физиология 1 зачет. физа 1 зачет. Вопросы коллоквиума по разделу Физиология возбудимых тканей
 Скачать 0.86 Mb. Скачать 0.86 Mb.
|

|
6. Потенциал действия, его фазы, ионные механизмы. Изменения проницаемости клеточной мембраны при возбуждении. П 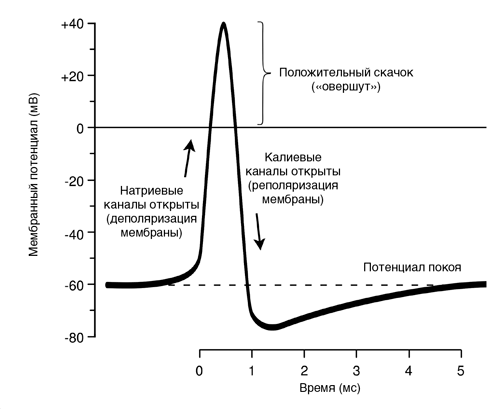 Д – процесс возбуждения, это быстрое колебание величины МП вследствие активации и инактивации ионных каналов и диффузии ионов в клетку и из клетки. ПД подчиняется закону «всё или ничего», но не подчиняется закону силы. 3 фазы. Д – процесс возбуждения, это быстрое колебание величины МП вследствие активации и инактивации ионных каналов и диффузии ионов в клетку и из клетки. ПД подчиняется закону «всё или ничего», но не подчиняется закону силы. 3 фазы. 1) Фаза деполяризации – уменьшение заряда клетки до нуля. Ворота Na-каналов начинают открываться при достижении деполяризации клетки, равной 60% от порогового потенциала, который обычно составляет -40 мВ, в результате Na+ начинает поступать в клетку. Когда деполяризация клетки достигает критического уровня (величины МП), открывается дополнительно большое число электроуправляемых m-ворот Nа-каналов, и Na+ лавиной устремляется в клетку согласно конц. и эл. градиентам (в первую фазу ПД клетка имеет «-» заряд). В итоге МП =0. 2) Фаза инверсии – изменение знака заряда клетки на противоположный. Имеет 2 части. Восходящая часть обеспечивается в основном входом Na+ в клетку, поэтому число «+» ионов в клетке превосходит число «-» ионов, заряд внутри клетки становится «+», снаружи – «-». Теперь эл градиент препятствует входу Na+ внутрь клетки, проводимость Na+ снижается. Нисходящая часть начинается через 1 мс и более после начала деполяризации. Рост ПД прекращается вследствие закрытия натриевых h-ворот и открытия ворот К-каналов. Поскольку К+ находится внутри клетки, он быстро выходит из клетки согласно конц. градиенту, в клетке уменьш. число «+» ионов и заряд клетки снова начинает уменьшаться. Так продолжается до полного исчезновения «+» заряда внутри клетки. 3) Фаза реполяризации – восстановление ПП. Она связана с тем, что проницаемость клеточной мембраны для К+ всё ещё высока, К+ продолжает быстро выходить из клетки по конц. градиенту. Поскольку клетка теперь снова имеет внутри «-», а снаружи «+», эл. градиент препятствует выходу К+ из клетки. Его проницаемость снижается, хотя К+ продолжает выходить из клетки. 7. Изменение возбудимости во время генерации потенциала действия. Характеристика рефрактерности и экзальтации. В 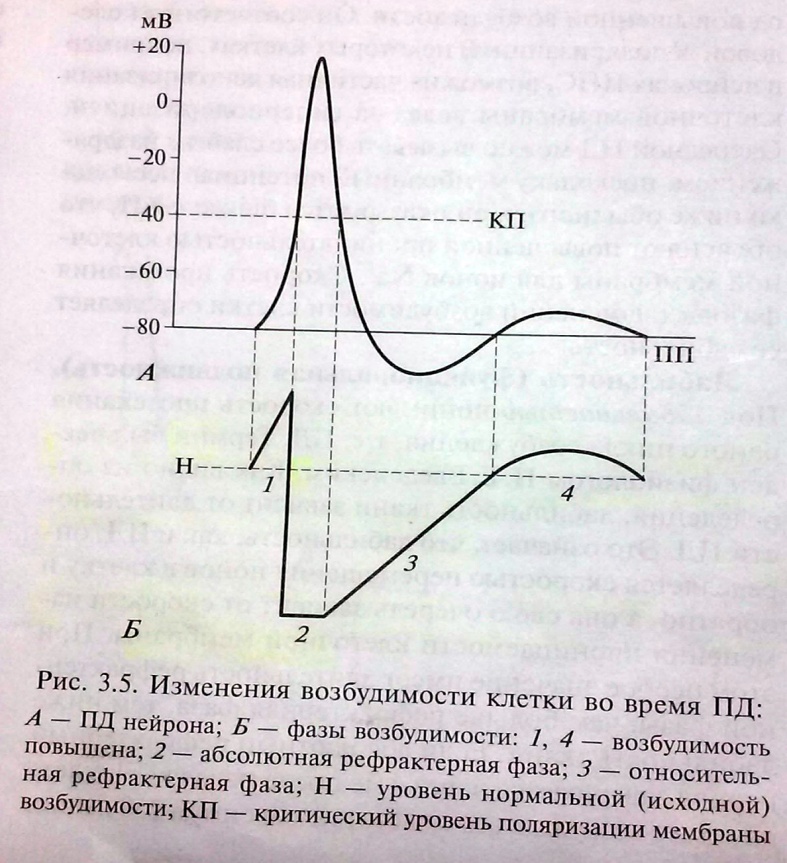 озбудимость клетки в разные фазы ПД. Во время возбуждения клетки ее возбудимость быстро и существенно изменяется. Различают несколько фаз изменения возбудимости, каждая из которых строго соответствует определенной фазе ПД и так же, как и фазы ПЛ. определяется состоянием проницаемости клеточной мембраны для ионов. озбудимость клетки в разные фазы ПД. Во время возбуждения клетки ее возбудимость быстро и существенно изменяется. Различают несколько фаз изменения возбудимости, каждая из которых строго соответствует определенной фазе ПД и так же, как и фазы ПЛ. определяется состоянием проницаемости клеточной мембраны для ионов. 