствола мозга между ядром Дейтерса и красным ядром наблюдается децеребрационная ригидность, характеризующаяся резким преобладанием тонуса мышцразгибателей в отсутствие влияний руброспинального тракта.
Кортикоспинальные тракты (передний и латеральный). Первые нейроны кортикоспинального тракта представлены пирамидными клетками (клетками Беца) двигательной коры мозга, которые проецируются на α-мотонейроны дистальных мышц-сгибателей. Следует отметить, что 8% проекций осуществляются моносинаптически, т. е. без вставочных нейронов, в отличие от всех других моторных трактов.

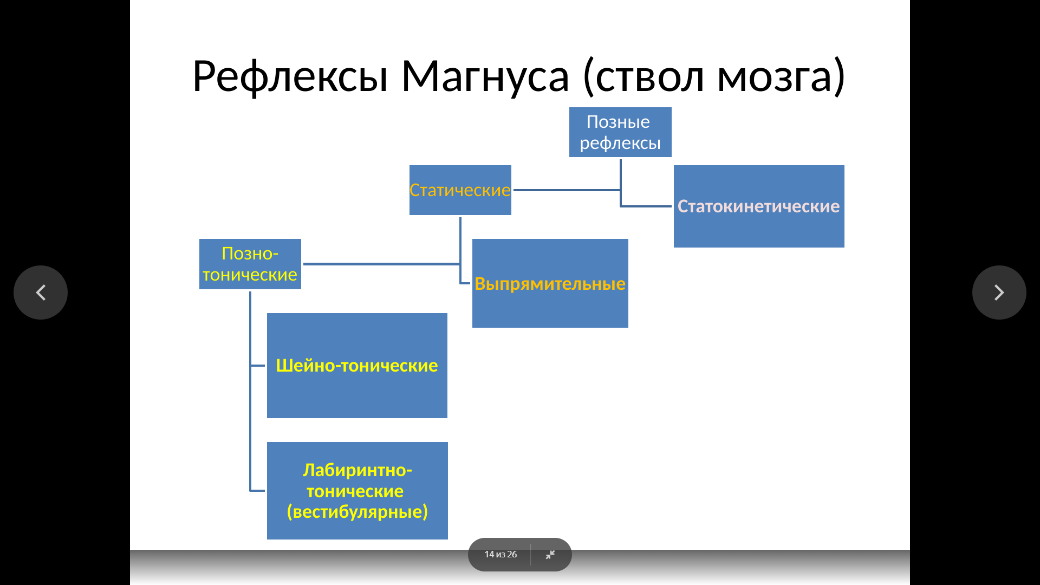
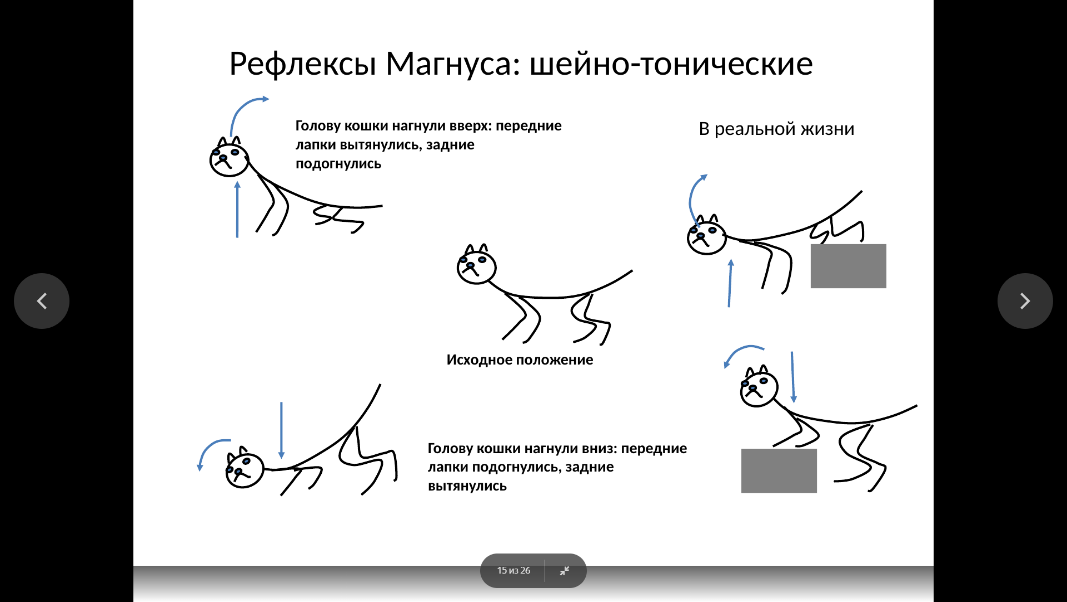
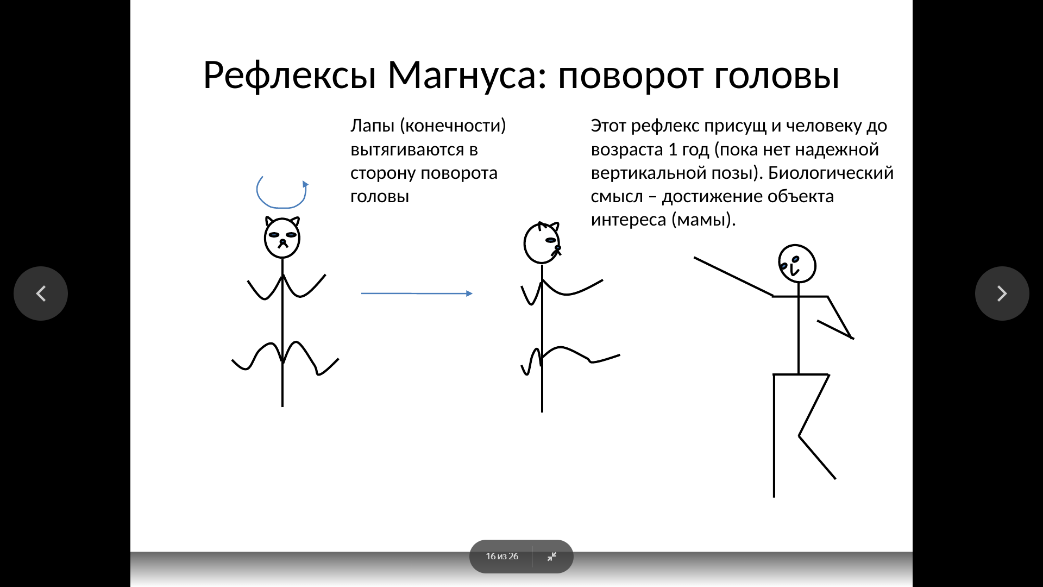
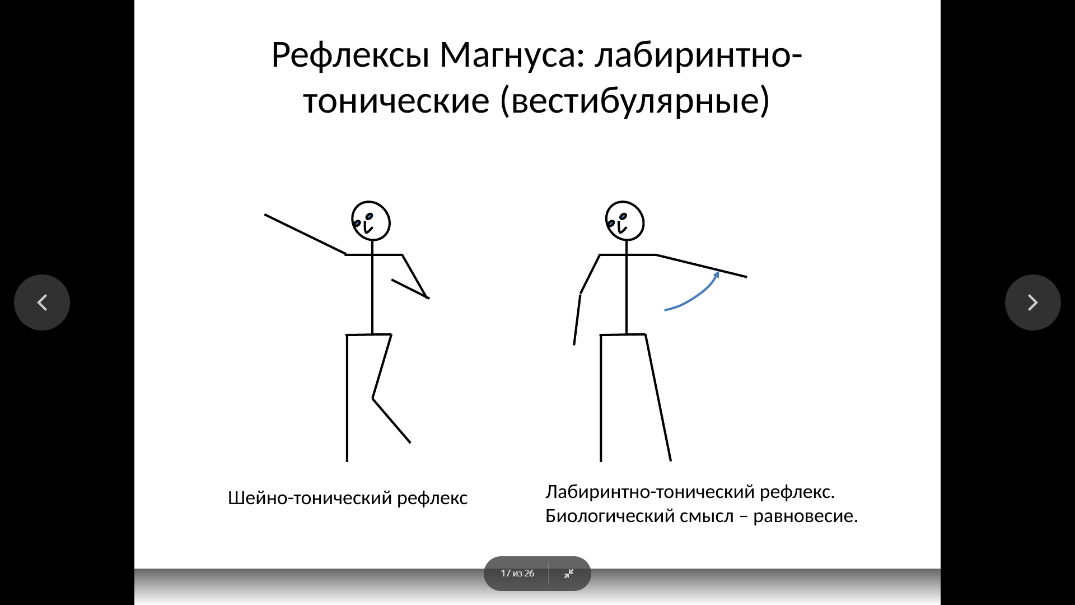
Позно-тонические рефлексы возникают при активации шейных рецепторов (шейно-тонические, или шейные рефлексы) и рецепторов лабиринтов внутреннего уха (лабиринтнотонические, или вестибулярные рефлексы).
Роль базальных ганглиев в формировании движений.
Базальные ганглии представляют собой достаточно крупное скопление серого вещества, которое находится в глубине полушарий головного мозга.
