Вопросы для самоподготовки
 Скачать 0.6 Mb. Скачать 0.6 Mb.
|

 Занятие 3. Проводимость. Механизмы распространения и передачи возбуждения. Вопросы для самоподготовки
Базовая информация. Мембраны клеток возбудимых тканей обеспечивают не только возникновение, но и распространение потенциала действия, обеспечивая такое свойство возбудимых тканей как проводимость. Проводимость (conductio) - способность живой ткани проводить возбуждение, которое, возникая в рецепторе, распространяется по нервной системе в виде нервного импульса и является для организма информацией, закодированной в нейроне в виде электрических или химических сигналов. Распространение возбуждения по организму реализуется за счет двух основных механизмов: механизма распространения возбуждения по мембранам возбудимых клеток и механизма передачи возбуждения с одной возбудимой клетки на другую. Несмотря на то, что практически любая возбудимая ткань способна к проведению возбуждения, но наиболее полно эта способность выражена в нервной ткани, для которой проводимость, вместе с возбудимостью, является основной функцией. Основные нервные структуры и их роль в распространении возбуждения. Нервная ткань (textus nervosus), комплексы нервных и глиальных клеток, специфичных для животных организмов. Эволюционно появляется у кишечнополостных и достигает наиболее сложного развития в коре больших полушарий головного мозга млекопитающих. Нервная ткань - основной структурно-функциональный элемент нервной системы. Нейроглия - основной структурный элемент нервной ткани, обеспечивает существование и специфические функции нейронов, выполняет опорную, трофическую, разграничительную и защитную функции. По численности глиальных клеток в 10 раз больше, чем нейронов, и они занимают половину объема ЦНС. Основным функциональным элементом нервной ткани является нервная клетка - нейрон. Нервные клетки (нейроны) – специализированные клетки, производные эктодермы, не делятся, способны принимать, обрабатывать, кодировать, передавать и хранить информацию, организовывать реакции на раздражения, устанавливать контакты с другими нейронами и возбудимыми клетками. Именно через нейроны осуществляется передача информации от одного участка нервной системы к другому, обмен информацией между нервной системой и различными участками тела. В нейронах происходят сложнейшие процессы обработки информации. С их помощью формируются ответные реакции организма (рефлексы) на внешние и внутренние раздражения. Функциональная активность нейрона обеспечивается покрывающей его плазматической мембраной - полупроницаемой клеточной оболочкой, которая обеспечивает регуляцию концентрации ионов внутри клетки и ее обмен с окружающей средой. При возбуждении проницаемость клеточной мембраны изменяется, что играет важнейшую роль в возникновении потенциала действия и передаче нервных импульсов. Особо отметим, что сам нейрон не способен самостоятельно генерировать активность, он возбуждается нервными импульсами, поступающими с периферии от рецепторов по центростремительным нервным путям или от других нейронов. Кроме того, нервные клетки могут активироваться под влиянием гуморальных воздействий, например, клетки дыхательного центра. Нервная клетка состоит из тела, или сомы, и различных отростков. Ее форма, длина и расположение на ней отростков чрезвычайно разнообразны, и зависят от функционального назначения нейрона. Различные структурные элементы нейрона имеют разное физиологическое значение в обеспечении функций нейрона. Тела нервных клеток, суммируя приходящие к ним нервные импульсы, обеспечивают обработку поступающей информации, т.