анатомия Варинт -8. Задача одновременного развития силы и объема мышц плеча решается с помощью базовых
 Скачать 365 Kb. Скачать 365 Kb.
|

Эпителиальные ткани. Строение, функции и классификация эпителиев. (сделать рисунки).Эпителии покрывают поверхность тела, серозные полости тела, внутреннюю и наружную поверхности многих внутренних органов, образуют секреторные отделы и выводные протоки экзокринных желез. Эпителий представляет собой пласт клеток, под которым есть базальная мембрана. Эпителии подразделяются на покровные, которые выстилают тело и все полости, имеющиеся в организме, и железистые, которые вырабатывают и выделяют секрет. Функции: 1. разграничительная (барьерная) (контакт с внешней средой); 2. защитная (внутренней среды организма от повреждающего действия механических, физических, химических факторов среды; выработка слизи, обладающей антимикробным действием); 3. обмен веществ между организмом и окружающей средой; 4. секреторная; 5. экскреторная; 6. развитие половых клеток и др.; 7. рецепторная (сенсорная). Развитие: из всех 3-х зародышевых листков: 1. Кожная эктодерма; 2. Кишечная энтодерма: - прехордальная пластинка; 3. Мезодерма: - нервная пластинка. Общие признаки строения эпителия: 1. Клетки лежат плотно друг к другу, образуя сплошной пласт. 2. Гетерополярность - апикальные (верхушка) и базальные части клеток отличаются по строению и по функции; а в многослойном эпителии - отличие в строении и функции слоев. 3. Состоит только из клеток, межклеточное вещество практически отсутствует (десмосомы). 4. Эпителий всегда располагается на базальной мембране (углеводнобелковолипидный комплекс с тончайшими фибриллами) и им отграничивается от подлежащей рыхлой соединительной ткани. 5. Эпителий участвует в выделении секрета. 6. Характерна повышенная регенераторная способность, обусловленная пограничностью. 7. не имеет собственных кровеносных сосудов, питается диффузно через базальную мембрану, за счет сосудов подлежащей рыхлой соединит. ткани. 8. Хорошо иннервирован (много нервных окончаний). КЛАССИФИКАЦИЯ ЭПИТЕЛИАЛЬНОЙ ТКАНИ Морфофункциональная классификация (А. А. Заварзина):  Схема строения различных видов эпителия: (1 — эпителий, 2 — базальная мембрана; 3 — подлежащая соединительная ткань) А — однослойный однорядный цилиндрический, Б — однослойный однорядный кубический, В — однослойный однорядный плоский; Г — однослойный многорядный; Д — многослойный плоский неороговевающий, Е — многослойный плоский ороговевающий; Ж1 — переходный при растянутой стенке органа, Ж2 — переходный при спавшейся. Ультраструктура мышечного волокна. Саркомер. Теория мышечного сокращения, (сделать рисунки).Уровень возбудимости клетки зависит от фазы потенциала действия. В фазу локального ответа возбудимость возрастает. Эту фазу возбудимости называют латентным дополнением. В фазу реполяризации потенциала действия, когда открываются все натриевые каналы и ионы натрия лавинообразно устремляются в клетку, никакой, даже сверхсильный, раздражитель не может стимулировать этот процесс. Поэтому фазе деполяризации соответствует фаза полной невозбудимости или абсолютной рефрактерности, т.е. в фазу реполяризации все большая часть натриевых каналов закрывается. Однако они могут вновь открываться при действии сверхпорогового раздражителя, т.е. возбудимость начинает вновь повышаться. Этому соответствует фаза относительной невозбудимости или относительной рефрактерности. Во время следовой деполяризации мембранный потенциал находится у критического уровня, поэтому даже допороговые стимулы могут вызвать возбуждение клетки. Следовательно, в этот момент ее возбудимость повышена. Эта фаза называется фазой экзальтации или супернормальной возбудимостью. В момент следовой гиперполяризации мембранный потенциал выше исходного уровня, т.