1. Кратковременное повышение возбудимости в начале развития ПД, когда уже возникла частичная деполяризация клеточной мембраны. Если деполяри-зация не достигает КП, то регистрируется локальный потенциал. При достижении величины КП развивается ПД. Возбудимость клетки повышена еще и потому, что при деполяризации ее на 60 % ΔV начинают открываться потенциалуправляемые быстрые Na-кaналы. При этом достаточно небольшого увеличения силы раздражителя, чтобы деполяризация достигла КП, при которой возникает ПД. 2. Абсолютная рефрактерная фаза — это полная невозбудимость клетки (возбудимость равна нулю). Она соответствует пику ПД и продолжается у нервного волокна 1 — 2 мс; если ПД более продолжителен, то более продолжительна и абсолютная рефрактерная фаза. В этот период времени клетка не отвечает на раздражения любой силы. Невозбудимость клетки в фазы деполяризации и инверсии (в первую ее половину — восходящая часть пика ПД) объясняется тем, что потенциалуправляемые m-ворота Na-каналов уже открыты и Na+ быстро поступает в клетку по всем открытым каналам. Те ворота Na-каналов, которые еще не успели открыться, открываются под влиянием деполяризации — уменьшения мембранного потенциала, поэтому дополнительное раздражение клетки относительно движения Na+ в клетку ничего изменить не может. ПД либо совсем не возникает при раздражении, если оно мало, либо возникает максимальным, если действует раздражение достаточной силы (пороговой или сверхпороговой). В период нисходящей части фазы инверсии клетка невозбудима потому, что закрываются инактивационные h-ворота Na-каналов (соответствует вершине ПД), в результате чего клеточная мембрана непроницаема для Na+ даже при сильном раздражении. Кроме того, в этот период открываются уже в большом количестве К-каналы и К+ быстро выходит из клетки, обеспечивая нисходящую часть фазы инверсии и реполяризацию. Абсолютная рефрактерная фаза продолжается и в период реполяризации клетки до достижения КП ±10 мВ. 3. Относительная рефрактерная фаза — это период восстановления возбудимости клетки, когда сильное раздражение может вызвать новое возбуждение. Она соответствует конечной части фазы реполяризации (начиная от КП ±10мВ) и следовой гиперполяризации клеточной мембраны, если таковая имеется. Пониженная возбудимость явл. следствием все еще повышенной проницаемости для К+ и избыточным выходом К+ из клетки. Поэтому, чтобы вызвать возбуждение в этот период, необходимо приложить более сильное раздражение, так как выход К+ из клетки препятствует ее деполяризации. Кроме того, в период следовой гиперполяризации мембранный потенциал больше и, естественно, дальше отстоит от КП. Если реполяризация в конце пика ПД замедляется, то относительная рефрактерная фаза включает и период замедления реполяризации, и период гиперполяризации, т. е. продолжается до возвращения мембранного потенциала к исходному уровню после гиперполяризации. Продолжительность относительной рефрактерной фазы вариабельна, у нервных волокон она невелика и составляет несколько миллисекунд. 4. Фаза экзальтации— период повышенной возбудимости. Он соответствует следовой деполяризации. В некоторых клетках, например в нейронах ЦНС, возможна частичная деполяризация клеточной мембраны вслед за гиперполяризацией. Очередной ПД можно вызывать более слабым раздражением, поскольку мембранный потенциал несколько ниже обычного, и он оказывается ближе к КП, что объясняют повышенной проницательностью клеточной мембраны для ионов Na+. Скорость протекания фазовых изменений возбудимости клетки определяет ее лабильность. 8. Формы возбуждения: локальное (местное) и распространяющееся (импульсивное).