К числу базальных ганглиев относятся: полосатое тело (neostriatum), включающее хвостатое ядро (n. caudatus) и скорлупу (putamen), бледный шар (globus pallidus), состоящий из внутреннего и внешнего сегментов, субталамическое ядро (Льюисово тело), ограда (claustrum).
Ф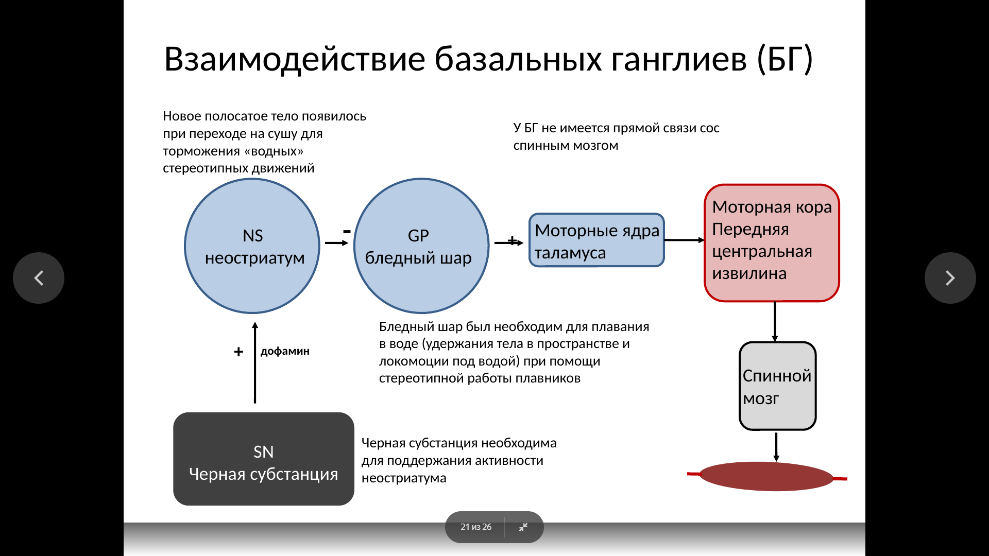

ункционально с базальными ганглиями связаны: черная субстанция (substantia nigra), вентролатеральное и вентромедиальное ядра таламуса, красное ядро (n. ruber), крыша среднего мозга.