е. интегративную функцию. Результатом такой обработки является формирование на их мембране потенциала действия, основную роль в возникновении которого играет так называемый аксонный холмик, имеющий близкую к пороговой величину мембранного потенциала и потому легко возбуждающийся. Кроме того, тело нейрона выполняет трофическую функцию по отношению к отросткам, регулируя их обмен веществ. М 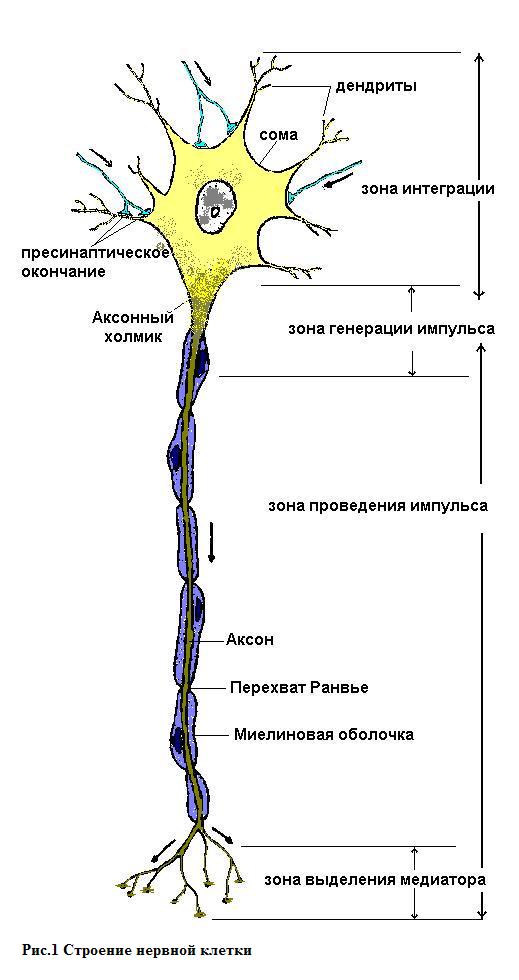 ногочисленные древовидно разветвленные отростки – дендриты – выполняют воспринимающую функцию и служат входами нейрона, по мембранам которых сигналы, то есть нервные импульсы, поступают к телу нервной клетки. Дендриты обычно образуют множество контактов с другими нервными клетками. ногочисленные древовидно разветвленные отростки – дендриты – выполняют воспринимающую функцию и служат входами нейрона, по мембранам которых сигналы, то есть нервные импульсы, поступают к телу нервной клетки. Дендриты обычно образуют множество контактов с другими нервными клетками. Аксон или нервное волокно (neurofibra) – является выходом нейрона, по которому возбуждение распространяется от тела нервной клетки дальше – к другой нервной клетке или рабочему органу (мышце, железе). Аксон всегда один, его длина по сравнению с диаметром очень велика и на периферии может достигать более метра. От сомы аксон начинается аксонным холмиком, в котором происходит формирование нервного импульса, а заканчивается синапсом – структурой, которая обеспечивает передачу возбуждения на другие возбудимые клетки. П 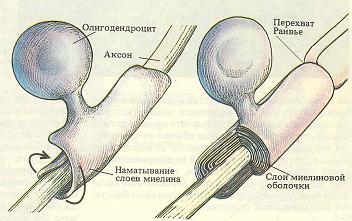 ериферические аксоны, кроме плазматической мембраны, окружены еще и оболочками, образованными различными видами глиальных клеток, эти оболочки образованы так называемыми Швановскими клетками - леммоцитами, описанными Т. Шванном в 1838 году. В зависимости от типа глиальных клеток образующих оболочки вокруг аксонов, различают безмякотные (немиелинизированные) нервные волокна в которых Швановские клетки формируют тонкую швановскую оболочку, заключающую в себе один или несколько аксонов, и мякотные (миелинизированные) нервные волокна покрытые тонкой шванновской и многослойной миелиновой оболочками. Миелиновая оболочка, состоящая из белого белково-липидного комплекса - миелина, (рис.2) образуется в результате многократного обертывания отростка Швановской клетки вокруг нервного волокна (его толщина может достигать 100 слоев) и выполняет изолирующую, опорную, барьерную, возможно трофическую и транспортную функции. ериферические аксоны, кроме плазматической мембраны, окружены еще и оболочками, образованными различными видами глиальных клеток, эти оболочки образованы так называемыми Швановскими клетками - леммоцитами, описанными Т. Шванном в 1838 году. В зависимости от типа глиальных клеток образующих оболочки вокруг аксонов, различают безмякотные (немиелинизированные) нервные волокна в которых Швановские клетки формируют тонкую швановскую оболочку, заключающую в себе один или несколько аксонов, и мякотные (миелинизированные) нервные волокна покрытые тонкой шванновской и многослойной миелиновой оболочками. Миелиновая оболочка, состоящая из белого белково-липидного комплекса - миелина, (рис.2) образуется в результате многократного обертывания отростка Швановской клетки вокруг нервного волокна (его толщина может достигать 100 слоев) и выполняет изолирующую, опорную, барьерную, возможно трофическую и транспортную функции. Процесс миелинизации является важнейшим механизмом созревания ЦНС, т.к. отсутствие миелиновой оболочки ограничивает функциональные возможности нервного волокна и делает работу ЦНС слабо координированной. Поэтому миелинизация начинается еще во внутриутробном периоде и в основном заканчивается к третьему году жизни, однако окончательно завершается только к 30 – 40 годам. Миелиновая оболочка не сплошная, по ее ходу расположены узловые перехваты Ранвье, соответствующие границам между Швановскими клетками. В местах перехвата, участок аксона не покрыт миелиновой оболочкой. Рис 2. Миелинизация нервного волокна В зависимости от скорости проведения возбуждения, длительности фаз потенциала действия и диаметра у теплокровных выделяют 3 основных группы нервных волокон, ( по Эрлангеру-Гассеру).