Повышение возбудимости клетки во время локального потенциала объясняется тем, что клеточная мембрана оказывается частично деполяризованной. Если КП во время локального потенциала остаётся на постоянном уровне, для достижения ВП во время локального потенциала нужен значительно меньшей силы раздражитель. Амплитуда ПД не зависит от силы раздражения, т.к. он возникает вследствие регенеративного процесса. Возникновение ПД обеспечивается величиной ПП, достигшей критического уровня.
Вывод: в организме животного и человека наблюдается местное и импульсное возбуждение. Возникновение того или иного вида возбуждения зависит от степени развития ткани и силы раздражителя. 9. Учение Н.Е. Введенского о физиологической лабильности. Под лабильностью понимают скорость протекания одного цикла возбуждения, т.е. ПД. Термин был введён физиологом Н.Е. Введенским. Как видно из определения, лабильность ткани зависит от длительности ПД. Это означает, что лабильность, как и ПД, определяется скоростью перемещения ионов в клетку и обратно, а она в сою очередь зависит от скорости изменения проницаемости клеточной мембраны. При этом особое значение имеет длительность рефрактерной фазы: чем ниже рефрактерная фаза (невозбудимость), тем ниже лабильность ткани. Если абсолютный рефрактерный период завершается через 4 мс после начала ПД, клетка может возбуждаться с частотой максимум 250 имс/с. Мерой лабильности явл. максимальное число ПД, которое ткань может воспроизвести в 1 с. В эксперименте лабильность исследуют в процессе регистрации максимального числа ПД, которое может воспроизвести клетка при увеличении частоты ритмического раздражения. Лабильность различных тканей существенно различается. Так, лабильность нервного волокна равна 200-400 имп/с, мышечного -100-200, нервно-мышечного синапса – 30-50. Лабильность ткани понижается при длительном бездействии органа и при утомлении, а также в случае нарушения иннервации. При постепенном увеличении ритмического раздражения лабильность ткани повышается, т.е. ткань отвечает более высокой частотой возбуждения по сравнению с исходной частотой. Это явление, называемое усвоением ритма раздражения, было открыто физиологом А.А. Ухтомским. 10. Законы раздражения (Пфлюгера). Генерация ПП подчиняется 3м законам раздражения. 1) Закон «всё или ничего». При достижении пороговой силы («всё») раздражающего стимула дальнейшее увеличение его интенсивности или продолжит. раздражения не изменяет характеристик ПД. Закон не являются абсолютным, а носит относительный характер: При действии раздражителей подпороговой силы видимая реакция отсутствует, но возникает местная реакция (локальный ответ) При действии пороговых раздражителей растянутая мышечная ткань дает большую амплитуду сокращения, чем не растянутая. При регистрации суммарной активности целостного образования (скелетная мышца, состоящая из отдельных мышечных волокон, нервный ствол, состоящий из множества нервных волокон) проявляется другая зависимость: Чем больше сила раздражителя, тем больше величина ответной реакции. 