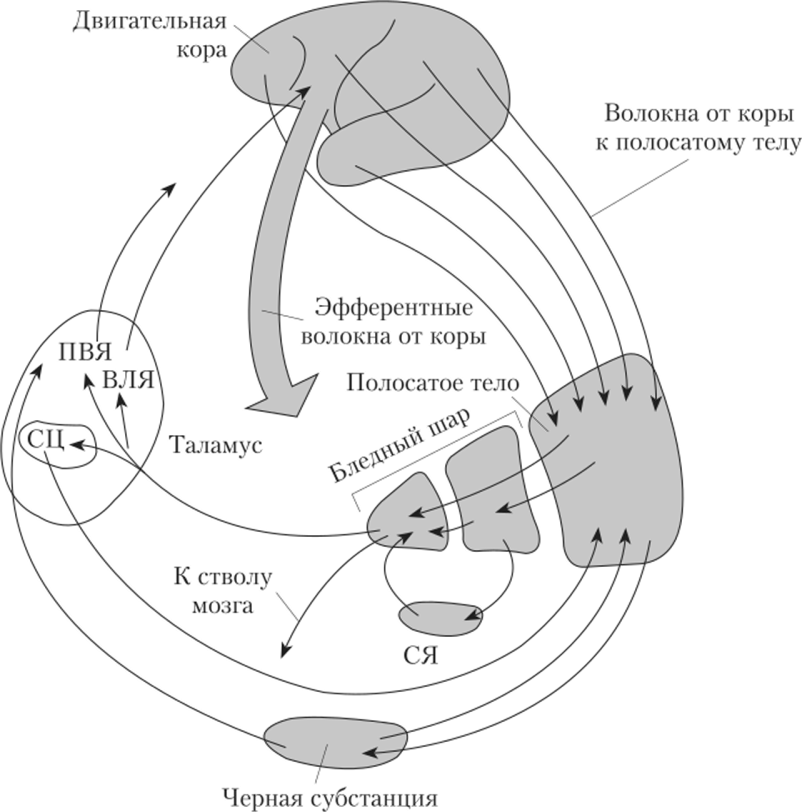
Афферентные связи (входы) базальных ганглиев. Вся информация, которая поступает в базальные ганглии, идет в первую очередь к полосатому телу. Она поступает туда из трех основных источников от всех областей коры напрямую или через таламус, из черной субстанции среднего мозга, из неспецифических ядер таламуса. Специфическим афферентным путем к стриопаллидарной системе является ретикулота-ламостриарный путь. Он обеспечивает проведение сенсорных сигналов в передний мозг. Вся кора представлена в полосатом теле, причем характерна так называемая соматотопическая организация этого представительства, т.е. каждому участку коры соответствует определенный участок в системе базальных ганглиев.
Эфферентные связи (выходы) базальных ганглиев. Информация из базальных ганглиев поступает по следующим направлениям:
• от полосатого тела к бледному шару, откуда начинается самый важный эфферентный путь базальных ганглиев в таламус (в его вентральные ядра), а затем в двигательную кору;
• из бледного шара и полосатого тела к стволу мозга (РФ, красное ядро) и далее в спинной мозг;
• от полосатого тела тормозные пути идут к черной субстанции, переключаются в ней и идут в таламус.