Нервные волокна в сумме составляют периферическую нервную систему и формируют проводящие пути в центральной нервной системе. Группы нервных волокон образуют нервы. Нервы (nervus) - тяжи нервной ткани, связывающие мозг и нервные узлы с другими тканями и органами тела. Нервы образованы пучками нервных волокон (аксонов), по аналогии это можно сравнить с многожильным телефонным кабелем, где каждый отдельный провод заизолирован и имеет определенного, точного адресата. Точность доставки сигнала имеет важнейшее значение в формировании адекватного ответа, и обеспечивается тем, что по каждому волокну нервный импульс распространяется изолированно, не переходя на другие волокна, благодаря наличию миелиновых оболочек. Кроме того, каждый нервный пучок окружен соединительнотканной оболочкой (периневрием), а весь нерв покрыт общей оболочкой (эпиневрием). Обычно нерв состоит из 103-104 волокон, у человека в зрительном нерве их даже свыше миллиона. Различают, чувствительные (афферентные, центростремительные), двигательные (эфферентные, центробежные) и смешанные нервы. У позвоночных от спинного мозга отходят спиномозговые нервы, а от головного - черепномозговые. Несколько соседних нервов могут образовывать нервные сплетения. По характеру иннервируемых органов нервы классифицируют на вегетативные и соматические, совокупность которых и образует периферическую нервную систему. Механизмы распространения возбуждения по мембранам нервных клеток Задача мембран нервного волокна состоит в распространении информации (или контролирующих сигналов), т. е. в проведении возбуждения. Распространение возбуждения по нервному волокну осуществляется по тем же законам, по которым происходит его возникновение на мембране возбудимой клетки, но в свете законов продольного распространения токов и потенциалов. Основную роль в этом процессе играют ионы Na+ и K+. Как известно, потенциалы могут быть локальными, способными распространяться с декрементом (затуханием) всего на 1 – 2 мм, и импульсными (ПД), которые распространяются без декремента по всей длине нервного или мышечного волокна: например, от мотонейронов спинного мозга до мышечных волокон конечностей с учетом и длины самих конечностей. Локальные потенциалы могут возникать в любой, клетке, способной генерировать ПД при действии подпорогового раздражения, а также в структурах нервной ткани, не имеющих, быстрых потенциалзависимых каналов (постсинаптические мембраны, мембраны сенсорных рецепторов, мембрана тела нейрона кроме аксонного холмика, мембрана дендрита, клетки нейроглии). Механизм проведения локального потенциала. Локальные потенциалы (препотенциал, рецепторный потенциал, возбуждающий постсинаптический потенциал) изменяют мембранный ПП, как правило, в сторону деполяризации в результате входа в клетку ионов Nа+. В результате этого между участком волокна, в котором возник локальный потенциал, и соседним участком мембраны формируется разность потенциалов, вызывающая передвижение ионов согласно электрическому градиенту. В частности, ионы Nа+ на наружной поверхности мембраны начинают перемещаться в сторону того участка, в котором первично возник локальный потенциал, при этом положительный потенциал наружной поверхности мембраны соседнего участка уменьшается. Аналогичные изменения наблюдаются на внутренней поверхности мембраны, где вошедшие в клетку ионы Nа+ движутся в обратном направлении, что также ведет к уменьшению отрицательного потенциала соседнего участка. В итоге поляризация соседнего участка мембраны уменьшится. Если эта частичная деполяризация соседнего участка клеточной мембраны достигнет 50% величины порогового потенциала, это вызовет открытие рядом расположенных потенциалчувствительных ионных каналов клеточной мембраны, что обеспечивает ионный ток в этом месте и, как следствие, частичную деполяризацию мембраны – локальный потенциал. Последний также действует на соседний участок мембраны и т.