е. дальше критического уровня деполяризации и ее возбудимость снижена. Она находится в фазе субнормальной возбудимости. Следует отметить, что явление аккомодации также связано с изменением проводимости ионных каналов. Если деполяризующий ток нарастает медленно, то это приводит к частичной инактивации натриевых и активации калиевых каналов. Поэтому развитие потенциала действия не происходит. В организме имеются 3 типа мышц: скелетные или поперечно-полосатые, гладкие и сердечные. Скелетные мышцы обеспечивают перемещение тела в пространстве, поддержание позы тела за счет тонуса мышц конечностей и тела. Гладкие мышцы необходимы для перистальтики органов желудочно-кишечного тракта, мочевыводящей системы, регуляции тонуса сосудов, бронхов и т.д. Сердечная мышца служит для сокращения сердца и перекачивания крови. Все мышцы обладают возбудимостью, проводимостью и сократимостью, а сердечная и многие гладкие мышцы – автоматией – способностью к самопроизвольному сокращению. Основным морфо-функциональным элементом нервно-мышечного аппарата скелетных мышц является двигательная единица. Она включает мотонейрон спинного мозга с иннервируемыми его аксоном мышечными волокнами. Внутри мышцы этот аксон образует несколько концевых веточек. Каждая такая веточка образует контакт – нервно-мышечный синапс на отдельном мышечном волокне. Нервные импульсы, идущие от мотонейрона, вызывают сокращения определенной группы мышечных волокон. Скелетные мышцы состоят из мышечных пучков, образованных большим количеством мышечных волокон. Каждое волокно – это клетка цилиндрической формы диаметром 10-100 мкм и длиной от 5 до 400 мкм. Оно имеет клеточную мембрану – сарколемму. В саркоплазме находится несколько ядер, митохондрии, образования саркоплазматического ретикулума и сократительные элементы – миофибриллы. Саркоплазматический ретикулум имеет своеобразное строение. Он состоит из системы поперечных, продольных трубочек и цистерн. Поперечные трубочки – это впячивания сарколеммы внутрь клетки. К ним примыкают продольные трубочки с цистернами. Благодаря этому, потенциал действия может распространяться от сарколеммы на систему саркоплазматического ретикулума. В мышечном волокне содержится более 1000 миофибрилл, расположенных вдоль него. Каждая миофибрилла состоит из 2500 протофибрилл или миофиламентов. Это нити сократительных белков актина и миозина. Миозиновые протофибриллы толстые, актиновые тонкие. На миозиновых нитях расположены отходящие под углом поперечные отростки с головками. У скелетного мышечного волокна при световой микроскопии видна поперечная исчерченность, т.е. чередование светлых и темных полос. Темные полосы называются А дисками или анизотропными, светлые I-дисками или изотропными. В А дисках сосредоточены нити миозина, обладающие анизотропией и поэтому имеющие темный цвет, I-диски образованны нитями актина. В центре I-дисков видна тонкая Z-пластинка. К ней прикрепляются актиновые протофибриллы. Участок миофибриллы между двумя Z-пластинками называется саркомером. Саркомер – структурный элемент миофибрилл. В покое толстые миозиновые нити лишь на небольшом расстояние входят в промежуток между актиновыми. Поэтому в средней части А диска имеется более светлая Н-зона, где нет актиновых нитей. При электронной микроскопии в ее центре видна очень тонкая М-линия. Она образована цепями опорных белков, к которым крепятся миозиновые протофибриллы. Саркомер — базовая сократительная единица поперечнополосатых мышц, представляющая собой комплекс нескольких белков, состоящий из трёх разных систем волокон. Из саркомеров состоят миофибриллы. Саркомер - это участок миофибриллы между двумя соседними Z-линиями, структурно-функциональная единица поперечнополосатой мышечной ткани. Сокращение — это изменение механического состояния миофибриллярного аппарата мышечных волокон под влиянием нервных импульсов. В 1939 г В.А. Энгельгардтом и М.