Вопросы для самоподготовки
 Скачать 0.6 Mb. Скачать 0.6 Mb.
|

|
Закономерности проведения возбуждения по нервным волокнам. 1. Двустороннее проведение возбуждения, т.е. способность возбуждения распространяться по нервному волокну в обе стороны от места возникновения. В условиях целого организма, в норме, возбуждение всегда распространяется по аксону от тела клетки (ортодромно). Двустороннее проведение наблюдается в аксонном холмике нейрона, возникающий в этом месте потенциал действия, переходит не только на аксон, но и на тело нейрона, но из-за отсутствия потециалчувствительных каналов, распространяться по телу клетки не может. 2. Изолированное проведение возбуждения в отдельных нервных волокнах. Обычно оно не передается с одного нервного волокна на другое. Это обусловлено тем, что петли тока в межклеточной жидкости ствола, имеющей низкое сопротивление, почти не проникают в невозбужденные волокна нерва вследствие высокого сопротивления их оболочек. Изолированное проведение импульсов по нервным волокнам обеспечивает высокую точность регуляторной деятельности ЦНС на другие нервные клетки и клетки эффекторы рабочего органа. Полная изолированность достигается за счет миелинизации нервных волокон, которая завершатся в основном к 3 году жизни, но окончательно только к 30 - 40. 3. Неутомляемость нервного волокна. Н.Е. Введенский (1883) обнаружил, что нерв сохраняет способность к проведению возбуждения в течение 6 – 8 ч непрерывного раздражения. Это обусловлено тем, что при проведении ПД по нервным волокнам используется 1/1 000 000 часть запасов трансмембранных ионных градиентов и, следовательно, нужны небольшие количества АТФ для восстановления ионных градиентов. Расход энергии в нерве примерно в 16 раз меньше, чем на соответствующую единицу массы в целом организме в условиях покоя. 4. Большая скорость проведения возбуждения, достигающая 120м/с. Скорость проведения возбуждения по нервному волокну, является основной функциональной характеристикой его работы. У разных волокон, эта скорость различна, она прямо пропорциональна диаметру волокна: с утолщением аксонов она увеличивается и всегда выше в миелинизированных нервных волокнах. Скорость проведения по нервному волокну можно определить путем сложного расчета, зная зависимость ионных токов от потенциала и времени, а также условия, определяющие электротоническое распространение - диаметр волокна, сопротивление мембраны и емкость мембраны. Результаты такого расчета близки к экспериментальным данным, что подтверждает справедливость ионной теории возбуждения и электротона. Здесь мы обсудим только качественные факторы, влияющие на скорость проведения. Одним из таких факторов является амплитуда входящего Nа+ -тока, поскольку, чем больше ток после возбуждающего разряда мембраны, тем больше ток, который потечет через соседние, еще не возбужденные участки, и деполяризация этих участков произойдет быстрее. Электротоническое распространение мембранных токов также является очень важным для скорости проведения. Поскольку сопротивление и емкость элементарного участка мембраны практически одинакова во всех возбудимых клетках, электротоническое распространение определяется главным образом диаметром волокна. Поверхность мембраны нервного волокна пропорциональна его диаметру, а поперечное сечение волокна возрастает пропорционально квадрату диаметра. Поэтому при увеличении диаметра волокна продольное сопротивление его внутренней среды, определяемое площадью поперечного сечения, снижается относительно сопротивления мембраны. В результате электротонические токи распространяются более широко (увеличивается постоянная длины) и возрастает скорость проведения. Хотя с увеличением диаметра волокна емкость мембраны тоже возрастает пропорционально площади мембраны (что ведет к уменьшению скорости проведения), преобладает эффект снижения продольного сопротивления. В конечном результате скорость проведения возрастает пропорционально корню квадратному от диаметра волокна. 