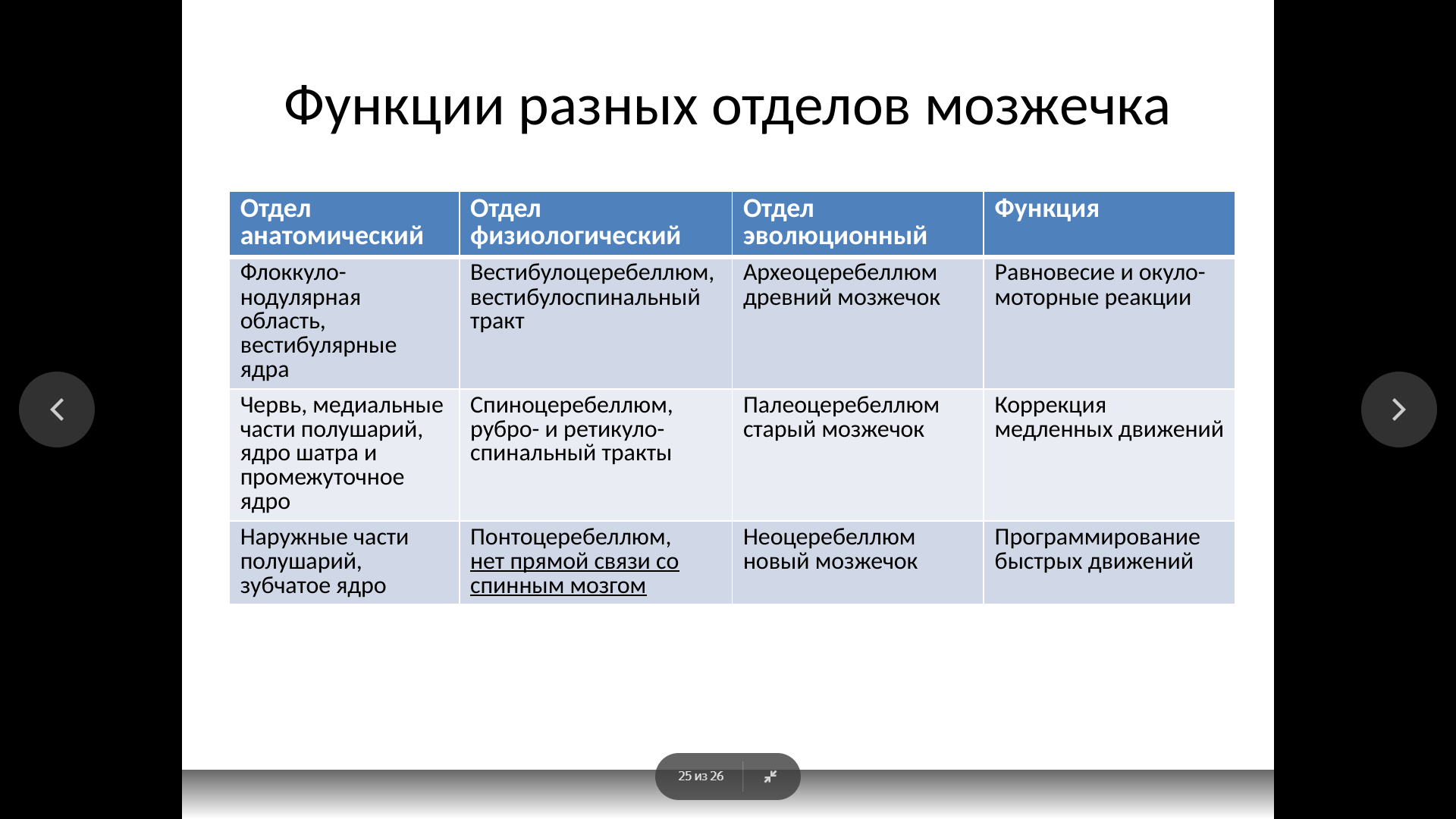
 Физиология мозжечка. Физиология мозжечка.
Мозжечок весит примерно 150 г (10% мозга) и состоит из двух полушарий, червя, флоккуло-нодулярной области, имеет ядра, сложно организованные входы и выходы, трехслойную кору, складчатую структуру коры (листки, напоминающие борозды коры больших полушарий).
В коре имеется 5 видов клеток. Главные клетки ─ клетки Пуркинье (15 млн.) ─ имеют разветвленное дендритное дерево, обращенное к поверхности коры, и упорядоченное кристаллическое расположение (рядами и колоннами). На уровне дендритов клеток Пуркинье располагаются корзинчатые и звездчатые клетки, которые, в сущности, являются интернейронами. Глубже располагаются чрезвычайно многочисленные клетки-зерна, дающие начало параллельным волокнам после Т-образного деления их аксонов, а также интернейроны в виде клеток Гольджи.