д., при этом главное направление движения ионов (Nа+– внутрь клетки, ионов К+– из клетки), т.е. перпендикулярно клеточной мембране. Если деполяризация соседнего участка мембраны также не достигает критического уровня, то она не обеспечивает возникновения ПД. При этом быстро развивается инактивация Nа-каналов, поэтому деполяризация быстро сменяется реполяризацией вследствие выхода ионов К+ из клетки по неуправляемым каналам (каналы утечки ионов), в результате чего локальный потенциал затухает. Подобный механизм распространения локального потенциала наблюдается только в тех мембранах, торые содержат потенциалчувствительные ионные каналы. Если возникшая деполяризация мембраны не сопровождается изменением проницаемости потенциалзависимых натриевых, кальциевых и калиевых каналов, такую деполяризацию называют электротонической (физической). Электротоническое распространение возбуждения характерно для фрагментов мембран возбудимых клеток, где нет потенциалзависимых ионных каналов, и ионы движутся только вдоль мембраны волокна. Такими участками являются, например, мембраны дендритов большинства нервных клеток, межперехватные промежутки в миелиновых нервных волокнах. Если распространяющееся локальное возбуждение достигает участков мембраны, способных генерировать ПД (перехваты Ранвье, аксонный холмик нейрона), и их амплитуда выходит на критический уровень деполяризации, формируется ПД, распространяющийся по всей длине волокна. Эффективность электротонического распространения биопотенциалов зависит от физических свойств нервного волокна: сопротивления, емкости мембраны, сопротивления цитоплазмы. Электротоническое проведение в нервном волокне улучшается при увеличении его диаметра, что связано с уменьшением сопротивления цитоплазмы, а также при миелинизации волокна, увеличивающей сопротивление мембраны. Проводимость нервного волокна для локального потенциала характеризует постоянная длины мембраны λm – расстояние, на которое может электротонически распространиться биопотенциал, пока его амплитуда не уменьшится до 37% от исходной величины. Постоянная длины мембраны тонких безмиелиновых волокон не превышает 1 мм, толстых миелиновых волокон достигает 5 мм. Передача информации на большие расстояния в пределах нервной системы осуществляется с помощью нервных импульсов (ПД) по аксонам нейронов. Механизм проведения потенциала действия. Обязательным условием проведения нервного импульса является наличие на всем протяжении или в ограниченных, но повторяющихся участках волокна быстрых потенциалзависимых (потенциалчувствительных) каналов, ответственных за формирование ПД. В распространении ПД можно выделить два этапа: этап электротонического проведения, обусловленного физическими свойствами нервного волокна, и этап генерации ПД в новом участке на пути его движения. В зависимости от расположения и концентрации ионных каналов в мембране волокна возможно два типа проведения ПД: непрерывный и сальтаторный (скачкообразный). Н 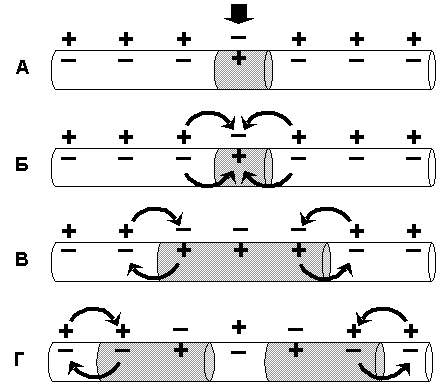 епрерывное проведение нервного импульса (ПД) осуществляется в безмиелиновых волокнах типа С, что объясняется равномерным распределением в них потенциалчувствительных ионных каналов, участвующих в генерации ПД. (рис. 3) епрерывное проведение нервного импульса (ПД) осуществляется в безмиелиновых волокнах типа С, что объясняется равномерным распределением в них потенциалчувствительных ионных каналов, участвующих в генерации ПД. (рис. 3)Во время развития потенциала действия в возбужденном участке мембраны происходит реверсия заряда (рис. 3А). Возникающий потенциал действия, обеспечивает открытие потенциалзависимых Nа-каналов на соседнем участке мембраны нервного волокна и электротоническое движение вдоль волокна ионов Nа+ снаружи, а анионов внутри волокна, (рис 3 Б), что обеспечивает развитие критического уровня деполяризации на соседнем участке нервного волокна и возникновение нового ПД. (рис. 3 В). В то же время ранее возбужденные участки возвращаются в состояние покоя (рис.3 Г). Таким образом, волна возбуждение охватывает все новые участки мембраны нервного волокна. Рис.3. Механизм непрерывного проведения нервного импульса. Постоянная длины мембраны (λm) безмиелиновых волокон составляет примерно 0,1 – 1,0 мм, т.