5. Необходимость анатомической и физиологической целостности нервного волокна. Проведение возбуждения по нервному волокну возможно лишь в том случае, если сохранена его анатомическая и физиологическая целостность. Различные факторы, воздействующие на нервное волокно (наркотические вещества, охлаждение, перевязка и т. д.) приводят к нарушению физиологической целостности, т. е. к нарушению механизмов передачи возбуждения. Несмотря на сохранение его анатомической целостности проведение возбуждения в таких условиях нарушается. Такое нарушение известно как парабиоз. 6. Возможность функционального блока проведения возбуждения при морфологической целостности нервных волокон (парабиоз). Понятие о парабиозе (para - около, bios- жизнь) в физиологию нервной системы введено Н. Е. Введенским. В 1901 г. вышла в свет монография Н. Е. Введенского «Возбуждение, торможение и наркоз», в которой он на основании своих исследований высказал предположение о единстве процессов возбуждения и торможения. Н. Е. Введенский обнаружил, что в ответ на действие различных повреждающих агентов приводящих к нарушению физиологической непрерывности нервных волокон, которые можно наблюдать при действии на нерв анестетиков, различных ядов, растворов солей, новокаина, при гипоксии или охлаждении возбудимые ткани отвечают своеобразной фазной, одинаковой во всех случаях реакцией, которую он назвал парабиозом. При этом на раздражаемом участке формируется функциональный блок, через который не проходит нервный импульс. Причиной блока являются специфические расстройства работы ионных каналов мембраны под влиянием альтерирующих агентов, в результате чего изменяются физиологические свойства ткани, в первую очередь резко снижается ее лабильность. Для создания блока протяженность парабиотического участка должна превысить постоянную длины мембраны, иначе ПД может распространиться через этот участок электротонически. Классические опыты Н. Е. Введенского по изучению парабиоза были выполнены на нервно-мышечном препарате лягушки. Нерв на небольшом участке подвергали повреждению (альтерация) химическими веществами (кокаин, хлороформ, фенол, хлорид калия), сильным фарадическим током, механическим фактором, Затем наносили раздражение электрическим током на альтерированный участок нерва или же выше его. Таким образом, импульсы должны были или возникать в альтерированном отрезке нерва, или проходить через него на своем пути к мышце. Сокращение мышцы свидетельствовало о проведении возбуждения по нерву. Развитие парабиоза протекает в три стадии: провизорную, парадоксальную и тормозную. Первая стадия парабиоза провизорная, уравнительная, или стадия трансформации. Эта стадия предшествует остальным, отсюда ее название провизорная. Уравнительной ее называют потому, что в этот период развития парабиотического состояния мышца отвечает одинаковыми по амплитуде сокращениями на сильные и слабые раздражения, наносимые на участок нерва, расположенный выше альтерировянного. В первую же стадию парабиоза наблюдается трансформация (переделка, перевод) частых ритмов возбуждеиия в более редкие. Все описанные изменения ответной реакции мышцы и характера возникновения волн возбуждения в нерве под влиянием раздражения являются результатом ослабления функциональных свойств, особенно лабильности, в альтерированном участке нерва. Вторая стадия парабиоза парадоксальная. Эта стадия возникает в результате продолжающихся и углубляющихся изменений функциональных свойств парабиотического отрезка нерва. Особенностью этой стадии является парадоксальное отношение альтерированного участка нерва к слабым (редким) или сильным (частым) волнам возбуждения, приходящим сюда с нормальных участков нерва. Редкие волны возбуждения проходят через парабиотический отрезок нерва и обусловливают сокращение мышцы. Частые же волны возбуждения либо совсем не проводятся, как бы затухают здесь, что наблюдается при полном развитии этой стадии, либо вызывают такой же сократительный эффект мышцы, как и редкие волны возбуждения, или менее выраженный. Третья стадия парабиоза тормозная. Характерной особенностью этой стадии является то, что в парабиотическом участке нерва не только резко снижены возбудимость и лабильность, но он также теряет способность проводить к мышце и слабые (редкие) волны возбуждения. Парабиоз явление обратимое. При устранении причины, вызвавшей парабиоз, физиологические свойства нервного волокна восстанавливаются. При этом наблюдается обратное развитие фаз парабиоза тормозная, парадоксальная, уравнительная. Наличие электроотрицательности в альтерированном участке нерва позволило Н. Е. Введенскому рассматривать парабиоз как особый вид возбуждения, локализованный в месте его возникновения и не способный распространяться. Синапсы в центральной нервной системе. Распространение возбуждения обеспечивается е только за счет его распространения по мембранам нервных клеток, но и за счет передачи его с одной возбудимой клетки на другую. Эту функцию выполняют синапсы. Синапс(-ы) (греч. sinapsis соединение, связь) - специализированная зона контакта образуемая между нервным окончанием (аксоном) и другими возбудимыми структурами, обеспечивающая передачу возбуждения с сохранением, изменением или исчезновением ее информационного значения. Название синапс, было дано Шеррингтоном в конце позапрошлого – начале - прошлого века. Синапсы играют решающую роль в функции мозга по следующим причинам. Во-первых, они работают по принципу клапана, проводя возбуждение только в одну сторону и обеспечивая, таким образом, упорядоченность в деятельности центральной нервной системы. Во-вторых, эффективность работы синапсов непостоянна, передача сигнала происходит тем лучше, чем чаще он используется в работе. При отсутствии активации синапса даже в течение нескольких дней уже происходит снижение (гипосинапсия), а при более длительном бездействии и полное угнетение (асинапсия) их функциональной активности. Обладая, таким образом, определенной степенью пластичности синапсы играют важнейшую роль в таких функциях, как научение и память. В-третьих, именно синапсы являются точкой приложения многих фармакологических веществ, начиная от блокаторов нервно-мышечной передачи и заканчивая психомиметическими средствами. По способу передачи сигнала, различают химические и электрические синапсы. Химический синапс – тот, в котором возбуждение от мембраны нервного окончания (пресинаптической мембраны) к мембране другой клетки (постсинаптической мембране) передается с помощью химического вещества - медиатора, содержащегося