К каждой клетке Пуркинье подходит только одно лиановидное афферентное волокно, которое начинается в нижней оливе продолговатого мозга. Лиановидные волокна являются возбуждающими и образуют норадренергические синапсы на теле и дендритах клетки Пуркинье. Мшистые волокна имеют более разнообразное происхождение и активируют клетки Гольджи и клетки-зерна. Параллельные волокна от клеток-зерен образуют до 200 тыс. синапсов на каждой клетке Пуркинье. И мшистые и лазящие волокна активируют мозжечковые ядра.
Несмотря на обилие возбуждающих глутаматергических входов, большая часть внутримозжечковых синапсов ─ тормозящие ГАМКергические. Входы от голубого пятна (норадренергические) и от ядер шва (серотонинергические) также тормозящие. Поэтому уже через 1─2 синапса всякое возбуждение в мозжечке тормозится. Единственным эфферентным выходом из коры мозжечка являются аксоны клеток Пуркинье, которые контактируют с нейронами мозжечковых ядер. Клетки Пуркинье тормозящие по своему действию, поэтому моторный код коры мозжечка необычен, т. к. он тормозит активность мозжечковых ядер.
Роль коры больших полушарий в формировании движений.
На поверхности коры человека существует несколько двигательных областей, которые соответствуют полям на карте мозга (по Бродману). Наиболее известна первичная двигательная область (поле 4), которая в целом совпадает с прецентральной извилиной. В этой области в пятом слое присутствуют гигантские пирамидные клетки Беца, которые формируют пирамидный тракт. При стимуляции поля 4 очень быстро возникают двигательные реакции в соответствующих частях тела (соматотопия). Поля 5, 6 и 7 также можно назвать двигательной корой, но порог раздражения этих областей выше, чем для первичной двигательной коры. При стимуляции полей 5 и 7 (париетальная кора) обычно происходит сокращение сгибателей и разгибателей