2  ) Закон силы-времени отражает зависимость пороговой силы раздражителя от времени его действия для возникновения возбуждения и гласит: возникновение распространяющегося возбуждения зависит не только от силы раздражителя, но и от времени, в течение которого он действует. Чем больше по силе раздражитель, тем меньшее время он должен действовать для возникновения возбуждения ) Закон силы-времени отражает зависимость пороговой силы раздражителя от времени его действия для возникновения возбуждения и гласит: возникновение распространяющегося возбуждения зависит не только от силы раздражителя, но и от времени, в течение которого он действует. Чем больше по силе раздражитель, тем меньшее время он должен действовать для возникновения возбужденияЗависимость носит обратный характер и имеет вид гиперболы. Из этого следует, что на кривой "силы-времени" имеются области, которые не подчиняются этому закону. Если сила раздражителя будет меньше некоторой (пороговой) величины, то возбуждение не возникнет даже при длительном его воздействии. Наоборот, если время воздействия будет очень коротким, то возбуждение тоже не возникнет даже при воздействии очень большого по силе раздражителя (в физиотерапии токи высокой частоты используются для получения калорического эффекта) При измерении хронаксии мышцы фактически получается зн-е хронаксии, иннервирующей её нервного волокна. Если нерв повреждён, тогда раздражающий стимул выявляет хронаксию мышечных волокон. 3) Закон аккомодации. Аккомодация – приспособление ткани к медленно нарастающему по силе раздражителю. Раздражающее действие тока зависит не только от амплитуда эл. сигнала, но и от скорости (крутизны) его нарастания во времени чем быстрее это нарастание, тем сильнее выражено раздражающее действие тока. При действии медленно нарастающей силы раздражителя генерации ПД не происходит, т.к. возбудимая клетка «приспособилась» к действию этого раздражителя, что и получило название аккомодации. 11. Структурно-функциональная классификация нервных волокон (Дж.Эрлангера - Х.Гассера). Законы проведения возбуждения в нервных волокнах. Нервные волокна – это отростки нейронов, с помощью которых осущ. связь нейронов между собой, а также с иннервируемыми клетками и рецепторами. Пучки нервных волокон образуют периферические нервы. 1) Безмиелиновые. Их оболочку образуют леммоциты (шванновские клетки), которые погружаются в осевые цилиндры нейронов. 2) Миелиновые. Их оболочку образуют в ПНС леммоциты , а в ЦНС – олигодендроциты. В миелиновых волокнах мезаксон удлиняется и спирально заркчивается вокруг осевого цилиндра, образуя слой миелина толщиной до 10 мкм. Миелиновая оболочка имеет перехваты Ранвье. Протяжённость перехватов в ПНС 0,25-1,0 мкм, в волокнах ЦНС – до 14 мкм. Участки волокон между узловыми перехватами наз. межузловыми сегментами. Классификация: Волокна А и В – миелинизированные, С – безмиелиновые.
|