в окончании аксона - синаптическом окончании. Передача возбуждения через химический синапс отличается большой специализированностью. К химическим, относятся абсолютное большинство синапсов и изучены они наиболее полно. Электрический - синапс, в котором возбуждение передается электрическим путем за счет местных токов и низкого сопротивления мембраны. Медиатор в этих синапсах не вырабатывается. Электрические синапсы встречаются значительно реже, чем химические, и отличаются от них большей скоростью передачи возбуждения, высокой надежностью передачи, возможностью двустороннего проведения возбуждения. Химические синапсы классифицируются по анатомическому, нейрохимическому и функциональному принципам. По анатомическому принципу, т.е. по месту расположения, синапсы делятся на нейросекреторные, нервно-мышечные и межнейронные. Нейросекреторный - синапс между нервом и экзокринной или эндокринной железой. Нервно-мышечный - между аксоном двигательного нейрона и скелетным мышечным волокном. Межнейронный - между двумя нейронами. Межнейронные синапсы, в зависимости от места их расположения, бывают аксо-аксональные, аксо-соматические, аксо-дендритические и дендро-дендритические. Отметим, что дендро-дендритические синапсы выделены только гистологически и функциональное значение их неясно. По нейрохимической классификации синапсы различаются по природе медиатора с помощью которого реализуется их эффект. Несмотря на то, что один нейрон, за счет ветвления аксона на его конце, может иметь несколько синапсов, во всех синапсах одной нервной клетки производится один и тот же медиатор (принцип Дейла), поэтому и возможна классификация синапсов по этому признаку. Различают адренергические синапсы – медиатор адреналин, холинергические синапсы – медиатор ацетилхолин, дофаминергические синапсы – медиатордофамин и т.д. В синапсах мозга роль медиаторов могут выполнять около 30 биологически активных веществ, которые помимо проводников возбуждения выполняют и нейросекреторную роль. Нервно-мышечные синапсы скелетных мышц, наоборот, все имеют один и тот же медиатор - ацетилхолин В функциональном плане синапсы делятся на возбуждающие и тормозные. Возбуждающий – тот, в котором под действием медиатора происходит деполяризация постсинаптической мембраны, и на ней возникает возбуждающий постсинаптический потенциал(ВПСП). При этом пришедшее к синапсу возбуждение распространяется дальше. Тормозной - тот, котором под действием медиатора происходит гиперполяризация постсинаптической мембраны, на ней возникает тормозной постсинаптический потенциал (ТПСП), затрудняющий распространение возбуждения. Кроме того, тормозной эффект может вызвать возбуждающий аксо-аксональный синапс, который при высокой частоте импульсации приводит к возникновению устойчивой деполяризации иннервируемой мембраны и возникновению на ней состояния рефрактерности, что делает невозможным проведение возбуждения через этот участок аксона. Ультраструктура химического синапса. Основными и наиболее изученными синаптическими образованиями играющими роль в деятельности нервной системы являются химические синапсы (Рис. 5). 