руки и ноги на своей стороне, а также противоположной ноги. Поле 6 (префронтальная кора), которое иначе называется вторичной двигательной областью, отвечает за повороты туловища, головы и глаз. Медиальную часть поля 6 называют дополнительной моторной корой, а латеральную ─ премоторной корой. При сильной стимуляции можно добиться моторных ответов также с полей 1, 2 и 3, которые относятся к первичной сенсорной коре.
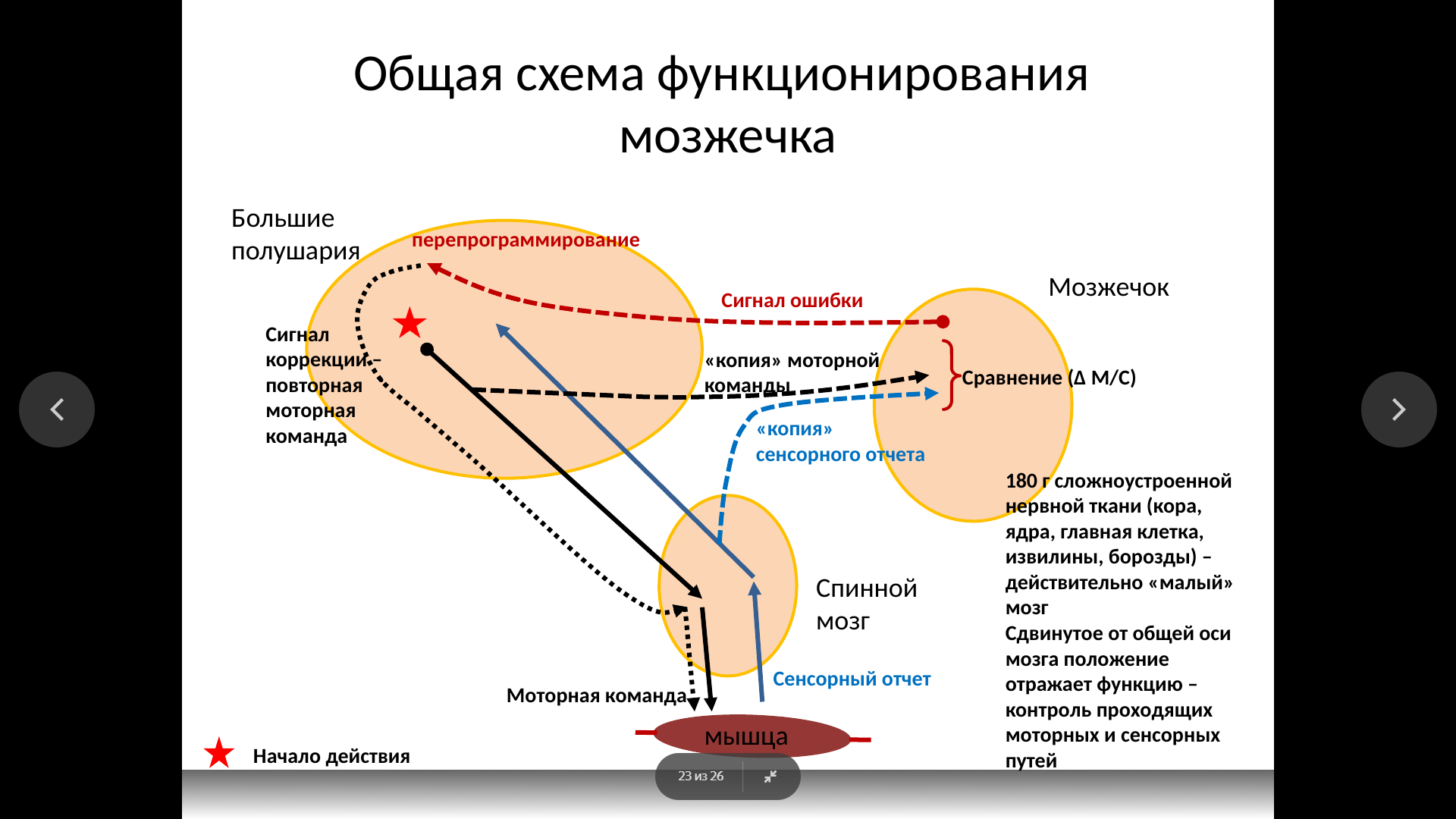
Функции первичной двигательной коры. Эфферентным выходом поля 4 является пирамидный (кортикоспинальный) тракт, формируемый гигантскими пирамидными клетками (30 000 аксонов) и малыми пирамидными клетками (500 000 аксонов). Существует устойчивое представление о том, что пирамидный тракт «отвечает» за выполнение произвольных движений. Поводом для этого служат факты паралича или пареза при инсульте, повреждающем пирамидный тракт. Вместе с тем аккуратная перерезка только самого пирамидного тракта, проведенная у приматов, показала, что устраняются не все произвольные движения, а только самые точные, требующие особенной тщательности исполнения, например, достать изюминку из углубления. Такая точность обеспечивается тем, что кортикоспинальный тракт устанавливает до 10% моносинаптических связей с мотонейронами спинного мозга. Частично (на 40%) пирамидный тракт образуется полями 5, 6 и полями сенсорной коры (1, 2 и 3), волокна из которых оканчиваются преимущественно в дорсальных рогах серого вещества спинного мозга, обеспечивая модулирующее влияние на прием соматосенсорной информации. Также при перерезке пирамидного тракта снижается мышечный тонус.
 Афферентным входом первичной моторной коры служат преимущественно волокна от мозжечка. Афферентным входом первичной моторной коры служат преимущественно волокна от мозжечка. |
 Скачать 1.38 Mb.
Скачать 1.38 Mb.