е. эта величина во много раз больше расстояния между отдельными каналами, что обеспечивает высокую надежность проведения ПД, способного деполяризовать мембрану до критического уровня и обеспечить генерацию нового ПД. Следует также отметить, что чисто электротонический этап распространения ПД (вдоль мембраны) в безмиелиновом волокне предельно мал и наблюдается только до достижения деполяризации мембраны 50% величины порогового потенциала. Далее включается перпендикулярное перемещение ионов за счет активации ионных каналов, при этом в каждом участке мембраны ПД возникает заново, поэтому он проводится без снижения амплитуды – без декремента. Непрерывное распространение нервного импульса идет через генерацию новых ПД по эстафете, когда каждый возникший импульс является раздражителем для соседнего участка нервного волокна и обеспечивает возникновение нового ПД. С 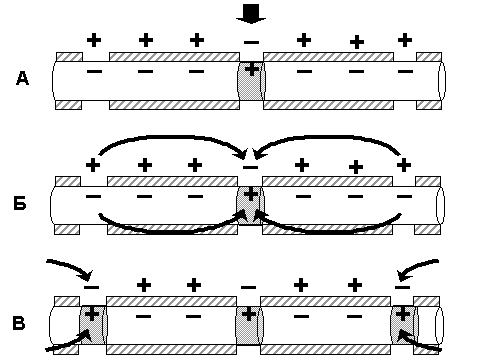 альтаторное проведение нервного импульса (ПД) осуществляется в миелиновых волокнах. В этих волокнах, в области миелиновых муфт (межузловые сегменты), обладающих высокими изолирующими свойствами, потенциалчувствительных каналов почти нет, вследствие чего мембрана там практически невозбудима. Потенциалчувствительные ионные каналы локализованы только в участках мембраны перехватов Ранвье, где при развитии ПД и происходит реверсия заряда мембраны (рис. 4 А). ПД, возникший в одном перехвате Ранвье, электротонически (как и в безмиелиновых волокнах) распространяется между электроотрицательными и электроположительными участками мембраны (рис.4 Б). Однако, в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье (рис. 3 В), т.е. возбуждение проводится скачкообразно (сальтаторно). альтаторное проведение нервного импульса (ПД) осуществляется в миелиновых волокнах. В этих волокнах, в области миелиновых муфт (межузловые сегменты), обладающих высокими изолирующими свойствами, потенциалчувствительных каналов почти нет, вследствие чего мембрана там практически невозбудима. Потенциалчувствительные ионные каналы локализованы только в участках мембраны перехватов Ранвье, где при развитии ПД и происходит реверсия заряда мембраны (рис. 4 А). ПД, возникший в одном перехвате Ранвье, электротонически (как и в безмиелиновых волокнах) распространяется между электроотрицательными и электроположительными участками мембраны (рис.4 Б). Однако, в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье (рис. 3 В), т.е. возбуждение проводится скачкообразно (сальтаторно). Движение ионов Nа+ внутрь миелинового волокна (перпендикулярно) имеется только в области перехвата Ранвье. Как отмечалось, постоянная длины мембраны (λm) миелинового волокна достигает 5 мм. Это обеспечивает высокую надежность проведения ПД, так как он, распространяясь электротонически на это расстояние, сохраняет 37% от своей амплитуды (около 30 мВ) и может деполяризовать мембрану до критического уровня (пороговый потенциал в перехватах Ранвье равен около 15 мВ). Даже в случае повреждения ближайших перехватов Ранвье на пути следования ПД он может электротонически возбудить 2 – 4-й и даже 5-й перехваты. Рис.4. Механизм сальтаторного проведения нервного импульса Сальтаторное проведение нервных импульсов является эволюционно более поздним механизмом, возникшим впервые у позвоночных в связи с миелинизацией нервных волокон. Оно имеет два Важных преимущества по сравнению с непрерывным механизмом проведения возбуждения:
|