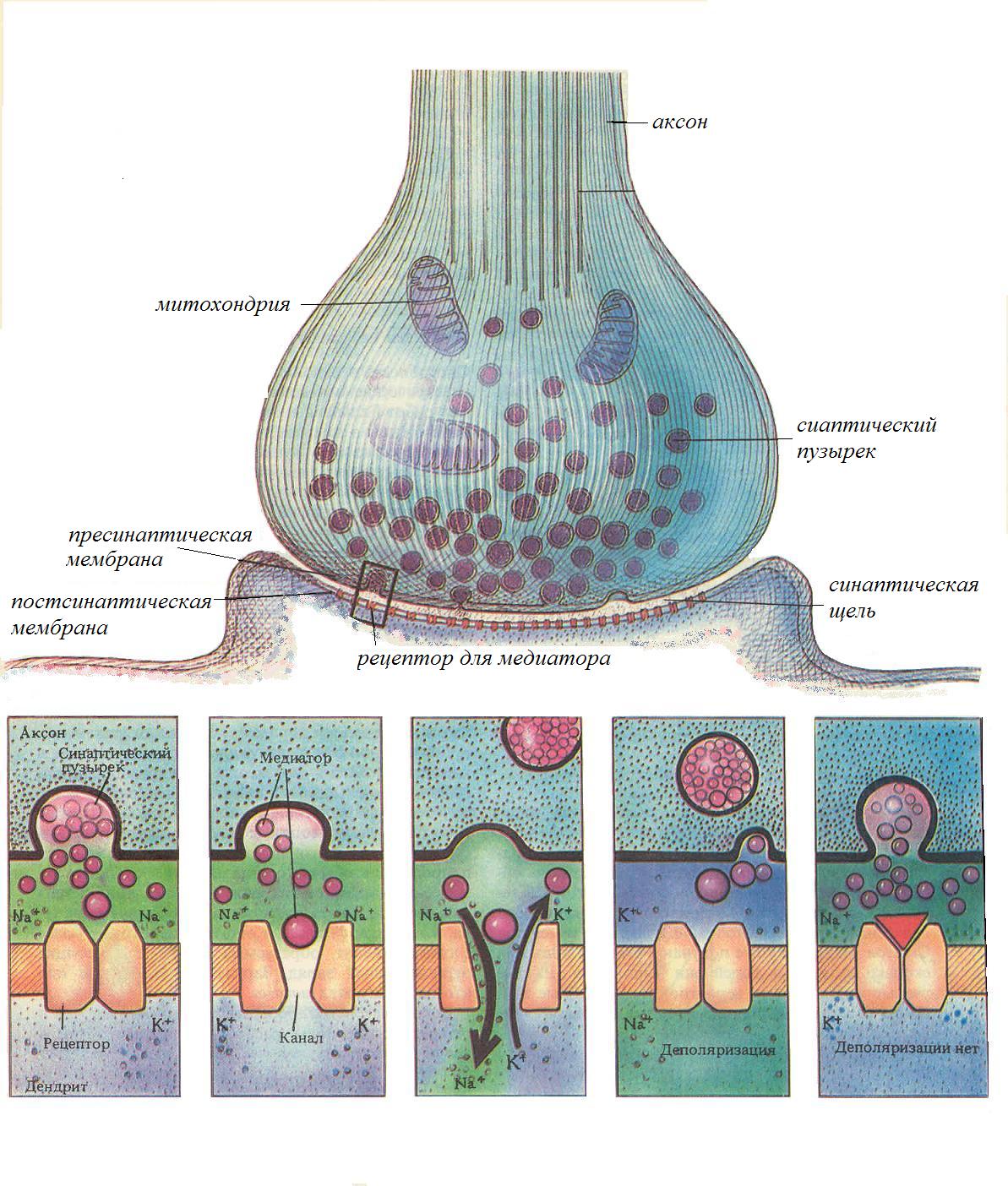 Рис. 5. Структура химического синапса. Их пресинаптическое окончание образует характерное утолщение, покрытое пресинаптической мембраной. Между нервным окончанием и эффекторной клеткой имеется пространство называемое синаптической щелью, которая отделяет нервное окончание от мембраны эффекторной клетки. Часть мембраны эффекторной клетки лежащая за синаптической щелью называется постсинаптической мембраной. Эта мембрана имеет белковые хеморецепторы, которые связаны с хемо-управляемыми ионными каналами. Связывание рецепторов с биологически активными веществами (медиаторами, гормонами, лекарственными препаратами), приводит к открытию канала, движению ионов через него и изменению заряда мембраны. Повышенная химическая чувствительность постсинаптической мембраны является важным свойством синапса, регулирующим его активность. Активность синапса заключается в синаптической передаче - распространении возбуждения через синапс. Этот процесс является достаточно сложным и протекает в несколько стадий. 1. Синтез медиатора. Химические медиаторы, это вещества различной химической природы, среди которых различают моноамины (ацетилхолин, дофамин, норадреналин, серотонин и др.); аминокислоты (гамма-аминомасляная кислота - ГАМК, глутаминовая кислота, глицин и др.); нейропептиды (вещество Р, эндорфины, нейротензин, АКТГ, ангиотензин, вазопрессин, соматостатин и др.), поэтому их биосинтез разнообразен. Ацетилхолин, например, синтезируется из холина (продукт секретируемый печенью) и уксусной кислоты при участии ферментов нервной клетки; норадреналин синтезируется из аминокислот тирозина и фенилаланина. Медиаторы непрерывно синтезируются в нервной клетке и депонируются (складируются) в синаптических пузырьках нервных окончаний куда они поступают: во-первых из околоядерной области тела нейрона с помощью быстрого аксонального транспорта, во-вторых за счет синтеза медиатора, протекающего в самих синаптических окончаниях из продуктов его расщепления. Кроме того, медиатор может поступать в синаптическое окончание за счет обратного захвата его из синаптической щели в неизменном виде. Следует отметить, что скорость биосинтеза медиатора невелика, и при больших нагрузках на синапс, может значительно отставать от скорости его разрушения. Работоспособность синапса при этом снижается. Именно эта причина является основной, при развитии всех видов утомления и время требуемое на отдых, это время требуемое на восстановление количества медиатора необходимого для полноценной работы синапса. 2. Секреция медиаторов. В 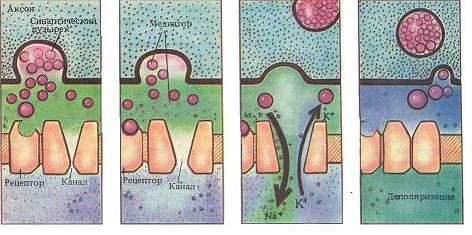 ысвобождение медиатора из синаптических пузырьков осуществляется дискретно, квантами (порциями). В состоянии покоя оно незначительно, и не вызывает никакого эффекта, но под влиянием нервного импульса, приходящего по аксону, резко усиливается. Возбуждение, деполяризует мембрану синаптического окончания, что приводит к активации имеющихся на ней Са2+-каналов и входу ионов кальция в синаптическое окончание. Пузырьки находящиеся у пресинаптической мембраны, под действием Са2+ разрушаются (по механизму экзоцитоза) и медиатор попадает в синаптическую щель. Медиатор выделившийся из синаптических пузырьков пресинаптического окончания диффундирует в синаптической щели и достигает постсинаптической мембраны. ысвобождение медиатора из синаптических пузырьков осуществляется дискретно, квантами (порциями). В состоянии покоя оно незначительно, и не вызывает никакого эффекта, но под влиянием нервного импульса, приходящего по аксону, резко усиливается. Возбуждение, деполяризует мембрану синаптического окончания, что приводит к активации имеющихся на ней Са2+-каналов и входу ионов кальция в синаптическое окончание. Пузырьки находящиеся у пресинаптической мембраны, под действием Са2+ разрушаются (по механизму экзоцитоза) и медиатор попадает в синаптическую щель. Медиатор выделившийся из синаптических пузырьков пресинаптического окончания диффундирует в синаптической щели и достигает постсинаптической мембраны.