Н. Любимовой было установлено, что миозин обладает свойствами фермента аденозинтрифосфатазы, расщепляющей АТФ. Вскоре было установлено, что при взаимодействии актина с миозином образуется комплекс — актомиозин, ферментативная активность которого почти в 10 раз выше активности миозина (А. С. Уент - Дьорди, 1940). В этот период и начинается разработка современной теории мышечного сокращения, которая получила название теории скользящих нитей. Согласно этой теории «скольжения» в основе сокращения лежит взаимодействие между актиновыми и миозиновыми нитями миофибрилл вследствие образования поперечных мостиков между ними. Во время скольжения сами актиновые и миозиновые нити не укорачиваются, но длина саркомера изменяется. В расслабленной, а тем более растянутой мышце активные нити располагаются дальше от центра саркомера, и длина саркомера больше. При изотоническом сокращении мышцы актиновые нити скользят по направлению к центру саркомера вдоль миозиновых нитей. Нити актина прикреплены к Z-мембране, тянут ее за собой, и саркомер укорачивается. Суммарное укорочение всех саркомеров вызывает укорочение миофибрилл, и мышца сокращается. В настоящее время принята следующая модель скольжения нитей актина. Импульс возбуждения по двигательному нейрону достигает нервно-мышечного синапса — концевой пластинки, где освобождается ацетилхолин, который взаимодействует с постсинаптической мембраной, и в мышечном волокне возникает потенциал действия, т.е. наступает возбуждение мышечного волокна. При связывании ионов Са++ с тропонином (сферические молекулы которого «сидят» на цепях актина) последний деформируется, толкая тропомиозин в желобки между двумя цепями актина. При этом становится возможным взаимодействие актина с головками миозина и возникает сила сокращения. Головки миозина совершают «гребковые» движения и продвигают актиновую нить по направлению к центру саркомера. Головок у миозиновых нитей множество, они тянут актиновую нить с объединенной, суммарной силой. При одинаковом гребковом движении головок саркомер укорачивается примерно на 1 % его длины (а при изотоническом сокращении саркомер мышцы может укорачиваться на 50 % длины за десятые доли секунды), следовательно, поперечные мостики должны совершать примерно 50 «гребковых» движений за тот же промежуток времени. Совокупное укорочение последовательно расположенных саркомеров миофибрилл приводит к заметному сокращению мышцы. Одновременно происходит гидролиз АТФ. После окончания пика потенциала действия активируется кальциевый насос (Са — зависимая АТФ-аза) мембраны саркоплазматического ретикулума. За счет энергии, выделяющейся при расщеплении АТФ, кальциевый насос перекачивает ионы Са++ обратно в цистерны саркоплазматического ретикулума, где Са++ связывается белком кальсеквестрином. Концентрация ионов Са++ в цитоплазме мышц снижается до 10-8 м, а в саркоплазматическом ретикулуме повышается до 10-3 м. Снижение уровня Са++ в саркоплазме подавляет АТФ-азную активность актомиозина; при этом поперечные мостики миозина отсоединяются от актина. Происходит расслабление, удлинение мышц в результате пассивного движения (без затрат энергии). Таким образом, сокращение и расслабление мышцы представляет собой серию процессов, развертывающихся в следующей последовательности: нервный импульс --> выделение ацетилхолина пресинаптической мембраной нервно-мышечного синапса --> взаимодействие ацетилхолина с постсинаптической мембраной синапса --> возникновение потенциала действия --> электромеханическое сопряжение (проведение возбуждения по Т-канальцам, высвобождение Са++ и воздействие его на систему тропонин-тропомиозин-актин) --> образование поперечных мостиков и «скольжение» актиновых нитей вдоль миозиновых --> снижение концентрации ионов Са++ вследствие работы кальциевого насоса --> пространственное изменение белков сократительной системы --> расслабление миофибрилл. После смерти мышцы остаются напряженными, наступает так называемое трупное окоченение, так как поперечные связи между филаментами актина и миозина не могут разорваться из-за отсутствия энергии АТФ и невозможности работы кальциевого насоса. При сокращении мышцы длина А-дисков не меняется, J-диски укорачиваются, а Н-зона А-дисков может исчезать. 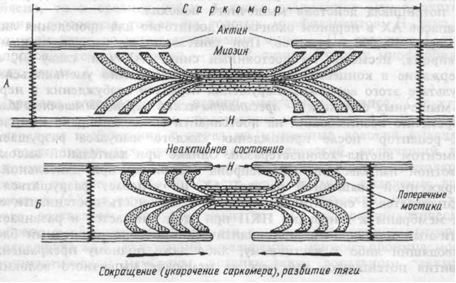 Рис.1. Сокращение мышцы. А — Поперечные мостики между актином и миозином разомкнуты. Мышца находится в расслабленном состоянии. Б — Замыкание поперечных мостиков между актином и миозином. Совершение головками мостиков гребковых движений по направлению к центру саркомера. Скольжение актиновых нитей вдоль миозиновых, укорочение саркомера, развитие тяги. |