Скорость экзоцитоза зависит от количества входящего кальция и активности метаболических процессов. Она увеличивается при введении лекарств содержащих кальций, АТФ, биогенных стимуляторов (алоэ, женьшень, боярышник), гормонов. 3. Взаимодействие медиатора с рецепторами постсинаптической мембраны. М едиатор достигший постсинаптической мембраны действует на ее хеморецепторы, связанные с различными хемо-управляемыми ионными каналами, вследствие чего изменяется проницаемость постсинаптической мембраны для различных ионов (Na+, K+, Cl-). В зависимости от иона и направления его движения постсинаптическая мембрана деполяризуется или гиперполяризуется и на ней возникает возбуждающий или тормозной потенциал.Отметим, что электроуправляемых каналов на постсинаптической мембране нет, поэтому сама к генерации ПД она не способна. Возбуждающий постсинаптический потенциал (ВПСП), возникающий в результате поступления в клетку ионов Na+, электротонически распространяется за пределы постсинаптической мембраны и если его величина достаточно велика, деполяризует уже плазматическую мембрану до критического уровня. В мышечных клетках плазматическая мембрана сразу за пределами синапса имеет потенциал-чувствительные каналы, на ней относительно легко возникает полноценный потенциал действия. В нервных клетках возникновение возбуждения возможно только в аксоном холмике, поэтому активности одного из синапсов, которые, как правило, расположены далеко от него недостаточно для возникновения ПД. Нервная клетка воспринимает (суммирует) несколько синаптических сигналов и генерирует потенциал действия избирательно. В этом заключается интегративная функция нейрона. Тормозной постсинаптический (ТПСП) потенциал возникает при усилении проницаемости для калия и хлора, он гиперполяризует постсинаптическую мембрану, препятствуя электротоническому распространению возбуждения по мембране нервной клетки. Процесс торможения имеет место только на межнейронных синапсах. Принципы работы возбуждающего и тормозного синапсов сходны. Разница заключается в том с какими специфическими каналами связаны рецепторы расположенные на постсинаптической мембране, от чего и зависит физиологическая реакция. 4. Инактивация медиатора И нактивация медиатора - последняя стадия синаптической передачи, обеспечивающая ее прекращение. Она необходима для реполяризации постсинаптической мембраны и восстановления исходного уровня ее мембранного потенциала, что обеспечивает возможность восприятия следующего сигнала. Наиболее распространенным является гидролитическое расщепление медиатора с помощью соответствующих ферментов, постоянно находящихся в синаптической щели. Для ацетилхолина, например, таким ферментом является холинэстераза, для норадреналина - моноаминооксидаза. Эти ферменты начинают разрушать медиатор на исходные компоненты, сразу, как только он попадает в синаптическую щель, и скорость разрушения настолько велика, что при небольших количествах медиатора, он разрушается еще до того, как достигает постсинаптической мембраны. Продукты расщепления либо поступают в кровь и циркулируют как его предшественники, либо за счет пиноцитоза осуществляется их «обратное всасывание» в синаптическое окончание, где происходит их восстановление и упаковка в синаптические пузырьки и медиатор может использоваться повторно. Свойства синапсов:
|